 2020, Vol. 39
2020, Vol. 39扩展功能
文章信息
- 罗昕裕, 周昭敏
- LUO Xinyu, ZHOU Zhaomin
- 3种城市生境间蚂蚁群落组成的比较研究——以四川省南充市城区为例
- Comparison of Ant Community Composition Among Three Urban Habitats: a Case Study in Nanchong, Sichuan
- 四川动物, 2020, 39(2): 148-155
- Sichuan Journal of Zoology, 2020, 39(2): 148-155
- 10.11984/j.issn.1000-7083.20190070
-
文章历史
- 收稿日期: 2019-02-24
- 接受日期: 2019-12-13






全球城市化进程自20世纪显著加快(Grimm et al., 2008),城市扩张将自然生境转化为各种形式的人造景观,并改变当地的气候和土壤性质等环境条件(Pataki et al., 2006;Rocha-Ortega & Castaño-Meneses,2015)。这些改变可能会降低陆生动物的迁移能力,影响自然斑块中的资源流动,并导致本地生物多样性的快速丧失(Lessard & Buddle,2005;Hahs et al., 2009;Alberti,2015)。然而,城市化也为一些能够适应城市生境的物种创造了适宜的栖息地(Buczkowski & Richmond,2012),使得群落间的遗传、分类或功能相似性增加,进而促进生物同质化(Mckinney & Lockwood,2001;Olden et al., 2004;Newbold et al., 2015)。
蚂蚁(膜翅目Hymenoptera蚁科Formicidae)是指示生物多样性和环境变化的重要类群(King et al., 1998;Hoffmann,2010;李巧等,2011;刘霞等,2017)。首先,蚂蚁提供了重要的生态系统服务功能,如作为捕食者、互利者及食物网中的猎物等(Heil & McKey,2003)。其次,蚂蚁分布广泛、种类和数量众多,具有相对较短的世代间隔、利于样本采集的特点(郭萧等,2014;李巧等,2015)。此外,绝大多数蚂蚁种类迁移能力较弱,易被自然地理屏障阻碍,并且对微环境的变化特别敏感,能对其做出迅速响应(McKinney,2008;Stringer et al., 2009)。
本研究在四川省南充市城区中选择“公园与绿地”“旱地”和“乔木林地”3种土地利用类型,对蚂蚁群落组成进行调查和比较分析,不仅将为研究城市化对野生动物群落结构影响提供基础资料,还将检验蚂蚁群落作为生物指示物种反映生态系统响应的有效性。
1 材料与方法 1.1 研究地点南充市位于四川盆地东北部,嘉陵江中下游,地处我国西南部水热条件充沛的亚热带季风气候区,在大陆动物地理分区上处于东洋界到古北界的过渡地带(张玮,郑哲民,2002)。受盆地气候影响,四季分明,雨热同期,年均降水量980~1 150 mm;年均气温15.8~17.8 ℃;相对湿度76%~86%(Xu et al., 2015);年均日照时间1 200~1 500 h。现有调查发现,分布于四川省(含重庆)的蚂蚁共有78种(张玮,郑哲民,2002),在城市生境中的有26种(谭速进等,2010)。
1.2 研究方法 1.2.1 生境植被调查与蚂蚁采集方法参照国家质检总局和国家标准化管理委员批准发布并实施的《土地利用现状分类标准(GB/T 21010-2017)》中的二级类划分,在南充市西华师范大学华凤校区内选择“公园与绿地”(编码0810)、“旱地”(编码0103)和“乔木林地”(编码0301)3种土地利用类型,并分别设置8个1 m×1 m的样方。参照McIntyre等(2001)的方法,生境植被调查在以样方中心点为圆心,20 m为半径的区域内进行。生境调查指标采用草本植被覆盖度、林分郁闭度和植被的Shannon-Wiener多样性指数( H )3个指标。按照点测法,沿样方对角线,每隔10 cm垂直放置1根尖锐的样针,每根样针直径不超过2 mm,对低于2 m的植被,记录其枝叶被样针击中的数目,该数目与总样针数之比即为草本植被覆盖度(Heraldol et al., 2010)。林分郁闭度则是被树冠垂直覆盖的样针数目与总样针数之比(Magnusson et al., 1999)。为计算植被的Shannon-Wiener多样性指数,在20 m半径的样圆内分别设置5个3 m半径的子样圆(圆心间距不小于10 m),调查乔木种类及数量(宋曰钦,高长宝,2002;曹艳杰,2006;王秀云,2011;司建朋等,2019);并设置5个1 m×1 m的子样方,调查草本种类及数量(王秀云,2011)。
3种生境的草本植被覆盖度( F= 5.68,df =2,P < 0.05)、林分郁闭度( F =1.86,df =2,P < 0.05)和Shannon-Wiener多样性指数( F= 8.79,df =2,P < 0.05)的差异均有统计学意义。“乔木林地”中有多种乔木(刺槐 Robinia pseudoacacia 、栾树 Koelreuteria paniculata 、重阳木 Bischofia polycarpa 、冬青 Ilex chinensis 等)和草本植物(白茅 Imperata cylindrica 、蒲公英 Taraxacum mongolicum 、马齿苋 Portulaca oleracea 、益母草 Leonurus japonicus 、天名精 Carpesium abrotanoides 、凤尾蕨 Pteris cretica 、瓶尔小草 Ophioglossum vulgatum 等),几乎无灌木分布。“旱地”在5月的主要作物为玉蜀黍 Zea mays ,零星分布有枫杨 Pterocarya stenoptera 和樟 Cinnamomum camphora ,除农作物以外的其他草本植物分布较少。“公园与绿地”中的乔木以香樟为主,草本植被则以沿阶草 Ophiopogon bodinieri 为主,零星分布有狗牙根 Cynodon dactylon 、车轴草 Trifolium repens 、马唐 Digitaria sanguinalis 、马兰 Aster indicus 。因此,这3种土地利用类型具有不同的植被状况,分别代表了不同的生境类型。
2018年5月,通过陷阱法在上述24个样方采集蚂蚁样本。采集持续时间为48 h。不同生境样方的间距不小于1 km,同一生境的样方间距不小于10 m。陷阱设置在样方顶角处的平坦区域。陷阱由2个500 mL塑料杯套叠组成,内杯杯唇与地面齐平,以避免连续采样过程中杯子被破坏(McIntyre et al., 2001)。在塑料杯中倒入150 mL丙二醇,以杀死蚂蚁,并使其在短时间内得到有效保存。将采集的样品储存在70%的乙醇溶液中,并保存在西华师范大学标本库中。
1.2.2 蚂蚁的分类鉴定采用DNA条形码技术对采集的蚂蚁标本测序,并将 CO Ⅰ测序结果与GenBank(http://www.ncbi.nlm.nih.gov/)及BOLD Systems(http://www.boldsystems.org/)所存序列进行对比。该结果用于快速确定其所属的系统发育支系,以方便随后的形态比对。DNA提取过程中,首先去除蚂蚁腹部,然后将剩余肢体置于研钵中,用手术剪将其剪碎;加入500 μL DNA提取液和20 μL的10 mg·mL-1的蛋白酶K,于57 ℃水浴2~3 h;加入等体积的平衡酚并混匀,离心后取上清液;加入等体积的氯仿-异戊醇混合溶液,离心后取上清液;加入2倍体积的预冷无水乙醇和1/10体积的乙酸钠,保留其沉淀物;在沉淀物中加入1 mL 70%的乙醇溶液,离心后弃上清液(李淑萍,徐亚军,2010;陈媛,2014);将离心管敞口放置至乙醇完全挥发,加入40 μL ddH2O,-20 ℃保存备用(陈媛,2014)。选用通用引物LCOI490、HCO2198对 CO Ⅰ片段进行扩增(Folmer et al., 1994;陈媛,2014)。DNA提取及测序均由中国科学院昆明动物研究所生命条形码南方中心完成。
物种形态鉴定参照“ A new general catalogue of the ants of the world ”(Bolton,1995)、《中国经济昆虫志第四十七册膜翅目蚁科(一)》(唐觉等,1995)、《中国蚂蚁》(吴坚,王常禄,1995)、《广西蚂蚁》(周善义,2001)、《湖北省蚂蚁分类研究》(王维等,2009)等专著以及“Encyclopedia of Life”(http://eol.org/)、“蚂蚁网”(http://www.ants-china.com/?sp=6604)、“ANTWEB”(http://www.antweb.org/)、“蚂蚁维基”(http://www.antwiki.org/wiki/Welcome_to_AntWiki)等数据库。利用LEICA M205C体视显微镜观察蚂蚁的形态特征。
1.2.3 数据处理采用物种数( S )、Shannon-Wiener多样性指数( H )、Simpson多样性指数( D )和Pielou均匀度指数( J )分别反映各样方蚂蚁群落的物种丰富度、物种多样性、物种优势度和物种均匀度,采用单因素方差分析(One-Way ANOVA)中的最小显著性差异(least significant difference)评估3种生境间蚂蚁群落各指标的差异性(显著性水平设置为α=0.05)。H =-∑ Pi ln Pi (李玉杰等,2018);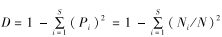
本研究共采集蚂蚁样品1 847只。检测出18种蚂蚁的 CO Ⅰ 基因(204 bp),其中6种蚂蚁与GenBank及BOLD Systems所存序列相似度达到98%~99%(津岛铺道蚁 Tetramorium tsushimae :AY64 1700;中华小家蚁 Monomorium chinense :MH754300;宽结大头蚁 Pheidole nodus :EF518375;布氏尼氏蚁 Nylanderia bourbonica :EF609962;无毛凹臭蚁 Ochetellus glaber :JQ913605;黄足尼氏蚁 Nylanderia flavipes :MH754327);12种蚂蚁与所存序列的相似度为84%~96%(法老小家蚁 Monomorium pharaonis :KJ86 1846;满斜结蚁 Plagiolepis manczshurica :AB019423;日本弓背蚁 Camponotus japonicus :KR784733;中华细颚猛蚁 Leptogenys chinensis :KC685034;山大齿猛蚁 Odontomachus monticola :FX986057;美洲切叶蚁 Myrmecina americana :KR42 1703;日本盘腹蚁 Aphaenogaster japonica :JQ742634;孟氏大头蚁 Pheidole mendanai :KJ14 2033;游举腹蚁 Crematogaster vagula :MH754499;黑褐举腹蚁 Crematogaster rogenhoferi :EU605790;汤加铺道蚁 Tetramorium tonganum :MG567163;茸毛铺道蚁 Tetramorium lanuginosum :GU709681)。经形态学比对,有5只个体为同种个体,仅能确定为蚁科Formicidae sp.,另有6只同种个体仅能确定为铺道蚁属 Tetramorium sp.;其余1 836只个体分属4亚科13属19种(表 1),其中,切叶蚁亚科Myrmicinae标本7属12种1 680只,蚁亚科Formicinae标本3属4种137只,猛蚁亚科Ponerinae标本2属2种15只,臭蚁亚科Dolichoderinae标本1属1种4只。在3种生境内均有分布的蚂蚁共有8种(津岛铺道蚁、中华小家蚁、宽结大头蚁、布氏尼氏蚁、陕西铺道蚁 Tetramorium shensiense 、法老小家蚁、满斜结蚁和日本弓背蚁),4种蚂蚁仅发现于“乔木林地”(中华细颚猛蚁、山大齿猛蚁、无毛凹臭蚁和游举腹蚁),3种蚂蚁仅发现于“公园与绿地”(台湾瘤颚蚁 Strumigenys formosa 、黑褐举腹蚁、美洲切叶蚁),汤加铺道蚁则仅发现于“旱地”。而与茸毛铺道蚁 CO Ⅰ 基因序列相似度为84%的样品为同属的陕西铺道蚁。
| 物种 | 个体数/只 | ||
| 乔木 林地 |
旱地 | 公园与 绿地 |
|
| 一 臭蚁亚科Dolichoderinae | |||
| 1.无毛凹臭蚁 Ochetellus glaber | 4 | — | — |
| 二 蚁亚科Formicinae | |||
| 2.日本弓背蚁 Camponotus japonicus | 3 | 5 | 1 |
| 3.布氏尼氏蚁 Nylanderia bourbonica | 5 | 70 | 3 |
| 4.黄足尼氏蚁 Nylanderia flavipes | 11 | — | 20 |
| 5.满斜结蚁 Plagiolepis manczshurica | 16 | 1 | 2 |
| 三 切叶蚁亚科Myrmicinae | |||
| 6.日本盘腹蚁 Aphaenogaster japonica | — | 75 | 1 |
| 7.游举腹蚁 Crematogaster vagula | 2 | — | — |
| 8.黑褐举腹蚁 Crematogaster rogenhoferi | — | — | 19 |
| 9.美洲切叶蚁 Myrmecina americana | — | — | 9 |
| 10.法老小家蚁 Monomorium pharaonis | 6 | 37 | 12 |
| 11.中华小家蚁 Monomorium chinense | 71 | 198 | 109 |
| 12.孟氏大头蚁 Pheidole mendanai | 4 | — | 7 |
| 13.宽结大头蚁 Pheidole nodus | 74 | 175 | 10 |
| 14.台湾瘤颚蚁 Strumigenys formosa | — | — | 1 |
| 15.陕西铺道蚁 Tetramorium shensiense | 10 | 36 | 11 |
| 16.汤加铺道蚁 Tetramorium tonganum | — | 20 | — |
| 17.津岛铺道蚁 Tetramorium tsushimae | 36 | 157 | 600 |
| 四 猛蚁亚科Ponerinae | |||
| 18.中华细鄂猛蚁 Leptogenys chinensis | 7 | — | — |
| 19.山大齿猛蚁 Odontomachus monticola | 8 | — | — |
在“公园与绿地”采集的蚂蚁标本共计813只(101.62只±25.18只,n =8),其中,津岛铺道蚁共600只(75.00只±26.52只,n =8),占“公园与绿地”标本总数的73.80%(66%±10%,n =8);在“乔木林地”中共采集标本257只(32.13只±12.14只,n =8),其中,宽结大头蚁数量最多,共74只(9.25只±6.31只,n =8),占“乔木林地”标本总数的28.79%(18%±6%,n =8);在“旱地”中共采集标本777只(97.13只±34.79只,n =8),其中,中华小家蚁数量最多,共198只(24.75只±11.70只,n =8),占“旱地”标本总数的25.48%(23%±6%,n =8)。津岛铺道蚁在“公园与绿地”蚂蚁群落中的个体数比例显著大于“乔木林地”中的宽结大头蚁和“旱地”中的中华小家蚁的比例( F= 11.651,df =2,P < 0.05)。
2.2 生物多样性比较3种生境间,蚂蚁群落的个体数( F =2.303,df =2,P >0.05)、物种数( F =0.078,df =2,P >0.05)和Shannon-Wiener多样性指数( F= 6.651,df =2,P >0.05)之间的差异均无统计学意义;Simpson多样性指数( F =8.902,df =2,P < 0.05)和Pielou均匀度指数( F= 14.211,df =2,P < 0.05)均为“乔木林地”>“旱地”>“公园与绿地”(表 2)。“旱地”与“乔木林地”的相似度系数为0.500,“公园与绿地”与“乔木林地”的为0.444,均为中等不相似;而“公园与绿地”与“旱地”相似度系数为0.647,为中等相似。
| 生境类型 | 物种数 | Shannon-Wiener 多样性指数 |
Simpson 多样性指数 |
Pielou 均匀度指数 |
| 乔木林地 | 6.38±0.89a | 1.46±0.08a | 0.70±0.02a | 0.83±0.03a |
| 旱地 | 6.25±0.90a | 1.39±0.13ab | 0.69±0.04a | 0.81±0.04a |
| 公园与绿地 | 5.88±1.01a | 0.80±0.19b | 0.39±0.09b | 0.45±0.09b |
| 注:不同上标字母表示生境间蚂蚁群落多样性指数之间的差异有统计学意义 Notes:different superscripts indicate there is a significant difference between the diversity indexes of ant communities in different habitats |
||||
在城市生态系统中,土地的开发利用主要是为人类的生活服务。因此,其原有生境的植被和表层土壤被清除,使其与未开发的自然栖息地截然不同,并具独特性。结果表明,蚂蚁的群落组成在“公园与绿地”“乔木林地”和“旱地”3种城市生境间的差异有统计学意义;其中,津岛铺道蚁在“公园与绿地”中较其他种类显得更为适应,其个体数量在群落中占有明显优势。
蚂蚁物种数以及个体数的采集结果会受采集方式的影响。陷阱法是一种简便、廉价的采集方法(黄杰灵等,2013),在开放生境中具有较高的效率(Melbourne,1999),对生境破坏小。该法能有效采集运动能力强、运动范围广以及在夜间活动的物种(Sabua & Shiju,2010)。尽管曾经有人对陷阱法不能有效收集活动空间极小或活动缓慢的物种表示担忧(Spence & Niemelä,1994)。但相关研究表明,陷阱法依然是采集蚂蚁最理想的方法之一,并且广泛应用于生物多样性的监测工作(Work et al., 2002;Sabua & Shiju,2010)。
城市中不同生境间的结构差异会影响动物群落的物种丰富度及生物多样性(Pacheco & Vasconcelos,2012;Pardee & Philpott,2014;Mata et al., 2017)。特别是良好的植被状况(植被物种多样性和空间异质性等)能够为更多蚂蚁物种提供更丰富和更优质的筑巢地、隐蔽地和食物资源(孟雪松等,2004;Buczkowski & Richmond,2012)。与“公园与绿地”和“旱地”相比,“乔木林地”的木本植物丰富度和林分郁闭度更高,地面也覆盖更多的枝叶凋零物,这可能有助于更多的蚂蚁种类在其中共存。在“公园与绿地”和“旱地”中,草本植被种类则相对单一,植被特征相似度较高;其间农药的广泛使用也可能会导致部分蚂蚁种类消失或个体数量降低(Brittain et al., 2010;Sonoda et al., 2011)。此外,常规耕作行为和土壤扰动也可能成为“旱地”蚂蚁群落生物多样性降低的重要原因(Altieri,1999;Uno et al., 2010)。本研究中,生境类型对蚂蚁群落的影响没有反映在物种和个体的绝对数量上,但“公园与绿地”和“旱地”的蚂蚁多样性指数较“乔木林地”低,“公园与绿地”生境还出现了明显的优势种。
津岛铺道蚁是具有极强竞争力的杂食性、侵略性物种。在人类居住地附近的“公园与绿地”内,津岛铺道蚁往往具有行为和数量上的优势,并有可能通过竞争或剥削的相互作用影响当地蚂蚁群落的多样性(Lessard & Buddle,2005)。目前,关于津岛铺道蚁的生态学资料依然匮乏,其生物学资料、与其他本地蚂蚁物种的竞争机制以及成为优势种的生态后果都还有待进一步研究。
本研究结果证实了蚂蚁群落能够很好地对不同城市生境类型予以响应。在未来的研究中,可以充分利用蚂蚁群落的指示作用,以反映土地利用类型及其景观格局对生态系统功能的影响。在城市化建设的进程中,在满足人类需求的同时,还应该从维护生物多样性的角度出发,更合理地规划土地利用,以构建对人与伴生动物都更宜居的城市生态体系。因此,有必要建立长期的、更广泛的监测研究体系,并涉及更多的城市、生境类型和动物类群,更深入、全面地认识城市化对生物多样性的影响。
曹艳杰. 2006. 《样圆法》在林业规划调查设计中的应用[J]. 内蒙古林业调查设计, 29(2): 39-40. |
陈媛. 2014.中国蚁属16种蚂蚁系统发育及DNA条形码研究[D]. 广西: 广西师范大学.
|
郭萧, 林强, 崔晋波, 等. 2014. 重庆市茶园蚂蚁物种多样性及空间分布格局[J]. 中国生态农业学报, 22(5): 585-593. |
黄杰灵, 胡广, 袁金凤, 等. 2013. 陷阱法和Winkler法调查土壤节肢动物多样性比较:以千岛湖岛屿封闭生境研究为例[J]. 应用昆虫学报, 50(6): 1679-1691. |
李巧, 卢志兴, 张威, 等. 2015. 金沙江干热河谷人工林地表的蚂蚁群落[J]. 林业科学, 51(8): 134-142. |
李巧, 涂璟, 熊忠平, 等. 2011. 节肢动物生物指示研究综述[J]. 西北林学院学报, 26(4): 155-161. |
李淑萍, 徐亚军. 2010. 蚂蚁基因组DNA提取方法比较[J]. 江苏农业科学, (1): 50-51. |
李玉杰, 张晋东, 白文科, 等. 2018. 北京不同功能绿地昆虫群落组成和生物多样性特征[J]. 生态环境学报, 27(6): 1044-1051. |
刘霞, 徐正会, 于娜娜, 等. 2017. 藏东南嘎隆拉和墨脱河谷蚂蚁群落研究[J]. 林业科学研究, 30(1): 34-40. |
梅象信, 徐正会, 张继玲, 等. 2006. 昆明西山森林公园东坡蚂蚁物种多样性研究[J]. 林业科学研究, 19(2): 170-176. |
孟雪松, 欧阳志云, 崔国发, 等. 2004. 北京城市生态系统植物种类构成及其分布特征[J]. 生态学报, 24(10): 2200-2206. |
司建朋, 何跃军, 汪玲, 等. 2019. 喀斯特生境不同功能型植物根系中AM侵染与氮磷含量的关系[J]. 重庆师范大学学报(自然科学版), 36(1): 107-115. |
宋曰钦, 高长宝. 2002. 南方人工林小班蓄积量调查方法比较研究[J]. 江苏林业科技, 29(4): 14-15. |
孙雪, 郑国. 2012. 人为活动对地表节肢动物多样性的影响——以沈阳市北陵公园地表蜘蛛为例[J]. 四川动物, 31(3): 402-405. |
谭速进, 魏翰均, 刘丹碧. 2010. 成都地区居室庭院蚂蚁区系及群落相似性分析[J]. 环境昆虫学报, 32(1): 11-19. |
唐觉, 李参, 黄恩友, 等. 1995. 中国经济昆虫志膜翅目蚁科(一)[M]. 北京: 科学出版社.
|
王维, 沈作奎, 赵玉宏. 2009. 湖北省蚁科昆虫分类研究[M]. 武汉: 中国地质大学出版社.
|
王秀云. 2011.不同年龄长白落叶松人工林碳储量分布特征[D]. 北京: 北京林业大学.
|
吴坚, 王常禄. 1995. 中国蚂蚁[M]. 北京: 中国林业出版社.
|
杨富亿, 吕宪国, 娄彦景, 等. 2014. 松嫩湖群鱼类群落相似性[J]. 生态学报, 35(4): 1022-1036. |
张玮, 郑哲民. 2002. 四川省蚂蚁区系研究(膜翅目:蚁科)[J]. 昆虫分类学报, 24(3): 216-222. |
周善义. 2001. 广西蚂蚁[M]. 广西: 广西师范大学出版社.
|
Alberti M. 2015. Eco-evolutionary dynamics in an urbanizing planet[J]. Trends in Ecology and Evolution, 30(2): 114-126. DOI:10.1016/j.tree.2014.11.007 |
Altieri MA. 1999. The ecological role of biodiversity in agroecosystems[J]. Agriculture Ecosystems and Environment, 74(1-3): 19-31. DOI:10.1016/S0167-8809(99)00028-6 |
Bolton B. 1995. A new general catalogue of the ants of the world[M]. Harvard: Harvard University Press.
|
Brittain CA, Vighi M, Bommarco R, et al. 2010. Impacts of a pesticide on pollinator species richness at different spatial scales[J]. Basic and Applied Ecology, 11(2): 106-115. DOI:10.1016/j.baae.2009.11.007 |
Buczkowski G, Richmond DS. 2012. The effect of urbanization on ant abundance and diversity: a temporal examination of factors affecting biodiversity[J/OL]. PLoS ONE, 7(8): e41729[2018-10-30]. https://doi.org/10.1371/journal.pone.0041729.
|
Folmer O, Black M, Hoeh W, et al. 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit Ⅰ from metazoan invertebrates[J]. Molecular Marine Biology and Biotechnology, 3: 294-299. |
Grimm NB, Faeth SH, Golubiewski NE, et al. 2008. Global change and the ecology of cities[J]. Science, 319: 756-760. DOI:10.1126/science.1150195 |
Hahs AK, Mcdonnell MJ, Mccarthy MA, et al. 2009. A global synthesis of plant extinction rates in urban areas[J]. Ecology Letters, 12(1): 1165-1173. |
Heil M, McKey D. 2003. Protective ant-plant interactions as model systems in ecological and evolutionary research[J]. Annual Review of Ecology, Evolution, and Systematics, 34: 425-453. DOI:10.1146/annurev.ecolsys.34.011802.132410 |
Heraldol V, Marcosf L, Vilhena J, et al. 2010. Ant diversity in an Amazonian savanna: relationship with vegetation structure, disturbance by fire, and dominant ants[J]. Austral Ecology, 33(2): 221-231. |
Hoffmann BD. 2010. Using ants for rangeland monitoring: global patterns in the responses of ant communities to grazing[J]. Ecological Indicators, 10(2): 105-111. DOI:10.1016/j.ecolind.2009.04.016 |
King JR, Andersen AN, Cutter AD. 1998. Ants as bioindicators of habitat disturbance: validation of the functional[J]. Biodiversity and Conservation, 7(12): 1627-1638. DOI:10.1023/A:1008857214743 |
Lessard JP, Buddle CM. 2005. The effects of urbanization on ant assemblages (Hymenoptera: Formicidae) associated with the Molson Nature Reserve, Quebec[J]. The Canadian Entomologist, 137(2): 215-225. DOI:10.4039/n04-055 |
Magnusson WE, deAraújo MC, Cintra R, et al. 1999. Contributions of C3 and C4 plants to higher trophic levels in an Amazonian savanna[J]. Oecologia (Berlin), 119(1): 91-96. |
Mata L, Threlfall CG, Williams NSG, et al. 2017. Conserving herbivorous and predatory insects in urban green spaces[J/OL]. Scientific Reports, 7: 40970 [2018-08-01]. https://doi.org/10.1038/srep40970.
|
McIntyre NE, Rango J, Fagan WF, et al. 2001. Ground arthropod community structure in a heterogenous urban environment[J]. Landscape and Urban Planning, 52(4): 257-274. DOI:10.1016/S0169-2046(00)00122-5 |
Mckinney ML, Lockwood JL. 2001. Biotic homogenization: a sequential and selective process[J]. Biotic Homogenization: 1-17. |
McKinney ML. 2008. Effects of urbanization on species richness: a review of plants and animals[J]. Urban Ecosystems, 11(2): 161-176. DOI:10.1007/s11252-007-0045-4 |
Melbourne BA. 1999. Bias in the effect of habitat structure on pitfall traps: an experimental evaluation[J]. Australian Journal of Ecology, 24(3): 228-239. DOI:10.1046/j.1442-9993.1999.00967.x |
Newbold T, Hudson LN, Hill SL, et al. 2015. Global effects of land use on local terrestrial biodiversity[J]. Nature, 520: 45-50. DOI:10.1038/nature14324 |
Olden JD, Poff NL, Douglas MR, et al. 2004. Ecological and evolutionary consequences of biotic homogenization[J]. Trends in Ecology and Evolution, 19(1): 18-24. DOI:10.1016/j.tree.2003.09.010 |
Pacheco R, Vasconcelos HL. 2012. Habitat diversity enhances ant diversity in a naturally heterogeneous Brazilian landscape[J]. Biodiversity and Conservation, 21(3): 797-809. DOI:10.1007/s10531-011-0221-y |
Pardee GL, Philpott SM. 2014. Native plants are the bee's knees local and landscape predictors of bee richness and abundance in backyard gardens[J]. Urban Ecosystems, 17(3): 641-659. DOI:10.1007/s11252-014-0349-0 |
Pataki DE, Alig RJ, Fung AS, et al. 2006. Urban ecosystems and the North American carbon cycle[J]. Global Change Biology, 12(11): 2092-2102. DOI:10.1111/j.1365-2486.2006.01242.x |
Rocha-Ortega M, Castaño-Meneses G. 2015. Effects of urbanization on the diversity of ant assemblages in tropical dry forests, Mexico[J]. Urban Ecosystems, 18(4): 1373-1388. DOI:10.1007/s11252-015-0446-8 |
Sabua TK, Shiju RT. 2010. Efficacy of pitfall trapping, Winkler and Berlese extraction methods for measuring ground-dwelling arthropods in moist-deciduous forests in the Western Ghats[J]. Journal of Insect Science, 10(98): 1-17. DOI:10.1673/031.010.9801 |
Sonoda S, Izumi Y, Kohara Y, et al. 2011. Effects of pesticide practices on insect biodiversity in peach orchards[J]. Applied Entomology and Zoology, 46(3): 335-342. DOI:10.1007/s13355-011-0041-2 |
Spence JR, Niemelä JK. 1994. Sampling carabid assemblages with pitfall traps: the madness and the method[J]. The Canadian Entomologist, 126(3): 881-894. DOI:10.4039/Ent126881-3 |
Stringer L, Stephens A, Suckling D, et al. 2009. Ant dominance in urban areas[J]. Urban Ecosystems, 12(4): 503-514. DOI:10.1007/s11252-009-0100-4 |
Uno S, Cotton J, Philpott SM. 2010. Diversity, abundance, and species composition of ants in urban green spaces[J]. Urban Ecosystems, 13(4): 425-441. DOI:10.1007/s11252-010-0136-5 |
Work TT, Buddle CM, Korinus LM, et al. 2002. Pitfall trap size and capture of three taxa of litter-dwelling arthropods: implications for biodiversity studies[J]. Environmental Entomology, 31(3): 438-448. DOI:10.1603/0046-225X-31.3.438 |
Xu X, Li Y, Wang B, et al. 2015. Salt stress induced sex-related spatial heterogeneity of gas exchange rates over the leaf surface in Populus cathayana Rehd.[J/OL]. Acta Physiologiae Plantarum, 37(1): 1709 [2018-10-30]. https://doi.org/10.1007/s11738-014-1709-3.
|