 2021, Vol. 40
2021, Vol. 40扩展功能
文章信息
- 晏鸣霄, 孙楠, 顾伯健, 贺如川, 刘瑛
- YAN Mingxiao, SUN Nan, GU Bojian, HE Ruchuan, LIU Ying
- 同域分布的绿孔雀与白鹇时空生态位分化
- Spatio-Temporal Niche Differentiation of Sympatric Green Peafowl (Pavo muticus) and Silver Pheasant (Lophura nycthemera)
- 四川动物, 2021, 40(2): 150-159
- Sichuan Journal of Zoology, 2021, 40(2): 150-159
- 10.11984/j.issn.1000-7083.20200418
-
文章历史
- 收稿日期: 2020-11-02
- 接受日期: 2021-01-04






2. 中国科学院大学, 北京 100049;
3. 郑州大学生命科学学院生物多样性研究所, 郑州 450001;
4. 复旦大学生命科学学院, 上海 200433
2. University of Chinese Academy of Science, Beijing 100049, China;
3. School of Life Science, Zhengzhou University, Zhengzhou 450001, China;
4. School of Life Sciences, Fudan University, Shanghai 200433, China
生态位分化对同域分布的物种共存的重要性是被普遍认可的(储诚进等,2017)。根据竞争排斥原理,生态需求相似的竞争者同域分布时,必须通过至少一个生态位维度的分化才能实现稳定共存(Hardin,1960)。生态位分化主要在生境、食物和时间3个维度上发生(Schoener,1974),其中,海拔(垂直空间)和植被类型(水平空间)属于生境维度,日活动节律和季节变化属于时间维度。对比同域分布近缘物种的生态位特征,有利于指导多物种保护工作的实践(Linnell & Strand,2010)。
云南是中国唯一有绿孔雀Pavo muticus分布的省份,且滇中地区是其重点分布区域(滑荣等,2018;Kong et al., 2018)。经过前期调查,在玉溪市峨山和新平交界处发现有绿孔雀与多种雉类共存现象。其中,绿孔雀和白鹇Lophura nycthemera为当地优势雉类物种(表 1)。绿孔雀和白鹇同属鸡形目Galliformes雉科Phasianidae,均为典型的热带-亚热带林栖物种(郑光美,2015),世界分布范围有重叠(图 1)。相近的体型和食性需求(Madge et al., 2000;郑光美,2015)使二者在同域分布时可能成为潜在的竞争者,是考察大型雉类生态位分化的理想对象。绿孔雀是国家Ⅰ级重点保护野生动物,被世界自然保护联盟(IUCN)濒危物种红色名录列为濒危(EN)等级(IUCN,2020)、《中国脊椎动物红色名录》(蒋志刚等,2016)列为极危(CR)等级,国内现存数量仅235~280只(滑荣等,2018);白鹇是国家Ⅱ级重点保护野生动物。国内对白鹇的研究历史较长,对其活动节律和生境利用等多有研究(高育仁,余德群,1995;余建平等,2017;刘佳等,2019)。关于白鹇及其同域分布的雉类种间生态位分化的研究也较多,陈俊豪等(2009)研究了白鹇与白颈长尾雉Syrmaticus elliooti秋冬季空间生态位差异,刘鹏等(2012)对比了江西官山自然保护区4种雉类的生境选择差异,王方等(2020)比较了云南哀牢山白鹇和原鸡Gallus gallus的活动节律等。而绿孔雀生性机警敏感不易观察,有关野生绿孔雀的生态学研究较少,且限于单一物种,集中在其生境利用和活动节律方面(杨晓君等,2000;刘钊等, 2008, 2018;Liu et al., 2009;李旭等,2016)。目前尚无对绿孔雀和与其同域分布的雉类物种之间生态位分化的研究。
| 物种 | 保护等级 | 拍摄到的相机位点数 | 独立事件数(旱季/雨季) | 相对多度指数RAI | |
| IUCN | RLCV | ||||
| 绿孔雀 Pavo muticus |
EN | CR | 25 | 73/40 | 0.73 |
| 白鹇 Lophura nycthemera |
LC | LC | 14 | 67/22 | 0.56 |
| 原鸡 Gallus gallus |
LC | NT | 8 | 11/7 | 0.12 |
| 黑颈长尾雉 Syrmaticus humiae |
NT | VU | 7 | 3/11 | 0.09 |
| 白腹锦鸡 Chrysolophus amherstiae |
LC | NT | 1 | 3/0 | 0.02 |
| 中华鹧鸪 Francolinus pintadeanus |
LC | NT | 0 | 0 | 0 |
| 总计 | 32* | 237 | |||
| 注:IUCN. 世界自然保护联盟濒危物种红色名录(IUCN, 2020),RLCV. 中国脊椎动物红色名录(蒋志刚等,2016);保护等级: LC. 无危,NT. 近危,VU. 易危,EN. 濒危,CR. 极危;*不计重复的相机位点 Notes: IUCN. The IUCN Red List of Threatened Species (IUCN, 2020), RLCV. Red List of China's Vertebrates (Jiang et al., 2016); protection category: LC. Least Concern, NT. Near Threatened, VU. Vulnerable, EN. Endangered, CR. Critically Endangered;* exclude the repetitive camera stations | |||||

|
| 图 1 绿孔雀和白鹇的世界分布(左)、研究区域红外相机位点(右) Fig. 1 Worldwide distribution of Pavo muticus and Lophura nycthemera (left), and the distribution of the camera-trap stations in the research area (right) 2个物种分布范围引自BirdLife International(2016, 2018) Distribution maps of the 2 species were downloaded from BirdLife International(2016, 2018) |
| |
在没有绿孔雀分布的研究区域中,白鹇通常是森林雉类的优势物种,占据更宽的生态位(陈俊豪等,2009;刘鹏等,2012)。然而,当这2种大型雉类同域分布时,二者的生态位会如何发生变化以实现稳定共存,这是本研究试图解答的问题。本研究在玉溪市峨山和新平交界处山地利用红外相机技术对绿孔雀和白鹇进行调查,比较二者在海拔、植被类型利用和日活动节律3个生态位维度上的异同,为深入探讨近缘雉类物种的共存机制提供理论依据,并提出合理的保护建议。
1 研究方法 1.1 研究区域概况研究区域位于云南省玉溪市峨山县和新平县交汇处(102°3′50″~102°9′41″E,24°3′27″~24°10′10″N;图 1),海拔1 150~1 650 m。植被类型为以云南松Pinus yunnanensis为主构成的暖性针叶林和半湿润常绿阔叶林。滇中地区属低纬高原,南亚热带、中亚热带半湿润季风气候,一年分为旱季(11月至次年4月)和雨季(5—10月)。旱季温度低且降水少,平均日出时间为07∶ 30,日落时间为19∶ 00;雨季温度高且湿度大,平均日出时间为06∶ 45,日落时间为19∶ 35。
1.2 数据收集和整理红外相机技术被广泛用于野生动物调查和研究,其隐蔽性强,对动物干扰小,且能够在大范围长时间内有效记录连续的时空数据(李晟等,2014;肖治术等,2014),是研究野生动物自然活动情况的有利工具。本研究于2018年6月—2019年5月基于红外相机技术在研究区域内开展雉类物种监测。在区域内制作1 km×1 km网格地图,每个网格中设置1台红外相机(Ltl-6511MC;猎科公司,珠海),任意2台相机直线距离大于500 m。布设过程中未使用诱饵,确保记录动物在自然状态下的活动。到达合适地点后,沿小路或有野生动物活动痕迹的地方布设红外相机,在面向小路或活动痕迹3~5 m处选择视野开阔的位置,将相机固定在离地面0.5~1.2 m的树干上,镜头避开阳光直射的角度。红外相机参数设置为“拍照”,连续拍摄3张,触发间隔10 s,24 h工作。相机固定后,清除相机前的树枝和杂草,避免错误触发。记录相机位点信息。每6个月回收一次数据,并更换电池和存储卡。
红外相机照片回收后,使用生物多样性检测仪管理系统(Bio-Photo V2.1;易安卫士高端技术装备研发中心)提取照片中日期和时间等基础信息,参考《中国野外鸟类手册》(约翰·马敬能等,2000)对照片中的野生动物进行物种鉴定,并由系统直接导出Excel,对数据进行后续整理。为避免过高估计物种探测率,将同一物种在同一相机位点连续30 min(±1 min)内的拍摄记录记为一次独立事件(O'Brien,2008)。
1.3 数据分析 1.3.1 相对多度指数(relative active index,RAI)相对多度指数适合用来衡量基于红外相机获得的调查区域内物种的相对多度。采用O'Brien等(2008)提出的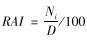
将海拔从1 650 m开始向下每100 m划为一个水平(1 350 m以下多为沟谷,面积较小,划为一个水平),共划分为4个水平。为对比绿孔雀和白鹇在各海拔水平的利用情况,使用Wilcoxon秩和检验对同季节多物种之间以及单物种多季节之间进行差异性检验,显著性水平设置为α=0.05。植被类型按当地主要林型分为针叶林、阔叶林、针阔混交林和灌丛4种。通过比较绿孔雀和白鹇在各植被类型中被探测到的独立事件数的百分比来考察不同季节不同物种对植被的利用情况。
1.3.3 日活动节律采用非参数核密度估计方法分析日活动节律。使用R Studio(R Core Team,2019)中的overlap(Meredith & Ridout,2014)绘制物种核密度曲线图,并计算重叠指数:重叠指数根据成对物种较小的样本数选取,来判断重叠程度:当样本量小于50时采用Delta1值,当样本量大于75时采用Delta4值。activity包(Rowcliffe,2016)利用Wald test对同一分布的循环预测值进行概率检验考察同一物种季节间以及不同季节物种间日活动节律的差异性。统计分析和制图均在R Studio中处理。
2 研究结果 2.1 雉类多样性情况在研究区域内布设红外相机位点77个,去除被偷和损坏的位点,最终获得63个有效相机位点,累计使用相机191台次。雨季降雨量大,许多相机后期工作中断,为确保调查强度一致,仅取5—9月的数据代表雨季;旱季相机均正常工作,有效数据时间为11—4月。调查期间在32个位点拍摄到绿孔雀、白鹇、黑颈长尾雉Syrmaticus humiae、白腹锦鸡Chrysolophus amherstiae和原鸡5种雉类(中华鹧鸪Francolinus pintadeanus在走样线时调查到,但未被红外相机探测到),共计237个独立事件,其中,绿孔雀的相对多度指数最高(RAI=0.73),其次为白鹇(RAI=0.56),二者为当地雉类的优势物种(表 1)。拍到绿孔雀和白鹇的相机位点数分别为25个和14个,二者都拍到的位点共有9个(表 2)。旱季探测到二者的独立事件数比雨季的更多。
| 生境 | 类型/水平 | 有效相机位点数 | 拍到的位点数/独立事件数 | |
| 绿孔雀 Pavo muticus |
白鹇 Lophura nycthemera |
|||
| 植被类型 | 阔叶林 | 24 | 8/16 | 9/79 |
| 针叶林 | 12 | 10/52 | 3/8 | |
| 针阔混交林 | 15 | 2/25 | 1/1 | |
| 灌丛 | 12 | 5/20 | 1/1 | |
| 海拔/m | < 1 350 | 13 | 4/8 | 2/7 |
| 1 351~1 450 | 22 | 8/19 | 7/72 | |
| 1 451~1 550 | 20 | 9/61 | 4/9 | |
| 1 551~1 650 | 8 | 3/25 | 1/1 | |
绿孔雀和白鹇对区域内4种植被类型均有利用,但是利用率显著不同。绿孔雀对各种植被类型的利用较平均,其中,对针叶林的利用率全年最高(旱季:34.25%,雨季:55.00%),旱季利用率次高的是针阔混交林(28.77%),雨季则更多出现在灌丛(32.50%)。白鹇对阔叶林的利用表现出明显偏好(旱季:93.00%,雨季:77.27%),对针叶林也有一定的利用(旱季:6.00%,雨季:18.18%),但在其他2个植被类型中各只有1个独立事件发生(图 2)。

|
| 图 2 绿孔雀和白鹇在不同季节对不同植被类型的利用情况 Fig. 2 Utilization of different vegetation types of Pavo muticus and Lophura nycthemera in different seasons |
| |
绿孔雀分布的海拔范围比白鹇更宽,其主要活动海拔明显更高(1 486 m±75.2 m vs. 1 390 m±71.9 m; w=8 122.5,P < 0.001)。绿孔雀旱季活动的海拔范围比雨季更宽,但是差异不明显(1 480 m±78.1 m vs. 1 498 m±69.1 m; w=1 245,P=0.19);白鹇旱季活动的海拔则比雨季明显更低(1 382 m±62.8 m vs. 1 414 m±92.1 m; w=356.5,P < 0.001)(图 3)。

|
| 图 3 绿孔雀和白鹇在不同季节中海拔分布情况 Fig. 3 Elevation distributions of Pavo muticus and Lophura nycthemera in different seasons |
| |
绿孔雀和白鹇均为昼行性物种。绿孔雀为晨昏活动模式,日间活动的2个高峰分别出现在日出后和日落前,活动低谷期在正午,季节间日活动节律之间的差异无统计学意义(Dhat1=0.75,P=0.15;图 4:a)。白鹇只有1个日间活动高峰,主要出现在午后,旱季的活动高峰(约17∶ 00)比雨季(约13∶ 00)更晚,季节间活动节律差异有统计学意义(Dhat1=0.62,P < 0.01;图 4:b)。绿孔雀和白鹇日活动节律在旱季(Dhat1=0.55,P < 0.001;图 4:c)和雨季(Dhat1=0.61,P < 0.01;图 4:d)之间的差异有统计学意义。

|
| 图 4 绿孔雀和白鹇的日活动节律:单物种季节间对比(a和b)和单季节物种间对比(c和d) Fig. 4 Daily activity of Pavo muticus and Lophura nycthemera: comparison of single species between seasons (a and b), and seasonal comparison between species (c and d) |
| |
在本研究区域中,绿孔雀和白鹇虽然都是优势雉类物种,但前者的相对多度却是其他所有同域分布的雉类相对多度之和,其空间生态位,无论在海拔分布还是植被类型利用上都比白鹇广泛得多。
在没有绿孔雀分布的研究区域中,白鹇通常是森林雉类的优势物种,海拔分布范围相对较宽(陈俊豪等,2009;刘鹏等,2012),日活动节律也是正常的晨昏双峰特征(高育仁,余德群,1995;刘鹏等,2012;余建平等,2017;刘佳等,2019)。然而,白鹇在本研究中却明显不同于其他研究区域,活动只集中在海拔1 350~1 400 m这一相对狭窄的区间内,日活动节律也仅有单个峰值,且与绿孔雀活动高峰明显错开。Schoener(1974)认为同域物种生态位分化的强度随空间、食物和时间的顺序递减。由于时间本身具有刚性不容易改变,日活动节律调整通常被认为是最后被启动的一种共存机制,特别是在活动空间受限制或干扰的情况下(Harrington et al., 2009)。因此,近缘物种同域分布时首先会在空间分布上发生显著变化,在有限空间内的种间竞争会导致日活动节律的改变(孙楠等,2020;Sun et al., 2020)。本研究区域中,白鹇和绿孔雀在时空生态位上发生的改变和分化也印证了上述规律。
在东南亚地区进行的研究显示,绿孔雀偏好利用较为开阔的“落叶龙脑香林”和半常绿季雨林(Brickle,2002;Saridnirun et al., 2017)。但是,本研究发现绿孔雀对栖息地的选择在旱季没有明显偏好,在雨季则主要利用针叶林。郑光美(2015)认为绿孔雀选择针叶林是被迫适应,本研究则认为,滇中地区针叶林干燥和林下较开阔的特征可能使其作为东南亚同类植被的替代林型。另一方面,求偶期的雄性绿孔雀需要开阔的林地开屏展示(Brickle,2000),针叶林可以满足此需求。实地调查时目击了绿孔雀在针叶林下刨食白蚁巢穴,说明针叶林中也有绿孔雀的食物分布;同样,当地灌丛以华西小石积Osteomeles schwerinae为优势物种,访问当地居民得知,这也是绿孔雀食物之一。另外,绿孔雀的体色使其在开阔林地也较易隐蔽。相反,白鹇却几乎不在针叶林和灌丛中出现,而是对盖度较高、湿度较大的阔叶林表现出明显偏好,这可能与其雄性体色与环境差异过大,需要隐蔽有关(卢汰春,1991)(图 5)。绿孔雀在旱季也会增加对阔叶林的利用,可能因为绿孔雀的雏鸟孵化期在5、6月(郑光美,2015),相对湿润茂密的阔叶林有利于雏鸟隐蔽,且有落果供其觅食(Brickle,2002)。白鹇在雨季则会增加对针叶林的利用,可能与该季节针叶林中可提供的食物增多有关。

|
| 图 5 在各自生境的绿孔雀(左)和白鹇(右) Fig. 5 Photos of Pavo muticus (left) and Lophura nycthemera (right) in their habitats |
| |
绿孔雀是中国最为珍稀的保护物种之一,盗猎和栖息地丧失是其受威胁的主要原因(IUCN,2020)。本研究区域中有6种雉类,其中5种在中国脊椎动物红色名录中被列为近危(NT)以上等级,相对多度指数最高的是绿孔雀。虽然本次调查尚不足以对种群数量进行估计,但根据红外相机拍摄到雉类的位点数(32/63)可以推算该地区约50%区域有野生雉类栖息,40%(25/63)的区域有绿孔雀活动。这说明当地政府的严格管理、常规巡护和对植被维护等工作使绿孔雀免受前述威胁,保护成效显著。但是,当地人为活动的影响仍不容忽视。有研究表明,绿孔雀主要选择河谷两侧的低地栖息(Brickle,2002;Liu et al., 2009;Saridnirun et al., 2017)。本研究中绿孔雀主要分布在1 500 m左右的山地,实地调查则发现当地海拔1 200 m左右的亚尼河河谷地带人为活动强度较高,这可能是迫使绿孔雀向较高海拔迁移的主要原因。此外,研究区域的西北部有一片雉类分布的空白地区为育苗基地,人为活动较多,且植被类型以单一针叶林为主,缺少阔叶林成份,可能无法在旱季为野生雉类提供水源和食物。因此,控制人为干扰、增加该区域半湿润常绿阔叶林的面积对提高栖息地质量以及维持当地大中型雉类种群有重要意义。绿孔雀在该地区的广泛适应性在其他地区并不多见,建议将该地区划为以绿孔雀为主的重点雉类保护区域,未来将开展更多关于野生绿孔雀的基础生态学研究,为制定更详尽的保护策略提供数据支持。
陈俊豪, 黄晓凤, 鲁长虎, 等. 2009. 白颈长尾雉与白鹇秋冬季空间生态位比较[J]. 生态学杂志, 28(12): 2546-2552. |
储诚进, 王酉石, 刘宇, 等. 2017. 物种共存理论研究进展[J]. 生物多样性, 25(4): 345-354. |
高育仁, 余德群. 1995. 白鹇海南亚种的生态和现状[J]. 动物学研究, 16(4): 353-358. |
滑荣, 崔多英, 刘佳, 等. 2018. 中国绿孔雀种群现状调查[J]. 野生动物学报, 39(3): 681-684. |
蒋志刚, 江建平, 王跃招, 等. 2016. 中国脊椎动物红色名录[J]. 生物多样性, 24(5): 500-551. |
李晟, 王大军, 肖治术, 等. 2014. 红外相机技术在我国野生动物研究与保护中的应用与前景[J]. 生物多样性, 22(6): 685-695. |
李旭, 刘钊, 周伟, 等. 2016. 云南楚雄恐龙河保护区绿孔雀春季栖息地选择和空间分布[J]. 南京林业大学学报(自然科学版), 3: 87-93. |
刘佳, 李生强, 汪国海, 等. 2019. 喀斯特生境中白鹇的活动节律时间分配及集群行为[J]. 广西师范大学学报(自然科学版), 37(3): 156-165. |
刘鹏, 黄晓凤, 顾署生, 等. 2012. 江西官山自然保护区四种雉类的生境选择差异[J]. 动物学研究, 33(2): 170-176. |
刘钊, 周伟, 蒋志刚, 等. 2018. 云南元江上游石羊江河谷绿孔雀夜宿地选择[J]. 野生动物学报, 39(3): 594-599. |
刘钊, 周伟, 张仁功, 等. 2008. 云南元江上游石羊江河谷绿孔雀不同季节觅食地选择[J]. 生物多样性, 16(6): 539-546. |
卢汰春. 1991. 中国珍稀濒危野生鸡类[M]. 福州: 福建科学技术出版社.
|
孙楠, 刘晨阳, Kongkasawat J, 等. 2020. 人造园林中斑文鸟和白腰文鸟的巢址选择[J]. 四川动物, 39(3): 316-322. |
王方, 叶思达, 汤永晶, 等. 2020. 云南哀牢山南段原鸡与白鹇活动节律及种间关系[J]. 野生动物学报, 41(3): 641-647. |
肖治术, 李欣海, 姜广顺. 2014. 红外相机技术在我国野生动物监测研究中的应用[J]. 生物多样性, 22(6): 683-684. |
杨晓君, 文贤继, 杨岚, 等. 2000. 春季绿孔雀的栖息地及行为活动的初步观察[C]//第四届海峡两岸鸟类学术研讨会论文集. 昆明: 海峡两岸鸟类学术研讨会.
|
余建平, 钱海源, 陈小南, 等. 2017. 基于红外相机技术的白鹇日活动节律研究[J]. 动物学杂志, 52(6): 937-944. |
约翰·马敬能, 卡伦·菲利普斯, 何芬奇. 2000. 中国鸟类野外手册[M]. 长沙: 湖南教育出版社.
|
郑光美. 2015. 中国雉类[M]. 北京: 高等教育出版社.
|
BirdLife International. 2016. Lophura nycthemera[DB/OL]. [2020-08-30]. https://dx.doi.org/10.2305/IUCN.UK.2016-3.RLTS.T22679220A92808107.
|
BirdLife International. 2018. Pavo muticus[DB/OL]. [2020-08-30]. http://dx.doi.org/10.2305/IUCN.UK.2018-2.
|
Brickle NW. 2002. Habitat use, predicted distribution and conservation of green peafowl (Pavo muticus) in Dak Lak Province, Vietnam[J]. Biological Conservation, 105(2): 189-197. DOI:10.1016/S0006-3207(01)00182-3 |
Hardin G. 1960. Competitive exclusion principle[J]. Science, 131(3409): 1292-1297. DOI:10.1126/science.131.3409.1292 |
Harrington LA, Harrington AL, Yamaguchi N, et al. 2009. The impact of native competitors on an alien invasive: temporal niche shifts to avoid interspecific aggression?[J]. Ecology, 90(5): 1207-1216. DOI:10.1890/08-0302.1 |
IUCN. 2020. The IUCN red list of threatened species. Version 2020-2[DB/OL]. [2020-08-30]. https://www.iucnredlist.org.
|
Kong DJ, Wu F, Shan P, et al. 2018. Status and distribution changes of the endangered green peafowl (Pavo muticus) in China over the past three decades (1990s-2017)[J/OL]. Avian Research, 9: 18[2020-07-01]. https://doi.org/10.1186/s40657-018-0110-0.
|
Linnell JDC, Strand O. 2010. Interference interactions, co-existence and conservation of mammalian carnivores[J]. Diversity & Distributions, 6(4): 169-176. |
Liu YQ, Han LX, Xie YC, et al. 2009. The status and habitat use of green peafowl Pavo muticus in Shuangbai Konglonghe Nature Reserve, China[J]. International Journal of Galliformes Conservation, 1: 32-35. |
Madge S, McGowan PJK, Kirwan GM. 2002. Pheasants, partridges and grouse: a guide to the pheasants, partridges, quails, grouse, guineafowl, buttonquails and sandgrouse of the world[M]. UK: A&C Black.
|
Meredith M, Ridout M. 2014. Overlap: estimates of coefficient of overlapping for animal activity patterns[EB/OL]. (2020-05-09)[2020-06-01]. https://CRAN.R-project.org/package=overlap.
|
O'Brien TG. 2008. On the use of automated cameras to estimate species richness for large- and medium-sized rainforest mammals[J]. Animal Conservation, 11(3): 179-181. DOI:10.1111/j.1469-1795.2008.00178.x |
R Core Team. 2019. R: a language and environment for statistical computing[EB/OL]. (2019-04-26)[2020-06-01]. Vienna, Austria: R Foundation for Statistical Computing. https://www.R-project.org/.
|
Rowcliffe M. 2016. 'Package "activity": animal activity statistics', R package version 1.1[EB/OL]. (2016-01-16)[2020-06-01]. https://CRAN.R-project.org/package=activity.
|
Saridnirun G, Meckvichai W, Dumrongrojwatthana P. 2017. Seasonal distribution and habitat use of the green peafowl Pavo muticus, Linnaeus 1766, in Wiang Lor Wildlife Sanctuary, Phayao Province, Northern Thailand[J]. Chiang Mai Journal of Science, 44(3): 824-838. |
Schoener TW. 1974. Resource partitioning in ecological communities[J]. Science, 185(4145): 27-39. DOI:10.1126/science.185.4145.27 |
Sun N, Cao GH, Li GG, et al. 2020. Macaca leonina has a wider niche breadth than sympatric M. mulatta in a fragmented tropical forest in southwest China[J/OL]. American Journal of Primatology, 82(2): e23100[2020-09-01]. https://doi.org/10.1002/ajp.23100.
|