 2022, Vol. 41
2022, Vol. 41扩展功能
文章信息
- 王新建, 刘新, 戴昭鑫
- WANG Xinjian, LIU Xin, DAI Zhaoxin
- 黄河三角洲东方白鹳的巢址选择预测分析
- Predictive Analysis of Nest Site Selection of Oriental Stork in the Yellow River Delta
- 四川动物, 2022, 41(3): 264-271
- Sichuan Journal of Zoology, 2022, 41(3): 264-271
- 10.11984/j.issn.1000-7083.20210408
-
文章历史
- 收稿日期: 2021-11-22
- 接受日期: 2022-03-30






2. 中国测绘科学研究院,北京 100102
2. Chinese Academy of Surveying and Mapping, Beijing 100102, China
东方白鹳Ciconia boyciana是一种大型迁徙水鸟,CITES附录Ⅰ物种,被世界自然保护联盟(IUCN)濒危物种红色物种名录列为EN,国家一级重点保护野生动物。受人类活动影响,东方白鹳在过去几十年中数量急剧减少,目前全球野生数量仅3 000只左右,主要分布在俄罗斯、日本、韩国以及中国的东北、长江流域及黄河三角洲(Wei et al., 2016)、内蒙古(张国钢等,2016)等区域。其中,黄河三角洲自然保护区作为东北亚鸟类迁徙路线咽喉要道,是东方白鹳重要的迁徙停歇地和越冬地,并自2003年后逐渐演变为东方白鹳北方繁殖区外的最大繁殖地(Shimazaki,2004)。近年来,区域内种群数量不断增加,保护区管理站提供的营巢位点逐渐饱和。巢穴作为东方白鹳栖息、繁殖的重要场所,其质量直接影响繁殖成功率。尤其面对黄河三角洲区域东方白鹳种群数量不断增加、巢穴点逐渐饱和的问题,开展黄河三角洲东方白鹳巢穴选择及空间预测对于未来保护该物种生存和繁殖具有重要现实意义(Cox et al., 2019)。
东方白鹳的巢址选择已有部分研究,包括巢址的定性及描述性研究(吴建东等,2001),结合生境适宜性模型和GIS技术等对东方白鹳的巢址选择开展定量研究,如鄱阳湖区域(Wei et al., 2016)、日本区域(Yamada et al., 2019)。不同的区域及生态系统内东方白鹳巢址的选择模式不同(Cheng et al., 2020)。朱明畅等(2015)在黄河三角洲开展了东方白鹳巢址预测研究,但主要聚焦在黄河三角洲的3个管理站附近。随着黄河三角洲东方白鹳种群数量增多,其巢穴点不再局限于3个管理站附近,亟需对管理站范围外的其他区域进行选址研究。
因此,本文拟通过二项Logistic回归模型对黄河三角洲(生态旅游区除外)东方白鹳巢址选择的生境因子进行分析,揭示不同生境因子对东方白鹳巢址选择的影响,绘制研究区东方白鹳巢址选择的空间分布预测图,并对研究区东方白鹳巢址选择的适宜性进行多等级划分。在此基础上,针对性地进行湿地的恢复与保护,建立栖息地保护区,为濒危物种的保护和科学管理提供依据。
1 数据与方法 1.1 研究区概况黄河三角洲位于山东省东营市(118°24′~119°20′E,37°29′~38°12′N),北临渤海,东临莱州湾;选择位于黄河三角洲范围内的东营市陆地区域为研究区,面积4 264.33 km2(图 1)。属于暖温带大陆性季风气候,四季分明,年均气温12~13 ℃,年均降水量550~600 mm,年均无霜期200 d左右,平均海拔11.2 m(陈浩等,2019)。地势平坦,拥有丰富的湿地、植被、水生动物等资源,为东方白鹳提供了充足的食物资源;其中水田214.51 km2,坑塘水面473.20 km2,内陆滩涂282.27 km2;植被主要有芦苇Phragmites australis、盐地碱蓬Suaeda salsa、柽柳Tamarix chinensis、白茅Imperata cylindrica等;水生动物主要有中国毛虾Acetes chinensis、三疣梭子蟹Portunus trituberculatus、黄姑鱼Nibea albiflora、泥鳅Misgurnus anguillicaudatus、黄鳝Monopterus albus等。

|
| 图 1 黄河三角洲东方白鹳巢址分布和研究区 Fig. 1 Study area and nest sites of Ciconia boyciana in the Yellow River Delta |
| |
为了确定东方白鹳巢址选择的影响因素,研究中使用黄河三角洲高精度的土地利用数据、高压塔分布数据以及2019年3—6月统计的研究区内57个东方白鹳巢址分布数据。为了获取对照样方,将研究区划分为20×20的格网,剔除含有东方白鹳巢址的格网后,随机选择57个点为对照样方。
通过中国测绘科学研究院WJ-Ⅲ工作站对土地利用数据进行图斑融合,融合前地类图斑相对比较零散,个别小面积的地类图斑会对数据采集过程带来一定程度的影响,融合后的土地利用图斑整体具有一定的连续性。
结合土地利用类型和东方白鹳的生活习性,参考相关研究(Gaillard et al., 2010;段玉宝等,2015),初步确定10个生境(水田、旱地、公路、港口、机场、高压塔、广场和商业服务用地、坑塘水面、河流、内陆滩涂),统计巢址样方和对照样方到各生境的距离: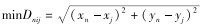
利用单个样本的Kolmogorov-Smirnov Test检验数据是否符合正态分布。若数据符合正态分布,使用独立样本的t检验,反之使用Spearman秩相关性检验各因子间的相关性。2个因子相关系数的绝对值越大,说明其相关性越强;为了保证因子之间的相互独立性,需要将相关系数达到一定值的因子进行选择性剔除,选择将2个因子之间相关性系数的绝对值>0.5的因子进行选择性剔除;将相互独立的变量采用“向前”条件(以假定参数为基础作似然比概率检验,向前逐步选择自变量)二项Logistic回归,得到资源选择函数,其模型为:W(x)=exp(β0+β1x1+……+βkxk),式中,x为相互独立的生境因子,β为生境因子的系数,k为独立生境因子个数。资源选择函数模型能够很好地体现东方白鹳巢址选择时对各因子偏好的正负相关性以及偏好程度。
根据资源选择函数可以得到物种对资源选择的概率: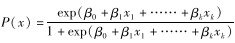
基于现存的每个巢址样方独立于每个单独的格网之中为原则,统计东方白鹳巢址之间的最近距离来确定格网分辨率。为了验证空间分辨率选择的有效性,适当对其他分辨率的格网进行空间预测,将其结果进行对比分析。通过资源选择函数和资源选择概率,对研究区进行空间格网化概率预测,求取每个格网中心点的概率值,通过自然断点法将获取的格网中心点的资源选择概率值分成12类,依次以间断点将数据分成2个概率区间,分别统计在该间断点处的验证概率P(i)=[nc+(nl-nu)]/N,式中,nc为第i个间断点处对照样方的个数,nl为巢址样方的个数,nu为第i个间断点处巢址样方的个数,N为对照样方和巢址样方的总数。
通过验证概率将研究区进行多等级划分。通过反距离权重法进行插值,得到东方白鹳巢址选择的适宜区空间分布。
数据处理在SPSS 20.0和ArcGIS 10.7中完成。
2 结果分析 2.1 生境因子的相关性对初选的10个生境因子进行Kolmogorov-Smirnov Test检验,发现各生境因子并不完全符合正态分布,因此选择Spearman相关检验分析因子之间的相关性;两者之间相关性系数的绝对值>0.5的因子有距港口距离与距水田距离、距内陆滩涂距离(图 2);为了保证因子之间的相互独立性,剔除距港口距离的生境因子。

|
| 图 2 生境因子间的相关性分析 Fig. 2 Correlation analysis among habitat factors Dpl. 距水田距离, Drs. 距河流水面距离, Ds. 距广场和商业服务用地距离, Dt. 距内陆滩涂距离, Dpw. 距坑塘水面距离, Ddl.距旱地距离, Da. 距机场距离, Dr. 距公路距离, Dp. 距港口距离, Dht. 距高压塔距离 Dpl. distance to paddy land, Drs. distance to river surface, Ds. distance to square and commercial services, Dt. distance to tideland, Dpw. distance to pit and water, Ddl. distance to dry land, Da. distance to airport, Dr. distance to road, Dp. distance to port, Dht. distance to high-voltage tower |
| |
通过Spearman相关性分析筛选的生境因子并进行标准化,采用“向前”条件进行二项Logistic回归(表 1),具有统计学意义的变量包括距水田距离、距广场和商业服务用地距离、距内陆滩涂距离、距坑塘水面距离和距高压塔距离;东方白鹳巢址选择的资源选择函数为:logist(P)=-1.715Dpl+0.810Ds-1.117Dpw-2.511Dht-constant;东方白鹳巢址选择的概率为:P=elogist(P)/[1+elogist(P)];模型的正确率为87.7%,拟合优度为0.663。东方白鹳巢址选择与距水田、内陆滩涂、坑塘水面、高压塔距离呈负相关,其中,距高压塔距离对东方白鹳巢址选择的影响最大;与距广场和商业服务用地距离呈正相关。结果表明:东方白鹳巢址的选择偏好距水田、内陆滩涂、坑塘水面、高压塔较近,而距广场和商业服务用地较远的地方。
| 生境因子 | 因子系数 | P | Odds ratio | 95%置信区间 | |
| 下限 | 上限 | ||||
| 距水田距离 | -1.715 | 0.003 | 0.183 | 0.061 | 0.551 |
| 距广场和商业服务用地距离 | 0.810 | 0.005 | 2.248 | 1.271 | 3.976 |
| 距内陆滩涂距离 | -0.889 | 0.012 | 0.411 | 0.206 | 0.821 |
| 距坑塘水面距离 | -1.170 | 0.001 | 0.311 | 0.152 | 0.633 |
| 距高压塔距离 | -2.511 | 0.011 | 0.081 | 0.012 | 0.567 |
| 常量 | -0.845 | 0.039 | 0.467 | ||
黄河三角洲研究区采集的57个巢址样方中,36个位于高压塔上,21个位于地面上。分别统计2类巢址到函数模型中的5个因子的平均距离发现:巢址与水田、高压塔、坑塘水面、内陆滩涂的距离相对较近,与广场和商业服务用地的距离相对较远,与资源选择函数的结果一致(表 2)。
| 筑巢类型 | 高压塔筑巢 | 地面上筑巢 | 总利用样方 |
| 距水田距离/m | 2 956.459 | 112.025 | 1 908.510 |
| 距广场和商业服务用地距离/m | 5 702.763 | 10 946.942 | 7 634.829 |
| 距内陆滩涂距离/m | 6 137.699 | 4 658.112 | 5 592.588 |
| 距坑塘水面距离/m | 2 569.890 | 2 877.366 | 2 683.173 |
| 距高压塔距离/m | 0 | 203.07 | 74.815 |
研究区东方白鹳巢址之间的最近距离为123.80 m,由此确定格网分辨率为80 m,以80 m× 80 m分辨率的格网对研究区进行规则格网化,分别统计每个格网中心点到模型中生境因子的距离,根据资源选择函数和资源选择概率求得每个格网中心点的生境选择概率值,根据自然断点法将研究区生境选择概率分为12类(0.04、0.13、0.22、0.31、0.41、0.50、0.60、0.70、0.78、0.87、0.93、1.00),分别统计每个间断点的验证概率,以0.60[P(0.60)=66.67%]、0.87[P(0.87)=72.81%]、0.93[P(0.93)=76.32%]为间断点将研究区分为4个等级(表 3)。
| 类别区间 | 对照样方/个 | 巢址样方/个 | 验证概率/% | 分类区间 | 划分类别 |
| 0.04 | 10 | 0 | 58.77 | ||
| 0.13 | 14 | 0 | 62.28 | ||
| 0.22 | 14 | 0 | 62.28 | ||
| 0.31 | 17 | 0 | 64.91 | 0~0.60 | 不太适宜区 |
| 0.41 | 19 | 0 | 66.67 | ||
| 0.50 | 19 | 0 | 66.67 | ||
| 0.60 | 20 | 1 | 66.67 | ||
| 0.70 | 27 | 2 | 71.93 | ||
| 0.78 | 28 | 4 | 71.05 | 0.60~0.87 | 一般适宜区 |
| 0.87 | 32 | 6 | 72.81 | ||
| 0.93 | 44 | 14 | 76.32 | 0.87~0.93 | 比较适宜区 |
| 1.00 | 57 | 57 | 50.00 | 0.93~1.00 | 最佳适宜区 |
东方白鹳巢址主要分布在黄河三角洲的东、东北、北部区域,整体呈环绕趋势;根据空间预测分布图,东方白鹳巢址选择以最佳适宜区为核心区依次向外扩散,分别为比较适宜区、一般适宜区、不太适宜区,其中,比较适宜区的辐射距离约1 000 m,一般适宜区的辐射距离约1 500 m。最佳适宜区的面积783.02 km2,比较适宜区486.62 km2,一般适宜区750.74 km2,不太适宜区2 243.95 km2(图 3)。

|
| 图 3 黄河三角洲研究区东方白鹳的适宜生境 Fig. 3 Distribution of suitable habitats of Ciconia boyciana in the Yellow River Delta Research Area |
| |
巢址是东方白鹳生存和繁殖的重要场所,对东方白鹳种群的繁衍具有重要意义。本文利用资源选择函数等模型,分析了生境因子对东方白鹳巢址选择的影响,并进行了空间巢址选择适宜性预测与等级划分。
巢址预测模型:研究根据优选的生境因子,创建了东方白鹳巢址选择预测模型(模型准确率达到87.7%);分析发现,东方白鹳巢址的选择倾向于距水田、内陆滩涂、坑塘、高压塔较近,而距广场和商业服务用地较远的地方。
适宜性分级划分:基于预测模型,采用自然断点法和验证概率对东方白鹳巢址适宜性区间划分为最佳适宜区、比较适宜区、一般适宜区、不太适宜区。
空间分布:基于空间预测结果,现存的巢址中的75.44%分布在最佳适宜区,占整个研究区面积的18.36%;空间分布格局主要以最佳适宜区为核心,按适宜性的等级向四周扩散。
本文对东方白鹳巢址选择时的生境因子的选择研究结果与现有的研究结果具有一致性(段玉宝等,2011;Zhou et al., 2013),主要表现在:巢址选择均受水资源(水田、坑塘)、人为因素(广场和商业服务用地)的影响,东方白鹳倾向于在距水资源丰富、距人为因素影响较小的区域筑巢;根据东方白鹳巢址选择空间预测分布图,可以发现,东方白鹳在一千二管理站附近选择筑巢的概率较低,而在大汶流管理站附近的概率较高,与朱明畅等(2015)对于自然保护区东方白鹳巢址的选择预测具有一致性。此外,研究发现黄河三角洲生态系统中东方白鹳的筑巢方式和其他生态系统存在一定的差异性。
3.1 不同生态系统的筑巢方式目前对于东方白鹳巢址选择适宜性的研究中,不同的生态系统中影响东方白鹳巢址选择的生境因子也有所不同。如本研究发现在黄河三角洲区域,1排高压电线塔贯穿于芦苇沼泽中,东方白鹳集中在高压塔上营巢繁殖,但这与东方白鹳历史繁殖地——三江平原地区以及洪河自然保护区(王健,李晓民,2006)在高大乔木和人工招引的三脚架上筑巢明显不同(张希国,2011),也与日本福井县东方白鹳筑巢点倾向于选择稻田(Yamada et al., 2019)存在一定的差异性,这表明不同的生态系统导致东方白鹳筑巢点的选择存在一定的差异性。黄河三角洲多盐碱地,少高大乔木,不能为东方白鹳提供充足的筑巢点,而区域内高压塔多分布在沼泽中,便于东方白鹳近距离隐蔽觅食。东方白鹳目前分布在欧洲、韩国、日本及中国东部不同地区,未来在进行东方白鹳巢址选择时,应根据不同地区生态系统特征进行适宜性评价。
3.2 同一生态系统的筑巢方式处于同一生态系统的筑巢方式存在一定的差异:相对于在高压塔上的巢址,地面上的巢址距水田的距离更近;在满足东方白鹳筑巢条件的区域,距水田较近的区域,东方白鹳选择在地面上筑巢。2类巢址到内陆滩涂和坑塘水面距离的平均值相差较小,表明东方白鹳筑巢受2个生境因子的影响较小。由2类巢址到距广场和商业服务用地距离的平均值可以发现,距广场和商业服务用地较近的巢址主要在高压塔上,而较远的巢址在地面上;距广场和商业服务用地距离较近,人为影响因素较大,高压塔上能够避免人为因素对东方白鹳的生存繁殖带来的影响,综上,人为因素是影响东方白鹳巢址选择方式的因素之一。
3.3 空间分辨率的选择对于目前东方白鹳巢址选择空间预测的格网分辨率的研究中,Wei等(2016)以0.05°×0.05°的栅格对鄱阳湖东方白鹳种群的空间分布进行了预测;Yamada等(2019)以东方白鹳栖息地的半径为依据,采用1 km×1 km分辨率的格网进行空间分布预测。本文确定的格网分辨率为80 m;为了验证空间分辨率选择的有效性,分别以250 m、500 m分辨率对研究区进行空间分布预测,并统计80 m、250 m、500 m分辨率下不同等级适宜区的巢址样方占比、面积占比(表 4): 巢址样方在不同等级适宜区的分布个数是一致的,不同等级适宜区的面积占比存在一定的差异,但差异较小,因此,在对东方白鹳巢址选择空间适宜性预测时,可适当减小分辨率以提高工作效率。
| 分辨率 | 不太适宜区 | 一般适宜区 | 比较适宜区 | 最佳适宜区 | ||||
| 巢址样方占比/% | 面积占比/% | 巢址样方占比/% | 面积占比/% | 巢址样方占比/% | 面积占比/% | 巢址样方占比/% | 面积占比/% | |
| 80 m | 1.75 | 52.62 | 8.77 | 17.61 | 14.04 | 11.41 | 75.44 | 18.36 |
| 250 m | 1.75 | 53.45 | 8.77 | 17.30 | 14.04 | 11.22 | 75.44 | 18.03 |
| 500 m | 1.75 | 52.37 | 8.77 | 17.95 | 14.04 | 11.44 | 75.44 | 18.24 |
本研究中生境因子的选择是基于东营市第三次国土调查数据,以土地利用类型命名生境因子的名称;统计57个巢址到高压塔的平均距离为74.815 m,其中21个地面上的巢址到高压塔的平均距离为203.07 m,36个在高压塔上,由此可以发现巢址选择与高压塔存在一定的相关性,因此,加入了距高压塔距离的生境因子;由回归模型可知,与距高压塔距离因子的相关系数为-2.511,显著性较强(P=0.011),由此可知,高压塔对东方白鹳巢址选择存在一定程度的影响。
通过二项Logistic回归建模,研究了东方白鹳巢址选择对生境因子的偏好性,通过反距离权重法对研究区东方白鹳巢址选择的适宜性进行多等级划分,能够筛选出不同质量的巢址选择栖息地(Chalfoun & Schmidt,2012),对黄河三角洲东方白鹳的保护和管理具有重要的意义。但是对于一个复杂的生态系统,本研究没有考虑东方白鹳种群内及与其他物种种群间的竞争关系;研究受生境因子的选择、样本数量的限制,预测结果可能存在一定的差异。本文对研究区进行多等级适宜性区域划分,缺少种群密度和环境容纳量分析,接下来可根据种群数量,针对不同等级适宜性区域进行湿地恢复与保护。
致谢: 山东省东营市自然资源局提供的相关巢址数据,数据处理和论文写作得到中国测绘科学研究院李成名研究员、吴政博士,山东科技大学刘新副教授的支持和帮助,在此表示感谢。
陈浩, 王仁卿, 刘建. 2019. 黄河三角洲城市土地利用与生态环境耦合关系[J]. 山东大学学报(理学版), 54(7): 11-20. |
段玉宝, 田秀华, 马建章, 等. 2015. 黄河三角洲东方白鹳繁殖期觅食栖息地的利用[J]. 生态学报, 35(8): 2628-2634. |
段玉宝, 田秀华, 朱书玉, 等. 2011. 黄河三角洲自然保护区东方白鹳的巢址利用[J]. 生态学报, 31(3): 666-672. |
王健, 李晓民. 2006. 黑龙江省洪河自然保护区东方白鹳巢址选择[J]. 东北林业大学学报, 34(1): 65-66. |
吴建东, 纪伟涛, 易武生. 2001. 在鄱阳湖越冬的东方白鹳的一些资料[J]. 四川动物, 20(1): 31-33. |
张国钢, 李淑红, 周景英, 等. 2016. 内蒙古图牧吉国家级自然保护区水鸟的资源状况[J]. 四川动物, 35(4): 617-621. |
张希国. 2011. 黑龙江省东方白鹳现状及种群恢复[J]. 野生动物学报, 32(3): 164-166. |
朱明畅, 曹铭昌, 汪正祥, 等. 2015. 黄河三角洲自然保护区水禽生境适宜性模糊综合评价[J]. 华中师范大学学报(自然科学版), 49(2): 287-294, 301. |
Chalfoun AD, Schmidt KA. 2012. Adaptive breeding-habitat selection: is it for the birds?[J]. The Auk, 129(4): 589-599. DOI:10.1525/auk.2012.129.4.589 |
Cheng L, Zhou LZ, Wu LX, et al. 2020. Nest site selection and its implications for conservation of the endangered oriental stork Ciconia boyciana in Yellow River Delta, China[J]. Bird Conservation International, 30(2): 323-334. DOI:10.1017/S0959270919000303 |
Cox AW, Bankovich B, Malachowski K, et al. 2019. Nest site selection by reddish egrets in Florida[J]. The Journal of Wildlife Management, 83(1): 184-191. DOI:10.1002/jwmg.21577 |
Gaillard JM, Hebblewhite M, Loison A, et al. 2010. Habitat-performance relationships: finding the right metric at a given spatial scale[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 365(1550): 2255-2265. DOI:10.1098/rstb.2010.0085 |
Shimazaki H, Tamura M, Darman Y, et al. 2004. Network analysis of potential migration routes for oriental white storks (Ciconia boyciana)[J]. Ecological Research, 19(6): 683-698. DOI:10.1111/j.1440-1703.2004.00684.x |
Wei ZH, Li YK, Xu P, et al. 2016. Patterns of change in the population and spatial distribution of oriental white storks (Ciconia boyciana) wintering in Poyang Lake[J]. Zoological Research, 37(6): 338-346. |
Yamada Y, Itagawa S, Yoshida T, et al. 2019. Predicting the distribution of released oriental white storks (Ciconia boyciana) in central Japan[J]. Ecological Research, 34(2): 277-285. DOI:10.1111/1440-1703.1063 |
Zhou LZ, Xue WW, Zhu SY, et al. 2013. Foraging habitat use of oriental white stork (Ciconia boyciana) recently breeding in China[J]. Zoological Science, 30(7): 559-564. DOI:10.2108/zsj.30.559 |