 2021, Vol. 40
2021, Vol. 40扩展功能
文章信息
- 苏英钰, 黎敏灵, 程彬彬, 刘洁, 黄慧琴, 徐良华, 左文波, 张微微
- SU Yingyu, LI Minling, CHENG Binbin, LIU Jie, HUANG Huiqin, XU Lianghua, ZUO Wenbo, ZHANG Weiwei
- 江西官山国家级自然保护区白鹇活动节律及时间分配研究
- Activity Rhythm and Time Budget of Lophura nycthemera in the Guanshan National Nature Reserve, Jiangxi
- 四川动物, 2021, 40(5): 517-524
- Sichuan Journal of Zoology, 2021, 40(5): 517-524
- 10.11984/j.issn.1000-7083.20200446
-
文章历史
- 收稿日期: 2020-11-27
- 接受日期: 2021-05-20






2. 江西官山国家级自然保护区管理局, 江西宜春 336330
2. Administration Bureau of Jiangxi Guanshan National Nature Reserve, Yichun, Jiangxi Province 336330, China
行为是指动物面对外界环境和体内环境变化而产生的外在反应(尚玉昌,2005),是动物长期进化的结果。同时,动物会不断调整行为以适应周围环境和自身生理状况的改变,进而形成特定条件下的时间分配和活动节律模式(Flannigan & Stookey, 2002;Everts,2004; 夏参军等,2009),这2种模式是研究动物生存策略、动物行为及生态环境关系的重要基础(孙儒泳,2001),并对保护区制定管理和保护野生动物措施具有重要意义。
白鹇Lophura nycthemera隶属于鸡形目Galliformes雉科Phasianidae,是国家二级重点保护野生动物,主要分布于我国福建、浙江、江西、安徽、湖南等省份(邵晨,胡一中,2005;吴友贵等,2017),主要栖息于亚热带季风常绿阔叶林、沟谷雨林、针阔混交林及竹林等树木茂密、林下灌丛稀少且靠近水源的生境(丁平等,1992;高育仁,1996;熊志斌等,2003)。官山国家级自然保护区拥有茂密的森林植被,多样的生态环境符合白鹇种群栖息地选择的要求(林大昌,林坚,2014;游卫云等,2019),白鹇种群数量较多(刘信中,2005),是保护区的主要保护对象之一。但目前对保护区的白鹇种群研究集中在种群数量(戴年华等,1997)、生境选择(刘鹏等,2012;陈俊豪等,2019)等方面,尚缺乏行为节律和时间分配的研究,不利于掌握区内白鹇的种群动态。白鹇警惕性高、反应敏捷,难以利用传统方法(如样线法)对其活动节律和时间分配状况展开调查。红外触发相机陷阱技术具有人工成本相对较低、准确性高、无创、对环境干扰小以及可探测到活动隐蔽、警惕性高的物种等优点(Karanth & Nichols,1998;Silveira et al., 2003)。因此,本研究基于红外相机技术对保护区白鹇的活动节律及时间分配进行分析,探讨其对保护区生境的适应策略,从而为该物种的保护和管理提供科学依据。
1 研究区域和研究方法 1.1 研究区域概况江西官山国家级自然保护区位于江西省铜鼓县、宜丰县九岭山脉中部,地理位置114°29′~114°45′E,28°30′~28°40′N,总面积11 500.5 hm2,海拔200~ 1 480 m。属中亚热带温暖湿润气候区,年均气温16.2 ℃,无霜期长,年均降水量1 950~2 100 mm (刘鹏等,2011)。主要植被类型有常绿阔叶林、山顶矮林、竹林、山顶灌草丛、沼泽植被等10个类型,是典型的亚热带森林生态系统(林大昌,林坚,2014)。
1.2 研究方法2015年1月—2016年8月,根据生境类型、海拔、人为活动及白鹇的活动习性等因素,在保护区内先后布设28个红外相机监测位点,相机主要固定在树干上距地面45~100 cm的位置,镜头略倾向于地面,海拔每100 m布置1台红外相机,同时记录布置时间、经纬度及生境。相机型号为猎科Ltl-5210,参数设置为工作时间24 h,触发间隔时间1 s,敏感度中等,自动连续编码,每次触发为拍摄照片3张后录制10~15 s视频,每隔3个月检查各相机是否需要更换存储卡、电池,以及是否需要更换位置以获取更多的白鹇种群动态信息。
1.3 数据分析根据动物行为谱定义习惯(赵玉泽等,2013;唐创斌等,2020),将拍摄资料中白鹇日常行为归类为4种:(1)觅食:移动时啄取地面食物或停下扒掘土取食;(2)移动:缓慢或急速地行走,飞行;(3)梳理:用喙啄理羽毛、用爪搔抓头部及颈前部分、用喙啄跗蹠及足、挺胸拍翅、抖羽等;(4)静息:整体基本不动,局部无明显活动,包括躺卧静栖、蹲伏静栖和站立静栖。所有行为记录频次,如一段视频中有2种及以上行为,则每种行为记录1次。
对所获得的红外相机数据按相机编号分类,统计各相机拍摄到的白鹇数量、性别、行为和拍摄时间等信息。单台红外相机在野外连续工作24 h记为1个有效相机工作日;对于连续拍摄的白鹇照片(视频视为照片),为降低同一物种照片的自相关性,将同一相机位点的拍摄时间间隔小于30 min的连续相同物种照片算作1张独立有效照片(Masatoshi,2004;李佳等,2017)。以100 m将相机监测位点的海拔划分为300~400 m、400~ 500 m、500~ 600 m、600~700 m、700~800 m、800~900 m、1 000~ 1 100 m和1 100~1 200 m。
红外相机拍摄率(CR)(赵玉泽等,2013):
通过时间段活动强度指数(time-period activity intensity index,TAII)分析白鹇的日活动节律及其季节和性别差异(赵玉泽等,2013;唐卓等,2017):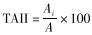
年行为时间分配频次比例(Pk)与季行为时间分配频次比例(Pj)(王海婴,2017):

分析白鹇性别差异时,首先采用Kolmogorov-Smirnov Z-test检验数据的正态性,符合正态分布的数据采用独立样本t检验;不符合正态分布的经转换后,再次进行正态性检验(刘鹏,张微微,2017)。用One-Way ANOVA分析季节差异性,多重比较使用Tukey法。所有数据利用SPSS 19.0和Excel 2013处理,显著性水平设为α=0.05。
2 结果与分析 2.1 白鹇拍摄概况28个红外相机监测位点均拍摄到白鹇,累计监测5 643个有效相机日,共获得1 316张白鹇独立有效照片,合计1 801只次;成体及亚成体雄性共587只次,雌性共785只次,雌雄比例为1.34∶ 1;幼体及初期亚成体共429只次。
不同海拔布设的红外相机拍摄到的白鹇频次不同,白鹇的集中活动区为海拔600~700 m和900~1 200 m(表 1)。红外相机拍摄数量最多的月份是6月和7月,共583只次,拍摄率分别为68.45%和52.42%(表 2),雌雄间拍摄率的差异无统计学意义(t22=0.916,P=0.370)。
| 红外相机布设海拔 Altitude of infrared cameras/m |
红外相机数量 Number of infrared cameras |
独立有效照片 Number of independent effective photos |
平均照片数 Average number of photos |
| 300~400 | 1 | 3 | 3 |
| 400~500 | 4 | 18 | 4.5 |
| 500~600 | 4 | 98 | 24.5 |
| 600~700 | 3 | 168 | 56 |
| 700~800 | 2 | 15 | 7.5 |
| 800~900 | 2 | 85 | 42.5 |
| 900~1 000 | 2 | 199 | 99.5 |
| 1 000~1 100 | 6 | 318 | 53 |
| 1 100~1 200 | 4 | 412 | 103 |
| 月份 Month |
有效相机日 Effective day |
曝光 Triggers |
个体数 Number of individuals/只次 |
拍摄率 Capture rate/% |
| 1月 | 324 | 17 | 28 | 8.64 |
| 2月 | 681 | 120 | 202 | 29.66 |
| 3月 | 714 | 104 | 135 | 18.91 |
| 4月 | 687 | 190 | 223 | 32.46 |
| 5月 | 590 | 187 | 215 | 36.44 |
| 6月 | 504 | 251 | 345 | 68.45 |
| 7月 | 454 | 178 | 238 | 52.42 |
| 8月 | 424 | 104 | 120 | 28.30 |
| 9月 | 360 | 41 | 45 | 12.50 |
| 10月 | 345 | 62 | 90 | 26.09 |
| 11月 | 281 | 38 | 70 | 24.91 |
| 12月 | 279 | 24 | 90 | 32.26 |
| 总计 | 5 643 | 1 316 | 1 801 |
从图 1可看出,白鹇活动集中在04∶ 00—20∶ 00,活动高峰为06∶ 00—08∶ 00与12∶ 00—14∶ 00。雌雄白鹇均有2个日活动高峰,第一个活动高峰都出现在06∶ 00—08∶ 00,第二个活动高峰为雌鸟(12∶ 00—14∶ 00)、雄鸟(10∶ 00—12∶ 00);雌雄白鹇在各时间段内的活动强度之间的差异无统计学意义(t22=0.979,P=0.338)。春、夏和秋季在06∶ 00—08∶ 00均出现第一个较为明显的活动高峰(图 2);第二个活动高峰为春季(12∶ 00—14∶ 00),夏季与秋季(14∶ 00—16∶ 00);而冬季仅在12∶ 00—14∶ 00出现1个活动高峰,相比其他3季的晚6 h。不同季节的日活动强度间的差异有统计学意义(F3=4.238,P=0.010)。觅食高峰期为10∶ 00—12∶ 00,移动高峰期为06∶ 00—08∶ 00和14∶ 00—16∶ 00(图 3),而静息与梳理行为因拍摄率较低,未做分析。

|
| 图 1 江西官山国家级自然保护区白鹇日活动节律和性别差异 Fig. 1 Diurnal behavioral rhythms and sex difference of Lophura nycthemera in the Guanshan National Nature Reserve, Jiangxi |
| |

|
| 图 2 江西官山国家级自然保护区白鹇日活动节律的季节性差异 Fig. 2 Seasonal differences of daily behaviors rhythm of Lophura nycthemera in the Guanshan National Nature Reserve, Jiangxi |
| |

|
| 图 3 江西官山国家级自然保护区白鹇日行为时间分配 Fig. 3 Time budget of daily behaviors of Lophura nycthemera in the Guanshan National Nature Reserve, Jiangxi |
| |
各行为时间分配比例依次为移动(61.71%)、觅食(37.01%)、静息(0.88%)和梳理(0.40%)。白鹇随季节变化会不断调整自身主要行为活动及时间分配,主要表现为:移动行为在春季的时间分配比例为71.52%、夏季60.87%、秋季53.00%、冬季41.10%;觅食行为在春季的时间分配比例为27.85%、夏季38.54%、秋季45%、冬季53.99%;但不同季节的觅食(F3=0.813,P=0.494)与移动行为(F3=0.680,P=0.569)之间的差异均无统计学意义(图 4)。

|
| 图 4 江西官山国家级自然保护区白鹇不同季节的行为时间分配 Fig. 4 Seasonal differences of behaviors' time budget of Lophura nycthemera in the Guanshan National Nature Reserve, Jiangxi |
| |
江西官山国家级自然保护区白鹇雌雄比为1.34∶ 1,高于王志淦等(2019)在江西齐云山国家级自然保护区的调查结果(1.08∶ 1.00),低于广西弄岗国家级自然保护区(2.62∶ 1.00)(刘佳等,2019)。一方面可能与红外相机布设的主要季节不同有关,另一方面,可能不同地区的白鹇雌雄比确实存在差异。根据王志淦等(2019)的研究,齐云山保护区白鹇的主要行为为警戒,并且日活动强度分析发现,警戒行为在每个时间段均占最大比例,表明白鹇在该地区有较大的被捕食风险。而弄岗保护区白鹇的警戒行为仅次于移动与觅食行为,表明白鹇在该地区的被捕食风险也很高,该地为喀斯特石山地貌,雄性白鹇体型较大且羽色鲜艳,在裸露的岩石中容易被发现,因此营集群生活的白鹇可能为降低被捕食风险而减少群体中的雄性个体(刘佳等,2019)。权衡被捕食风险而倾向以单性集群的方式作为反捕食对策,也见于白冠长尾雉Syrmaticus reevesii等鸟类(张晓辉等,2004;赵玉泽等,2016)。
本研究结果表明白鹇主营昼间生活,而夜间活动较少被记录,这与其夜栖行为有关(邵晨,胡一中,2005)。由于红外相机多设于兽径,固定高度一般为50~100 cm,无法拍摄到树栖鸟类(朱淑怡等,2017)。同样,白颈长尾雉S. ellioti(丁平等,2002)、白冠长尾雉(孙全辉等,2002)、白腹锦鸡Chrysolophus amherstiae(康明江,郑光美,2007)、红腹角雉Tragopan temminckii(丛培昊,郑光美,2008)、黑颈长尾雉S. humiae(贝永建等,2009;原宝东等,2017)等雉类也是以昼间活动为主,夜栖于乔木盖度较高的环境。雉类为日行性鸟类,有2个活动高峰,普遍集中于晨昏(06∶ 00—09∶ 00、17∶ 00—19∶ 00),如石鸡Alectoris chukar(周晓禹等,2008)、红腹锦鸡C. pictus(刘小斌等,2017)、白眉山鹧鸪Arborophila gingica和灰胸竹鸡Bambusicola thoracicus(陈立军等,2019)、原鸡Gallus gallus(王方等,2020)、绿孔雀Pavo muticus(晏鸣霄等,2021)等,均表现出相似的晨昏活动模式,而官山保护区白鹇的活动高峰则为清晨及午后,可能与其生存环境有关。鸟类在不同的环境下会表现出不同的行为节律(曾宾宾等,2013),这可能与各地区的温度差异有关。同样利用红外相机技术,有研究发现白鹇活动集中于午间(李生强等,2016;徐建国等,2019)、晨午昏(余建平等,2017;王志淦等,2019)或晨昏(王方等,2020)3个时间段,说明白鹇的行为表现出更大的弹性。此外,本研究表明白鹇的活动高峰存在季节差异,第一个活动高峰全年均出现在07∶ 00,即日出后1 h左右,春季和秋季的第二个高峰出现在15∶ 00,夏季和冬季的则出现在13∶ 00,可能与不同季节食物的可获得性不同有关,食物丰富的季节白鹇倾向于推迟觅食。白鹇日活动强度季节之间存在显著差异,表现为春季与秋季午间出现的活动高峰比日出前后更活跃,而夏季是日出前后出现活动高峰;其他季节均在午后出现1个高峰,且比第一个高峰晚6 h左右,但峰值更高,说明白鹇活动节律随季节温度的变化而变化。野生动物常常通过降低活动频率以躲避高温或低温造成的能量浪费(施泽攀,2015;潘丹等,2019);江西属温暖带多雨气候,主要特征为冬冷、春寒、夏热(7、8月气温最高)和秋干(陆晴等,2019),因此白鹇活动表现为夏季明显回避高温,冬季回避低温,类似活动节律在勺鸡Pucrasia macrolopha(韩芬茹,周天林,2005)、血雉Ithaginis cruentus(赵联军等,2020)等雉类研究中均有出现。
白鹇各行为时间分配比例依次为移动(61.71%)、觅食(37.01%)、静息(0.88%)和梳理(0.40%),以移动和觅食行为为主。由于不同季节的食物资源不同,动物个体会根据能量需求采取最优行为时间分配策略,尤其是在食物紧缺的冬季,白鹇需要更多时间觅食以满足自身营养的需要(Pépin,2006;连新明等,2012),觅食时间随季节的食物丰富度变化而变化;移动行为在春季所占的时间分配比例最高,夏季次之,冬季最低,与觅食行为在季节变化上呈负相关。王志淦等(2019)的研究结果亦出现同样现象。说明白鹇在食物丰富度高的时候更愿意投入能量到移动等消耗性行为,而在食物缺乏时,则减少移动能量投入。此外,春季是白鹇的主要繁育期,雌雄需通过更大范围的活动获得配偶以及抚育后代,可能是白鹇对栖息地环境的一种行为适应和生存策略。
结合样线法和样方法,刘鹏等(2012)发现官山国家级自然保护区的白颈长尾雉、勺鸡、灰胸竹鸡和白鹇等4种雉类在海拔200~1 400 m分布,其中白鹇几乎分布在保护区的各个海拔梯度,集中分布在200~600 m及1 000~1 200 m,与陈俊豪等(2009)采用同种方法发现白鹇集中分布区为海拔300~500 m和1 000~1 200 m的结论类似,而本研究发现,白鹇主要在600~700 m和900~1 200 m的海拔活动,2种调查方法的结论有差异。说明红外相机调查受相机布设位置及抽样强度的影响。因此,为更全面掌握白鹇的种群现状,应合理使用红外相机技术,并结合其他调查方法,以消除单一调查方法可能存在的误差。
贝永建, 陈伟才, 李汉华, 等. 2009. 再引入黑颈长尾雉夜栖行为与夜栖地选择[J]. 四川动物, 28(1): 39-43. |
陈俊豪, 黄晓凤, 鲁长虎, 等. 2009. 白颈长尾雉与白鹇秋冬季空间生态位比较[J]. 生态学杂志, 28(12): 2546-2552. |
陈立军, 束祖飞, 肖治术. 2019. 应用红外相机数据研究动物活动节律——以广东车八岭保护区鸡形目鸟类为例[J]. 生物多样性, 27(3): 266-272. |
丛培昊, 郑光美. 2008. 四川老君山地区红腹角雉的夜栖行为和夜栖地选择[J]. 生物多样性, 16(4): 332-338. DOI:10.3321/j.issn:1005-0094.2008.04.003 |
戴年华, 刘玮, 蔡汝林. 1997. 江西省官山自然保护区鸟类调查初报[J]. 江西科学, 15(4): 243-246. |
丁平, 姜仕仁, 石斌山, 等. 1992. 浙江西部山区白鹇生态的初步研究[J]. 动物学杂志, 27(3): 20-23. |
丁平, 杨月伟, 李智, 等. 2002. 白颈长尾雉的夜宿地选择研究[J]. 浙江大学学报: 理学版, 29(5): 564-568. DOI:10.3321/j.issn:1008-9497.2002.05.015 |
高育仁. 1996. 鼎湖山保护区白鹇的季节活动和结群行为[J]. 动物学报, (S1): 74-79. |
韩芬茹, 周天林. 2005. 笼养勺鸡非繁殖季节时间分配和活动节律[J]. 河西学院学报, 21(2): 45-47. DOI:10.3969/j.issn.1672-0520.2005.02.016 |
康明江, 郑光美. 2007. 白腹锦鸡(Chrysolophus amherstiae)的夜栖地选择[J]. 生态学报, 27(7): 2929-2934. DOI:10.3321/j.issn:1000-0933.2007.07.033 |
李佳, 刘芳, 李迪强, 等. 2017. 基于红外相机监测分析的红腹角雉日活动节律[J]. 林业科学, 53(7): 170-176. |
李生强, 汪国海, 施泽攀, 等. 2016. 红外相机技术监测喀斯特生境兽类和鸟类多样性及活动节律[J]. 兽类学报, 36(3): 272-281. |
连新明, 李晓晓, 颜培实, 等. 2012. 夏季可可西里雌性藏原羚行为时间分配及活动节律[J]. 生态学报, 32(3): 663-670. |
林大昌, 林坚. 2014. "生态孤岛": 官山国家级自然保护区的由来和发展[J]. 长江丛刊, (17): 11-15. |
刘佳, 李生强, 汪国海, 等. 2019. 喀斯特生境中白鹇的活动节律、时间分配及集群行为[J]. 广西师范大学学报(自然科学版), 37(3): 156-165. |
刘鹏, 顾署生, 黄晓凤, 等. 2011. 官山自然保护区勺鸡冬季栖息地选择[J]. 江西农业大学学报, 33(6): 1257-1262. DOI:10.3969/j.issn.1000-2286.2011.06.040 |
刘鹏, 黄晓凤, 顾署生, 等. 2012. 江西官山自然保护区四种雉类的生境选择差异[J]. 动物学研究, 33(2): 170-176. |
刘鹏, 张微微. 2017. 官山自然保护区白颈长尾雉季节性生境选择[J]. 生态学报, 37(18): 6005-6013. |
刘小斌, 韦伟, 郑筱光, 等. 2017. 红腹锦鸡和红腹角雉活动节律——基于红外相机监测数据[J]. 动物学杂志, 52(2): 194-202. |
刘信中. 2005. 江西官山自然保护区科学考察与研究[M]. 北京: 中国林业出版社.
|
陆晴, 闫冰, 赵东升. 2019. 1961-2016年江西省气候时空变化特征[J]. 水土保持研究, 26(5): 166-173. |
潘丹, 吴炳贤, 张冰, 等. 2019. 武陵源世界自然遗产地兽类和鸟类多样性的红外相机初步监测[J]. 兽类学报, 39(2): 209-217. |
尚玉昌. 2005. 动物生态学[M]. 北京: 北京大学出版社.
|
邵晨, 胡一中. 2005. 白鹇的夜栖息地选择及夜栖息行为[J]. 浙江林学院学报, 22(5): 562-565. |
施泽攀. 2015. 基于红外相机对弄岗森林动态监测样地及周边的鸟兽多样性及生态学研究[D]. 广西: 广西师范大学.
|
孙全辉, 张正旺, 朱家贵, 等. 2002. 白冠长尾雉冬季夜栖行为与夜栖地利用影响因子的研究[J]. 北京师范大学学报(自然科学版), 38(1): 108-112. |
孙儒泳. 2001. 动物生态学原理[M]. 北京: 北京师范大学出版社.
|
唐创斌, 汪国海, 施泽攀, 等. 2020. 基于红外相机技术的赤腹松鼠活动节律和时间分配分析[J]. 广西师范大学学报(自然科学版), 38(1): 133-139. |
唐卓, 杨建, 刘雪华, 等. 2017. 利用红外相机研究卧龙国家级自然保护区绿尾虹雉的活动规律[J]. 四川动物, 36(5): 582-587. |
王方, 叶思达, 汤永晶, 等. 2020. 云南哀牢山南段原鸡与白鹇活动节律及种间关系[J]. 野生动物学报, 41(3): 641-647. |
王海婴. 2017. 福建武夷山赤麂(Muntiacus vaginalis)活动节律和行为时间分配的研究[D]. 福建: 厦门大学.
|
王志淦, 李言阔, 钟毅峰, 等. 2019. 基于红外相机的白鹇集群模式与活动节律研究[J]. 江西师范大学学报(自然科学版), 43(6): 592-597. |
吴友贵, 叶珍林, 吴义松, 等. 2017. 基于红外相机技术的野生白鹇活动规律[J]. 四川动物, 36(1): 25-29. |
夏参军, 乔建芳, 杨维康, 等. 2009. 冬季鹅喉羚昼间行为时间分配及活动节律[J]. 生态学杂志, 28(2): 283-287. |
熊志斌, 余登利, 谭成江, 等. 2003. 茂兰自然保护区白鹇种群数量与栖息地保护[J]. 贵州大学学报(自然科学版), 20(2): 200-204. |
徐建国, 陈斌, 赖雅芬, 等. 2019. 戴云山山系野生白鹇活动节律研究[J]. 福建林业, (2): 45-48. |
晏鸣霄, 孙楠, 顾伯健, 等. 2021. 同域分布的绿孔雀与白鹇时空生态位分化[J]. 四川动物, 40(2): 150-158. |
游卫云, 高欣, 梅爱君, 等. 2019. 基于红外触发相机技术的白鹇栖息地选择研究[J]. 四川动物, 38(5): 564-570. |
余建平, 钱海源, 陈小南, 等. 2017. 基于红外相机技术的白鹇日活动节律研究[J]. 动物学杂志, 52(6): 937-944. |
原宝东, 闫永峰, 程志营, 等. 2017. 黑颈长尾雉春夏夜栖地特征与差异性分析[J]. 林业科学, 53(9): 143-150. |
曾宾宾, 邵明勤, 赖宏清, 等. 2013. 性别和温度对中华秋沙鸭越冬行为的影响[J]. 生态学报, 33(12): 3712-3721. |
张晓辉, 徐基良, 张正旺, 等. 2004. 河南陕西两地白冠长尾雉的集群行为[J]. 动物学研究, 25(2): 89-95. |
赵联军, 刘鸣章, 罗春平, 等. 2020. 四川王朗国家级自然保护区血雉的日活动节律[J]. 四川动物, 39(2): 121-128. |
赵玉泽, 曹婉露, 余进, 等. 2016. 基于红外相机技术的白冠长尾雉集群行为研究[J]. 生态学报, 36(9): 2513-2520. |
赵玉泽, 王志臣, 徐基良, 等. 2013. 利用红外照相技术分析野生白冠长尾雉活动节律及时间分配[J]. 生态学报, 33(19): 6021-6027. |
周晓禹, 王晓明, 姜振华. 2008. 贺兰山石鸡越冬期昼间行为时间分配及活动规律[J]. 东北林业大学学报, 36(5): 44-46. |
朱淑怡, 段菲, 李晟. 2017. 基于红外相机网络促进我国鸟类多样性监测: 现状、问题与前景[J]. 生物多样性, 25(10): 1114-1122. |
Everts LG, Strijkstra AM, Hut RA, et al. 2004. Seasonal variation in daily activity patterns of free-ranging European ground squirrels (Spermophilus citellus)[J]. Chronobiology International, 21(1): 57-71. |
Flannigan G, Stookey JM. 2002. Day-time time budgets of pregnant mares housed in tie stalls: a comparison of draft versus light mares[J]. Applied Animal Behaviour Science, 78(2): 125-143. |
Karanth UK, Nichols JD. 1998. Estimation of tiger densities in India using photographic captures and recaptures[J]. Ecology, 79(8): 2852-2862. |
Masatoshi Y. 2004. Monitoring diversity and abundance of mammals with camera traps: a case study on Mount Tsukuba, central Japan[J]. Masatoshi Yasuda, 29(1): 37-46. |
pépin D, Renaud P, Dumont B, et al. 2006. Time budget and 24-h temporal rest-activity patterns of captive red deer hinds[J]. Applied Animal Behaviour Science, 101(3-4): 339-354. |
Silveira L, Jácomo ATA, Diniz-Filho JAF. 2003. Camera trap, line transect census and track surveys: a comparative evaluation[J]. Biological Conservation, 114(3): 351-355. |