 2021, Vol. 40
2021, Vol. 40扩展功能
文章信息
- 曾键文, 张亚祖, 王超, 庆保平, 张跃明, 叶元兴, 何健
- ZENG Jianwen, ZHANG Yazu, WANG Chao, QING Baoping, ZHANG Yaoming, YE Yuanxing, HE Jian
- 朱鹮对冬季农田的觅食选择
- Foraging Habitat Selection of Nipponia nippon in Farmlands in Winter
- 四川动物, 2021, 40(5): 525-531
- Sichuan Journal of Zoology, 2021, 40(5): 525-531
- 10.11984/j.issn.1000-7083.20200471
-
文章历史
- 收稿日期: 2020-01-21
- 接受日期: 2021-06-17






2. 陕西汉中朱鹮国家级自然保护区管理局, 陕西洋县 723300;
3. 北京林业大学生态与自然保护学院, 北京 100083
2. Shaanxi Hanzhong Crested Ibis National Nature Reserve Administration, Yangxian, Shaanxi Province 723300, China;
3. School of Ecology and Nature Conservation, Beijing Forestry University, Beijing 100083, China
食物、水和隐蔽条件是影响野生动物觅食地选择的重要因素(牛翠娟等,2006;Qiao et al., 2006;蒋爱伍等,2012)。冬季大面积冰封常导致小型非啮齿类动物(啮齿动物主要通过储藏食物度过冬季)获取食物困难(牛翠娟等,2006),长期封冻甚至导致野生动物因缺乏食物而大量死亡(孙儒泳等,2001;蒋志刚等,2002)。研究发现,安徽升金湖附近越冬的白头鹤Grus monacha种群数量主要受食物资源的限制(万文静,2016)。冬季野生动物的觅食活动除了应对寒冷等恶劣环境因素,保障生存安全,还要为来年繁殖做好能量和物质储备。
朱鹮Nipponia nippon是大中型涉禽,国家一级重点保护野生动物,被世界自然保护联盟(IUCN) 濒危物种红色名录列为濒危(EN)物种。自1981年朱鹮被重新发现以来,政府部门和国内外学者为拯救这一濒危物种做了大量研究和保护工作(史东仇,曹永汉,2001;丁长青,2004;Li et al., 2017),极大促进了朱鹮种群数量的增长,分布范围逐步扩散至陕西省汉中市城固县、西乡县、汉台区、南郑区、勉县、佛坪县、宁强县、石泉县、汉阴县及太白县等地(刘冬平等,2014;胡灿实,2016;王超等,2020)。截止2019年底,朱鹮野生种群数量增长至2 600只(王超等,2020)。
朱鹮以鱼类、两栖类、甲壳类、软体动物和昆虫类为食(丁长青,刘冬平,2002;丁长青,2004)。食物是朱鹮活动和扩散的主要影响因子(胡灿实,2016)。研究发现,冬季食物不足是限制其种群发展的重要原因(路宝忠等,1989;丁长青,2004)。农田,特别是冬水田,是朱鹮冬季最主要、利用率最高的觅食地(马志军等,2001;丁长青,2004)。目前,主要通过社区共管机制组建野生朱鹮保护监测网络,引导和鼓励当地村民对冬水田进行翻犁蓄水,改善朱鹮冬季的觅食地(丁长青,刘冬平,2007;刘冬平等,2014;曾键文等,2018)。
以往研究表明,影响朱鹮对农田觅食地选择的主要因素为海拔、开阔度、水田面积、蓄水深度、明水面积、土壤松软度、植被覆盖率和人为干扰等(马志军等,2001;宋虓,2012)。朱鹮偏好在人为活动频繁但干扰强度低、土壤松软、水田面积较大、植被覆盖度低的水田中觅食(马志军等,2001;丁长青,2004;宋虓,2012)。近年来,随着朱鹮种群数量增长,种群不断扩散,其觅食环境发生了较大变化,首先是栖息地由高海拔山区转移到汉江周边的平川和低海拔丘陵地带(颜文博等,2016),其次是朱鹮分布区城镇化、生态移民和农民外出务工等社会因素对其觅食地的数量和质量造成了干扰(刘冬平等,2014)。这些变化已经对朱鹮繁殖期幼鸟的生长发育造成了影响(曾键文等,2017)。野生动物可以通过调整自身的行为和栖息地选择来适应环境因素的变化(陈水华等,2002;Guthrie et al., 2009)。研究表明,朱鹮对觅食地的偏好已逐步发生变化,倾向于在明水面积较大、有植被覆盖的水田浅水区觅食(宋虓,2012),水田面积不再是觅食地选择的影响因素(Hu et al., 2016)。在当前环境变化下,定量研究朱鹮对农田的觅食选择偏好,可以深入了解朱鹮种群的生态适应能力,为合理制定科学的栖息地保护和恢复措施提供参考。
1 研究区域研究区域为陕西汉中朱鹮国家级自然保护区,位于汉中秦岭南坡的洋县和城固县(107°21′~107°44′E,33°08′~33°35′N)境内,总面积375.49 km2,海拔400~1 200 m,属北亚热带湿润季风气候,具有温暖湿润,雨热、干冷同季的特点,地形以低山丘陵过渡地带为主,境内拥有以湑水河、溢水河和傥水河为主的众多汉江支流,塘库及附近成片的水田组成的丰富的湿地资源,是野生朱鹮主要分布地(丁长青,2004)。
2 研究方法 2.1 调查方法利用ENVI 4.0对从LANDSAT 8.0上下载的2014年TM影像进行裁剪和校准,通过支持向量机法(SVM)提取出保护区农田地类数据,运用ArcGIS 10.2矢量化功能制成保护区农田分布图,并在保护区农田分布图上随机选取12条1 000 m×50 m的样线(图 1)。农田分为未翻犁蓄水稻田(UI)、翻犁蓄水稻田(PI)、翻犁未蓄水稻田(PD)、未翻犁未蓄水稻田(UD)、旱地(DF)和撂荒地(AL)6种农田类型(表 1)。

|
| 图 1 陕西汉中朱鹮国家级自然保护区内的农田及调查样线 Fig. 1 Farmlands and sample lines in the Shaanxi Hanzhong Crested Ibis National Nature Reserve |
| |
| 农田类型 Farmland type |
定义 Definition |
| 翻犁蓄水稻田 | 稻田一季耕作结束后翻犁蓄水,有水层,无稻茬 |
| 未翻犁蓄水稻田 | 稻田一季耕作结束后只蓄水,有水层,有稻茬 |
| 翻犁未蓄水稻田 | 稻田一季耕作结束后只翻犁,无水层,无稻茬 |
| 未翻犁未蓄水稻田 | 稻田一季耕作结束后未翻犁且未蓄水,无水层,有稻茬 |
| 旱地 | 进行旱作的农田,包括小麦地、油菜地、蔬菜地及其他旱生作物种植的农田 |
| 撂荒地 | 本年未被耕作、管理,而形成退化和自然演替的荒地 |
调查时间为2016年11月—2017年2月、2017年11月—2018年2月,每条样线隔20 d调查1次,每条样线累计调查10次。以约1.8 km·h-1速度在样线上步行,统计样线区域内6种农田内觅食朱鹮的数量。研究人员使用手持式GPS仪(GARMIN GPSMAP60CSx)和手持式激光测距仪(SNDWAY SW-900A,5~900 m)辅助测定不同类型农田面积。待朱鹮从觅食水田离开后,随机在朱鹮觅食的田块选5个点,用钢卷尺(精度1 mm)测量水深,平均测量3次,去掉最大值和最小值,取平均值作为水田蓄水深度。另外,在7条朱鹮活动频繁的样线上随机抽取28个样点,用长100 cm×宽100 cm×高40 cm的自制取样器,按样点所在田块的面积取1~4个样方,取水层和10 cm深土壤层体长大于5 mm的生物作为样本,进行农田底栖动物丰富度调查,每年调查1次,累计调查3次。生物样本按《中国土壤动物检索图鉴》(尹文英,1998)和《普通动物学(第4版)》(刘凌云,郑光美,2009)处理。
2.2 数据处理方法通过Vanderploeg和Scavia(1979a, 1979b)选择系数(Wi)和选择指数(Ei)作为衡量朱鹮对觅食地喜好程度的指标。公式如下:
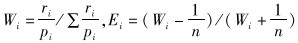
|
式中,i为农田类型,n为农田类型总数;ri为选择农田类型i觅食的朱鹮占所有朱鹮数量的比例,pi为环境中农田类型i的可利用面积占所有农田的比例。Ei的数值在-1到1之间,若-1≤Ei<0表示回避农田类型i,Ei=0表示随机选择,0<Ei≤1表示偏好农田类型i。利用Kruskal-Wallis非参数检验对朱鹮在不同类型农田上选择性以及不同农田物种多样性指数进行差异性分析(Kruskal & Wallis,1952)。
随机选取的28个生物资源调查样点,由于翻犁未蓄水的稻田、未翻犁未蓄水的稻田、旱地和撂荒地土地质地坚硬,并且多种植蔬菜等原因,无法有效采集底栖动物,最终采集了未翻犁蓄水稻田(n=11)和翻犁蓄水稻田(n=13)的底栖动物数据,这2种农田类型在本次调查中发现朱鹮的比例为84.78%。利用Shannon-Wiener指数计算2种蓄水农田类型底栖动物的生物多样性,并计算底栖动物密度。利用Mann-Whitney U非参数检验对比2种蓄水农田的底栖动物密度和生物多样性差异。所有数据均在SPSS中进行统计分析,显著性水平设置为α=0.05。
3 结果 3.1 朱鹮觅食栖息地选择朱鹮对各类农田的选择系数和选择指数具有显著性差异,偏好选择未翻犁蓄水稻田(Ei=0.49± 0.11);回避选择翻犁蓄水稻田(Ei=-0.49±0.25)。翻犁未蓄水稻田、未翻犁未蓄水稻田、旱地和撂荒地的选择指数分别为-1.00±0、-0.86±0.14、-0.71±0.20和-0.94±0.06(表 2)。
| 项目 Item |
农田类型Farmland type | Kruskal-Wallis检验 | |||||
| 翻犁蓄水稻田 Ploughed and impounding land |
未翻犁蓄水稻田 Unploughed and impounding land |
翻犁未蓄水稻田 Ploughed and dry land |
未翻犁未蓄水稻田 Unploughed and dry land |
旱地 Dry farming land |
撂荒地 Abandoned land |
||
| 可利用率/% | 4.60±1.44 | 23.78±4.87 | 5.92±1.64 | 17.38±5.84 | 35.57±8.04 | 12.75±4.79 | χ2=19.451, P=0.002 |
| 实际利用率/% | 4.31±2.40 | 81.50±7.71 | 0±0 | 0.85±0.85 | 8.89±6.11 | 4.44±4.44 | χ2=35.040, P<0.001 |
| 选择系数Wi | 0.18±0.09 | 0.66±0.11 | 0±0 | 0.03±0.03 | 0.08±0.06 | 0.01±0.01 | χ2=31.568, P<0.001 |
| 选择指数Ei | -0.49±0.25 | 0.49±0.11 | -1.00±0 | -0.86±0.14 | -0.71±0.20 | -0.94±0.06 | χ2=31.561, P<0.001 |
朱鹮对农田蓄水深度的选择具有显著的差异性(表 3)。朱鹮多选择在蓄水深度3~6 cm的水田内觅食,选择指数为0.16±0.21;不选择0 cm、0~ 3 cm、6~10 cm、10~15 cm和>15 cm的农田,选择指数分别为-0.94±0.58、-0.09±0.26、-0.46±0.28、-0.85±0.15和-1.00±0(表 3)。
| 项目 Item |
蓄水深度Water storage depth/cm | Kruskal-Wallis检验 | |||||
| 0 | 0~3 | 3~6 | 6~10 | 10~15 | >15 | ||
| 可利用率/% | 48.43±9.95 | 26.73±5.56 | 9.74±3.85 | 21.56±11.32 | 1.38±1.18 | 0±0 | χ2=27.619, P<0.001 |
| 实际利用率/% | 0.96±0.96 | 48.01±12.54 | 39.39±10.70 | 19.62±9.25 | 1.28±1.28 | 0±0 | χ2=21.723, P=0.001 |
| 选择系数Wi | 0.01±0.01 | 0.41±0.15 | 0.47±0.15 | 0.20±0.13 | 0.02±0.02 | 0±0 | χ2=20.196, P=0.001 |
| 选择指数Ei | -0.94±0.58 | -0.09±0.26 | 0.16±0.21 | -0.46±0.28 | -0.85±0.15 | -1.00±0 | χ2=20.196, P=0.001 |
分析底栖动物的食物样方,发现水田中底栖动物以昆虫为主,占总生物数量的64.02%(表 4)。通过统计检验发现,未翻犁蓄水田和翻犁蓄水田底栖动物的生物多样性和密度均没有显著差异(U检验:P>0.05;表 4)。
| 项目Item | 未翻犁蓄水稻田 Unploughed and impounding land (n=11) |
翻犁蓄水稻田 Ploughed and impounding land (n=13) |
累计Total | 占比Percentage/% |
| 两栖类数量/只 | 4 | — | 4 | 0.59 |
| 田螺数量/只 | 4 | 40 | 44 | 6.46 |
| 昆虫数量/只 | 301 | 135 | 436 | 64.02 |
| 其他 | 127 | 70 | 197 | 28.93 |
| Shannon-Wiener多样性指数 | 3.55±0.08 | 3.49±0.49 | — | |
| 食物密度/(只/m2) | 29.1±11.8 | 17.4±5.5 | — | |
| 注:"其他" 以水丝蚓等环节动物和蛛形纲为主 Note:"Others" are mainly Annelids and Arachnids | ||||
以往研究发现,朱鹮冬季的觅食地主要为水田,地形因素(海拔)、水田面积和人为干扰强度是影响朱鹮冬季觅食地选择的重要因素(马志军等,1999)。随着野生朱鹮种群数量增长,朱鹮觅食地重叠、局部地区环境容纳量下降(曾键文等,2017)。与2000年前相比,朱鹮的觅食地环境发生了较大变化,近些年朱鹮个体偏好在觅食地土壤更软、植物郁闭度更大、蓄水深度更浅的水田上觅食,并且水田面积不再是朱鹮觅食地选择的限制性因素(Hu et al., 2016)。
本研究发现,水田仍然是朱鹮冬季最重要的觅食地,且倾向于在不翻犁的冬水田中觅食。未蓄水的稻田、旱地和撂荒地土壤质地比较坚硬,不是朱鹮冬季理想的觅食地。这也与水域内动物是朱鹮食物主要组成的习性有关(史东仇,曹永汉,2001)。在洋县,每年冬季农田进行冬灌期间都吸引了大量朱鹮在其中觅食。翻犁田底栖动物生物多样性和密度与不翻犁田均没有显著差异,这与陕西省宁陕地区放归朱鹮栖息地评价的结果一致(赵洪峰等,2010),可能是因为翻耕后,朱鹮可以取食一些如鳞翅目Lepidoptera和蜻蜓目Odonata的昆虫幼虫,但是也造成了一些分布于地表的成体昆虫翻到下层难以获取。
水深是影响水鸟觅食地选择的重要因素(叶元兴等,2012)。朱鹮偏好蓄水深度为3~6 cm冬水田。一方面,一定深度的蓄水水田中,水生植物、稻茬露出水面,水层作为底栖动物捕食、活动场所,泥层可以为底栖动物躲避敌害,水田生物在水上部分、水层和泥层间往返活动,构成了稳定完善、多度和丰富度较高的冬季水田小环境(冯韵等,2014);另一方面,朱鹮跗趾长8.5~9 cm,喙长18 cm,这样的体征限制了其觅食的水深(范光丽等,1999)。朱鹮是靠触觉觅食的涉禽(Ye et al., 2017)。冬季水田生物如蛙类Rana spp.、泥鳅Misgurnus anguillicaudatus和黄鳝Monopterus albus等多处于冬眠状态,昆虫的成虫和幼虫是朱鹮秋冬季主要食物(史东仇,曹永汉,2001;赵洪峰等,2010)。因此,近年来保护区开展的农田冬季蓄水、引导农民一季耕作等湿地修复措施为朱鹮种群安全越冬提供了重要的觅食地。
本研究建议在今后保护管理过程中,一是继续保持稻田冬季适宜深度的蓄水,加强蓄水深度精细化管理;二是根据当前朱鹮野生种群增长以及扩散区域,应该针对性扩大蓄水水田面积。
陈水华, 丁平, 范忠勇, 等. 2002. 城市鸟类对斑块状园林栖息地的选择性[J]. 动物学研究, 23(1): 31-38. DOI:10.3321/j.issn:0254-5853.2002.01.006 |
丁长青, 刘冬平. 2002. 朱鹮(Nipponia nippon)[J]. 动物学杂志, 37(2): 84-85. DOI:10.3969/j.issn.0250-3263.2002.02.026 |
丁长青, 刘冬平. 2007. 野生朱鹮保护研究进展[J]. 生物学通报, 42(3): 1-4. DOI:10.3969/j.issn.0006-3193.2007.03.001 |
丁长青. 2004. 朱鹮研究[M]. 上海: 上海高等教育出版社.
|
范光丽, 吴建云, 马新武. 1996. 中国朱鹮骨骼解剖学观察[J]. 西北农业大学学报, 24(3): 72-76. DOI:10.3321/j.issn:1671-9387.1996.03.016 |
冯韵, 郑雪莉, 卜书海, 等. 2014. 陕西宁陕县朱鹮觅食地秋季食物丰富度调查[J]. 陕西林业科技, 1: 1-4. |
胡灿实. 2016. 朱鹮(Nipponia nippon)活动性及其扩散模式研究[D]. 北京: 北京林业大学.
|
蒋爱伍, 周放, 覃玥, 等. 2012. 中国大陆鸟类栖息地选择研究十年[J]. 生态学报, 32(18): 5918-5923. |
蒋志刚, 李春旺, 曾岩. 2002. 中国野生动物研究进展[C]. 中国生物多样性保护与研究进展——第五届全国生物多样性保护与持续利用研讨会论文集. 北京: 气象出版社.
|
刘冬平, 王超, 庆保平, 等. 2014. 朱鹮保护30年——基于社区的极小种群野生动物保护典范[J]. 四川动物, 33(4): 612-619. |
刘凌云, 郑光美. 2009. 普通动物学(第4版)[M]. 北京: 高等教育出版社.
|
路宝忠. 1989. 朱鹮的人工投食[J]. 野生动物, 5: 23-24. |
马志军, 丁长青, 李欣海, 等. 2001. 朱鹮冬季觅食地的选择[J]. 动物学研究, 22(1): 46-50. DOI:10.3321/j.issn:0254-5853.2001.01.007 |
牛翠娟, 娄安如, 孙儒泳, 等. 2006. 基础生态学(第二版)[M]. 北京: 高等教育出版社.
|
史东仇, 曹永汉. 2001. 中国朱鹮[M]. 北京: 中国林业出版社.
|
宋虓. 2012. 野生朱鹮种群的觅食地选择研究[D]. 北京: 北京林业大学.
|
孙儒泳, 李博, 诸葛阳, 等. 2001. 普通生态学[M]. 北京: 高等教育出版社.
|
万文静. 2016. 升金湖越冬白头鹤(Grus monacha)种群环境容纳量研究[D]. 合肥: 安徽大学.
|
王超, 张亚祖, 曾键文, 等. 2020. 中国野生朱鹮的繁殖现状和种群数量[J]. 林业科学, 56(11): 165-172. |
颜文博, 王琦, 王超. 2016. 应用Maxent模型分析秦巴地区朱鹮适宜繁殖地的分布[J]. 动物学杂志, 50(2): 185-193. |
叶元兴, 王楠, 丁长青. 2012. 鹮嘴鹬越冬觅食地选择与食物资源调查[J]. 动物学杂志, 47(2): 46-51. |
尹文英. 1998. 中国土壤动物检索图鉴[M]. 北京: 科学出版社.
|
曾键文, 庆保平, 路晋, 等. 2017. 陕西汉中野生朱鹮雏鸟的生长发育特征[J]. 动物学杂志, 52(5): 777-782. |
曾键文, 权海云, 庆保平, 等. 2018. 野生朱鹮秋季种群数量调查评价方法初探[J]. 野生动物学报, 39(2): 329-333. DOI:10.3969/j.issn.1000-0127.2018.02.019 |
赵洪峰, 罗磊, 常秀云, 等. 2010. 陕西宁陕野化放飞朱鹮秋季觅食地选择与食物丰富度[J]. 动物学杂志, 45(1): 83-89. |
Guthrie S, Zimmerman RJ, Gutiérrez WE, et al. 2009. Multiscale habitat selection by ruffed grouse at low population densities[J]. The Condor, 111(2): 294-304. DOI:10.1525/cond.2009.080036 |
Hu CS, Song X, Ding CQ, et al. 2016. The size of winter-flooded paddy fields no longer limits the foraging habitat use of the endangered crested ibis (Nipponia nippon) in Winter[J]. Zoological Science, 33(4): 345-351. DOI:10.2108/zs150012 |
Kruskal WH, Wallis WA. 1952. Use of ranks in one-criterion variance analysis[J]. Journal of the American Statistical Association, 47(260): 583-621. DOI:10.1080/01621459.1952.10483441 |
Li YF, Ye XP, Wang M, et al. 2017. Survival rates of a reintroduced population of the crested ibis (Nipponia nippon) in Ningshan County (Shaanxi, China)[J]. Bird Conservation International, 28(1): 145-156. |
Qiao JF, Yang WK, Gao XY. 2006. Natural diet and food habitat use of the Tarim red deer, Cervus elaphus yarkandensis[J]. Chinese Science Bulletin, 51: 147-152. |
Vanderploeg H, Scavia D. 1979a. Two selectivity indices for feeding with special reference to zooplankton grazing[J]. Journal of the Fisheries Research Board of Canada, 36(4): 362-365. DOI:10.1139/f79-055 |
Vanderploeg H, Scavia D. 1979b. Calculation and use of selectivity coefficients of feeding: zooplankton grazing[J]. Ecology Modeling, 7(2): 135-149. DOI:10.1016/0304-3800(79)90004-8 |
Ye Y, Jiang Y, Hu C, et al. 2017. What makes a tactile forager join mixed-species flocks? A case study with the endangered crested ibis (Nipponia nippon)[J]. The Auk, 134(2): 421-431. DOI:10.1642/AUK-16-191.1 |