 2020, Vol. 39
2020, Vol. 39扩展功能
文章信息
- 李孪鑫, 潘鸿茹, 田晔, 雷泽锋, 邹红菲
- LI Luanxin, PAN Hongru, TIAN Ye, LEI Zefeng, ZOU Hongfei
- 吉林莫莫格国家级自然保护区白鹤与豆雁迁徙期资源利用特征与生态位分化
- Utilization Characteristics of Resources and Niche Differentiation of Grus leucogeranus and Anser fabalis During Migration Period in the Momoge National Nature Reserve, Jilin
- 四川动物, 2020, 39(4): 417-423
- Sichuan Journal of Zoology, 2020, 39(4): 417-423
- 10.11984/j.issn.1000-7083.20190412
-
文章历史
- 收稿日期: 2019-12-02
- 接受日期: 2020-04-15






生态位理论是生态学最重要的基础理论之一,对理解群落结构、阐明生物群落内的种间竞争及共存机制起着重要作用(李等,2003;马英等,2011;郭宏等,2016)。根据生态位理论,同域分布的物种通过生态位的分化,可有效减轻或避免有限资源条件下的种间竞争(Pianka,1981)。由于生境、资源特征的不同和鸟类适应环境的多样性,生态位分化表现出多种形式(杨小农等,2012)。湿地是众多水鸟越冬、繁殖、迁徙的良好场所和主要聚居地,湿地类型的多样性、栖息地环境条件的异质性和食物资源分布的差异性,深刻地影响着鸟类的群落组成,以及由此引起的鸟类资源生态位互补与重叠特征,体现了鸟类栖息地选择和迁徙习性形成的多样化规律(张晶等,2012)。
白鹤Grus leucogeranus和豆雁Anser fabalis为大型迁徙水鸟,吉林莫莫格国家级自然保护区不仅是众多水鸟迁徙途中的重要停歇地,也是濒危物种白鹤主要的迁徙停歇地。实地调查发现,在春季和秋季的迁徙期保护区的一些湿地中存在白鹤种群与大量豆雁种群混群现象,两者利用了相似的栖息环境,且两者的主要取食地有一定重叠,可能有潜在的竞争关系。这类物种会在生境选择等方面形成生态位分化以减轻种间竞争并实现共存(Zhou et al., 2014)。本研究以保护区内的白鹤与豆雁为主要研究对象,试图探讨迁徙期白鹤与豆雁之间的生态位分化和资源分割机制,对白鹤与豆雁的种间关系进行更深层次地认知,以期为濒危鹤类与其他水鸟的种间关系以及生态学研究提供一定理论依据;为进一步对保护区内水鸟群落的共存研究提供一定的参考价值。
1 研究区和研究物种 1.1 研究区概况吉林莫莫格国家级自然保护区地处吉林省白城市镇赉县,地理坐标为123°27′~124°4′E,45°42′~46°18′N,总面积14.40×104 hm2(左悦,2008),其中自然湿地面积占总面积的80%以上,是松嫩平原保存最完整的湿地之一。保护区平均海拔142 m,属大陆性季风气候,年均气温4.2 ℃,年均降水量391.8 mm,降水主要集中在6—8月,且呈现出光、热同季的特征(杨成玉,2008)。保护区地理位置优越、植被条件良好以及觅食条件丰富的特点,吸引了大量水禽等珍稀野生动物来此繁殖或栖息。保护区主要保护对象是珍稀禽类与其他濒危物种及其生存环境。区内有丰富的野生动物资源:鱼类4目11科52种,两栖、爬行类3目6科12种,鸟类17目55科298种,兽类4目9科25种(范巍巍,2012)。
1.2 白鹤与豆雁的生物学信息白鹤属大型涉禽,体长130~140 cm,体质量7~10 kg,嘴峰长约16.2 cm,跗蹠长24.1~26.2 cm,喙较长且喙端较尖。白鹤是对栖息地要求较为特化的一个物种,对有浅水的湿地环境有较强的需求(范巍巍,2012),常栖息于开阔平原沼泽草地、苔原沼泽和大的湖泊边缘及浅水沼泽地带,主要以植物的根、地下茎、芽、种子以及一些软体动物为食。豆雁属大型雁类,体长69~ 80 cm,体质量2.6~3.4 kg,嘴峰长6.6~7.8 cm,跗蹠长6.3~8.4 cm,有扁平的喙,边缘锯齿状,主要栖息于开阔平原草地、沼泽、水库、江河湖泊及农田地区,主要食物为植物嫩芽、叶,芦苇Phragmites communis、苔草Carex spp.等和一些农作物的种子(赵正阶,2001)。苔草滩是豆雁在安徽升金湖国家级自然保护区的主要栖息地,苔草也是豆雁食物的重要组成部分(刘静,2011)。实地调查中发现,白鹤和豆雁有在相同觅食地进行觅食的行为发生,表明二者食性确有相似之处,可能存在一定的竞争关系。
2 研究方法 2.1 调查方法2018年秋季(9—10月)和2019年春季(3—4月)在吉林莫莫格国家级自然保护区内共进行6次野外调查,釆用固定路线寻查的样线法和定点观察法相结合的方法,在少力泡子、报马吉岗子、蒙古索口、孟天昭、格力吐泡等18个观测点进行了调查(图 1),调查过程中使用8×42双筒望远镜和20~60×80单筒望远镜对各个观察点的白鹤与豆雁进行观察识别,对白鹤和小群豆雁,采用直接计数法计数;对大群豆雁,采用集团计数法记录,并记录相应的生境类型、觅食地类型以及所在环境因子。对于栖息在湿地中的白鹤和豆雁,选择手持激光测距仪测量其所在位置的离岸距离,作为栖息地空间的水平分布。

|
| 图 1 吉林莫莫格国家级自然保护区的调查区域 Fig. 1 Survey area in the Momoge NationalNature Reserve, Jilin |
| |
根据实地调查并综合考虑湿地利用类型及水鸟栖息地利用状况后(赵凤婷,2013;李全亮,2017),将生境划分为农田、草甸、芦苇沼泽、苔草沼泽和明水面5种类型。再根据观察到的水鸟的空间分布,将栖息地空间划分为3个方面,分别为: (1)栖息环境,即有水鸟分布的环境,根据生境类型划分为农田、草甸、芦苇沼泽、苔草沼泽和明水面5种类型;(2)离岸距离,划分为0~50 m、50~100 m、100~150 m、150~200 m、>200 m共5个等级;(3)觅食地类型,即有水鸟食物存在且水鸟有觅食行为发生的环境,划分为草地(以羊草Leymus chinensis等草本植物为主)、泥地(没有明显植物或有少量苦草Vallisneria natans等沉水草本植物)、农田(以玉米Zea mays等农作物为主)、苔草滩(以苔草等莎草科Cyperaceae植物为主)、芦苇滩(以芦苇等禾本科Gramineae植物为主)5种类型。分别记录白鹤和豆雁在每种栖息地空间出现的次数,计算生态位宽度和重叠。
2.3 数据处理根据白鹤和豆雁在各个生境类型中的数量分布数据,计算它们对各种生境资源的利用率P(Levins,1970):Pij=Nij/N。
其中,Nij为第j种资源类型中记录到的第i种鸟类数量;N为所有资源类型中记录到的第i种鸟类数量。
计算白鹤与豆雁在各个迁徙期每个栖息地空间的生态位宽度与重叠并进行比较,并对各个栖息地空间的差异采用Mann-Whitney U检验。各个栖息地空间的比较分析借助Excel和SPSS完成。
生态位宽度采用Shannon-Wiener多样性指数为基础的生态位宽度指数(Colwell & Futuyma,1971):Bi=-Σ1RPijlnPij;
生态位重叠采用Schoener(1974)提出的公式: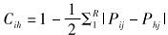
其中,Pij和Phj分别是第i种鸟类和第h种鸟类在第j种资源中出现的数目占各自个体总数的比例,R为生态位资源等级数值。Bi的值为0~1,0表示没有利用,1表示对所有等级都有同样的利用。Cih的值为0~1,0表示完全不重叠,1表示完全重叠(Colwell & Futuyma,1971)。
3 结果与分析在春迁期和秋迁期,白鹤对农田、草甸、芦苇沼泽、苔草沼泽和明水面5种生境均有利用,且利用形式基本相同,利用最多的是芦苇沼泽,对明水面的利用率最低。豆雁也利用了这5种生境,利用形式基本相同,利用最多的是芦苇沼泽,对苔草沼泽的利用率也相对较高,而对农田的利用率最低。春迁期白鹤的生态位宽度为0.527,豆雁为0.544,两者的生态位重叠值为0.557,差异有高度统计学意义(Z=-26.93,P < 0.01);秋迁期白鹤的生态位宽度为0.553,豆雁为0.533,两者的生态位重叠值为0.654,差异有高度统计学意义(Z=-13.57,P < 0.01)。豆雁对苔草沼泽和明水面的利用率相对较高,而白鹤偏好草甸、芦苇沼泽和农田(图 2)。

|
| 图 2 吉林莫莫格国家级自然保护区春迁期(A)和秋迁期(B)白鹤和豆雁对不同栖息环境的利用率 Fig. 2 Utilization rates of different habitats by Grus leucogeranus and Anser fabalis during spring (A) and autumn (B) migration in the Momoge National Nature Reserve, Jilin |
| |
在离岸距离方面,春迁期白鹤的生态位宽度为0.628,豆雁为0.611,两者的生态位重叠值为0.485,差异有高度统计学意义(Z=-29.43,P < 0.01);秋迁期白鹤的生态位宽度为0.645,豆雁为0.647,两者的生态位重叠值为0.612,差异有高度统计学意义(Z=-21.74,P < 0.01)。白鹤对离岸0~50 m和50~100 m的环境利用率均明显多于豆雁,而豆雁偏好离岸150~200 m以及>200 m的环境(图 3)。

|
| 图 3 吉林莫莫格国家级自然保护区春迁期(A)和秋迁期(B)白鹤和豆雁对不同离岸距离的利用率 Fig. 3 Utilization rates of different offshore distances by Grus leucogeranus and Anser fabalis during spring (A) and autumn (B) migration in the Momoge National Nature Reserve, Jilin |
| |
在觅食地类型方面,春迁期白鹤的生态位宽度为0.590,豆雁为0.486,两者的生态位重叠值为0.595,差异有高度统计学意义(Z=-35.01,P < 0.01);秋迁期白鹤的生态位宽度为0.576,豆雁为0.388,两者的生态位重叠值为0.466,差异有高度统计学意义(Z=-14.43,P < 0.01)。白鹤偏好芦苇滩、农田以及泥地作为觅食地,而豆雁偏好苔草滩(图 4)。

|
| 图 4 吉林莫莫格国家级自然保护区春迁期(A)和秋迁期(B)白鹤和豆雁对不同觅食地类型的利用率 Fig. 4 Utilization rates of different foraging habitats by Grus leucogeranus and Anser fabalisduring spring (A) and autumn (B) migration in the Momoge National Nature Reserve, Jilin |
| |
在迁徙中,随着环境的改变,水鸟会不断选择适宜的栖息地(Beerns et al., 2011)。本研究中的白鹤和豆雁在迁徙期对农田、草甸、芦苇沼泽、苔草沼泽和明水面这5种生境的利用有所不同,是生境中各种资源可获得性的不同导致资源分割的情况发生(Kober & Bairlein,2009)。而资源的可获得性是导致水鸟种群动态变化的重要原因之一(Gawlik,2002),水鸟的资源利用特征也会随资源可获得性而发生动态变化(Wermer & Hall,1979)。
白鹤与豆雁对栖息地的利用在各方面均有一定重叠,说明二者对资源有相似的需求。在栖息环境方面,白鹤与豆雁都利用了草甸、农田、芦苇沼泽、苔草沼泽和明水面,且对芦苇沼泽具有较高的利用率,可见芦苇沼泽是二者最大的竞争资源,这主要是因为芦苇沼泽能够为水鸟提供隐蔽场所和满足食物资源的需求。鸟类对资源利用策略会随着当地植被群落结构的改变而改变(Kushlan,1981),且水位高低与水鸟利用的生境和食物的可获得性密切相关(Safran et al., 2000;Zhao et al., 2010)。
白鹤与豆雁春迁期生态位重叠值比秋迁期小,是因为春迁期冰雪融化后暴露出大部分湿地,且保护区植被增多,为白鹤与豆雁供给了更多的可利用资源。当可利用的资源有限时,水鸟会选择聚集到有限的生境内来获得资源,导致竞争强度增加,从而使生态位重叠度增加(Kober & Bairlein,2009)。秋迁期的空间生态位重叠值变高,这是由于旱季可利用资源相对较少,2种水鸟可利用的生境减少,共同利用的生境增加,生态位重叠增加。
在离岸距离方面,两者的重叠度相对较低,白鹤主要选择了离岸较近的环境,而豆雁有一半甚至一半以上都选择了离岸距离>150 m的环境。主要是因为作为游禽的豆雁除了在浅滩补充足够的能量以外,比白鹤偏好水域生境,所以会离岸距离较远,而白鹤属于涉禽,主要栖息在挺水植物群落分布区,食源植物为扁秆藨草Scirpus planiculmis,适宜水深为30~50 cm(崔桢等,2018),集中在浅滩环境,所以离岸距离较近。由于两者不同的生态习性,以及所选择的生境类型也不同,使它们在不同的距离区间栖息,这样降低了在离岸距离方面的生态位重叠,从而也降低了种间竞争,有利于二者在一定区域内实现共存。这与同域分布的鸿雁Anser cygnoides和白额雁Anser albifrons的研究结果相似(郭宏等,2016)。
在觅食地类型方面,白鹤与豆雁表现出一定程度的分化。白鹤与豆雁在觅食地类型上存在显著性差异,表明它们在觅食地类型方面出现生态位的分离,导致这种差异的原因是,当2个物种共存时,二者对相同生境资源的利用方式存在差异(Oksanen,1987)。而鸟类的形态与生理特征,如鸟类体型的大小以及鸟喙的长度、宽度、形状等决定了其取食的方式(Nebel et al., 2005)。白鹤不仅利用了芦苇滩生境,对农田等其他类型的觅食地也有一定的利用;豆雁主要利用苔草滩生境,而对农田利用较少。这与两者的生理形态特征有关,白鹤的喙尖长,可以在泥地挖食或拾取农田中小颗粒农作物,而豆雁的喙扁且较短小,主要以挖掘苔草的块茎以及一些植物的地上部分为食,所以偏好短苔草草滩。所以白鹤与豆雁在觅食地类型上存在实质性的空间分离,这种空间分离可以使物种的资源竞争强度减小(González-Solís et al., 2007)。而从结果来看,无论是春迁期还是秋迁期,白鹤的生态位宽度均大于豆雁,说明相对于数量巨大的豆雁,白鹤选择了利用多种食物资源以拓宽生态位来缓解竞争。
虽然白鹤与豆雁显示了不同的生态位策略,在一定程度上体现出了生态位的分化,但同时它们又有相似的生境需求,对于芦苇沼泽和苔草沼泽这2种生境,两者都有一定的利用,并且在栖息环境和觅食地类型上有较大的生态位重叠。这也表明二者对在迁徙期湿地资源的利用存在一定的竞争关系。因此,为了更好地保护濒危物种白鹤,不可忽视白鹤与豆雁这2个物种之间的竞争关系。
崔桢, 章光新, 张蕾, 等. 2018. 基于白鹤生境需求的湿地生态水文调控研究——以莫莫格国家级自然保护区白鹤湖为例[J]. 湿地科学, 16(4): 509-516. |
范巍巍. 2012.莫莫格国家级自然保护区白鹤迁徙期栖息地觅食生境选择研究[D].哈尔滨: 东北林业大学. http://cdmd.cnki.com.cn/Article/CDMD-10225-1012445935.htm
|
郭宏, 邵明勤, 胡斌华, 等. 2016. 鄱阳湖南矶湿地国家级自然保护区2种大雁的越冬行为特征及生态位分化[J]. 生态与农村环境学报, 32(1): 90-95. |
李, 朱金兆, 朱清科, 等. 2003. 生态位理论及其测度研究进展[J]. 北京林业大学学报, (1): 100-107. |
李全亮. 2017.松嫩平原丹顶鹤与白枕鹤生境利用对比研究[D].哈尔滨: 东北林业大学. https://www.ixueshu.com/document/79da8b70f0a1bdba88c91cd506a59c29318947a18e7f9386.html
|
刘静. 2011.安徽升金湖国家级自然保护区豆雁的越冬食性和行为研究[D].合肥: 中国科学技术大学. http://cdmd.cnki.com.cn/Article/CDMD-10358-1012008991.htm
|
马英, 魏有文, 罗军, 等. 2011. 青海省海东地区小型兽空间生态位分析[J]. 动物学杂志, 46(4): 126-130. |
杨成玉. 2008.莫莫格国家级自然保护区湿地生态系统服务功能价值评估[D].长春: 东北师范大学. http://cdmd.cnki.com.cn/Article/CDMD-10200-2008107623.htm
|
杨小农, 朱磊, 郝光, 等. 2012. 瓦屋山2种山雀的生态位分化和共存[J]. 动物学杂志, 47(4): 11-18. |
张晶, 赵成章, 任悦, 等. 2018. 张掖国家湿地公园优势鸟类种群生态位研究[J]. 生态学报, 38(6): 2213-2220. |
赵凤婷. 2013.越冬白头鹤种群与三种雁的觅食生境利用与资源分割研究[D].合肥: 安徽大学. http://cdmd.cnki.com.cn/Article/CDMD-10357-1013260164.htm
|
赵正阶. 2001. 中国鸟类志上卷非雀形目[M]. 吉林: 吉林科学技术出版社.
|
左悦. 2008.莫莫格国家级自然保护区昆虫多样性初步研究[D].长春: 东北师范大学. https://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CMFD&filename=2008107841.nh
|
Beerns JM, Gawlik DE, Herring G, et al. 2011. Dynamic habitat selection by two wading bird species with divergent foraging strategies in a seasonally fluctuating wetland[J]. The Auk, 128(4): 651-622. |
Colwell RK, Futuyma DJ. 1971. On the measurement of niche breadth and overlap[J]. Ecology, 52(4): 567-576. DOI:10.2307/1934144 |
Gawlik DE. 2002. The effects of prey availability on the numerical response[J]. Ecological Monographs, 72(3): 329-346. DOI:10.1890/0012-9615(2002)072[0329:TEOPAO]2.0.CO;2 |
González-Solís J, Croxall JP, Afanasyev V. 2007. TOffshore spatial segregation in giant petrels Macronectes spp.:differences between species, sexes and seasons[J]. Aquatic Conservation Marine and Freshwater Ecosystems, 17(S1): 22-36. |
Kober K, Bairlein F. 2009. Habitat choice and niche characteristics under poor food conditions. A study on migratory nearctic shorebirds in the intertidal flats of Brazil[J]. Ardea, 97(1): 31-42.
|
Kushlan JA. 1981. Resource use strategies of wading birds[J]. Wilson Bull, 93(2): 145-163. |
Levins R. 1970. Ordering the phenomena of ecology[J]. Science, 167(3924): 1478-1480. |
Nebel S, Jackson DL, Elner RW. 2005. Functional association of bill morphology and foraging behaviour in calidrid sandpipers[J]. Animal Biology, 55(3): 235-243. DOI:10.1163/1570756054472818 |
Oksanen L. 1987. Interspecific competition and the structure of bird guilds in Boreal Europe:the importance of doing fieldwork in right season[J]. Trends Ecology & Evolution, 2(12): 376-379. |
Pianka ER. 1981. Competition and niche theory[M]//Maverick A. Theoretical ecology(second edition). Durham: Blackwell.
|
Safran RJ, Colwell MA, Isola CR, et al. 2000. Foraging site selection by non-breeding white-faced Ibos[J]. The Condor, 102: 211-215. |
Schoener TW. 1974. Some methods for calculating competition coefficients from resource-utilization spectra[J]. The American Naturalist, 108(961): 332-340. |
Wermer EE, Hall DJ. 1979. Foraging efficiency and habitat switching in competition sunfishes[J]. Ecology, 60(2): 256-264. |
Zhao MJ, Cao L, Fox AD. 2010. Distribution and diet of wintering tundra bean geese Anser fabalis serrirostris at Shengjin Lake, Yangtze River Floodplain, China[J]. Wildfowl, 60: 52-63. |
Zhou Q, Wei H, Tang H, et al. 2014. Niche separation of sympatric macaques, Macaca assamensis and M. mulatta, in limestone habitats of Nonggang, China[J]. Primates, 55(1): 125-137. |