 2020, Vol. 39
2020, Vol. 39扩展功能
文章信息
- 陶星宇, 翟晓飞, 王同亮, 崔硕, 汪继超
- TAO Xingyu, ZHAI Xiaofei, WANG Tongliang, CUI Shuo, WANG Jichao
- 海南睑虎种群密度调查研究
- Estimation of the Population Density of Goniurosaurus hainanensis
- 四川动物, 2020, 39(1): 63-67
- Sichuan Journal of Zoology, 2020, 39(1): 63-67
- 10.11984/j.issn.1000-7083.20190337
-
文章历史
- 收稿日期: 2019-10-07
- 接受日期: 2019-12-10






海南睑虎Goniurosaurus hainanensis隶属于爬行纲Reptilia有鳞目Squamata睑虎科Eublepharidae睑虎属Goniurosaurus,为海南省特有种,首次发现于海南省五指山。文献记载该物种分布在我国海南岛的陵水、保亭、海口和琼中(Grismer et al., 2002;Blair et al., 2009;张汾,李友邦,2017)。近年来,由于海南睑虎栖息地破碎化、分布区狭窄以及宠物市场的大量需求等因素,该物种野外种群资源受到极大威胁(齐旭明等,2011),已被《中国濒危动物红皮书(第二卷):脊椎动物(下册)》列为易危(VU)等级(汪松,解焱,2009)。
目前关于睑虎属物种的研究主要集中在新种的描述和系统进化等方面(Grismer et al., 2002;Wang et al., 2019;Zhai et al., 2019),种群密度量化研究较少。本课题组前期对霸王岭睑虎G. bawanglingensis种群密度开展了相关研究,为该物种资源现状评估提供了基础数据(陈一帆等,2019)。目前有关海南睑虎的研究也主要集中在分类分布方面(Grismer et al., 2002;Blair et al., 2009;张洁等,2014;Liang et al., 2018),关于其野生种群密度和基础生态学资料基本空白,掌握物种种群密度及其动态变化对于物种资源现状评估和保护具有重要意义。野生动物种群数量及密度的有效评估较为复杂,不同物种的种群密度调查和统计的方法也有所不同(阳春生等,2017)。种群密度调查研究中常用的方法有样区法和标记重捕法,其中标记重捕法常见的种群密度估算方法有:Lincoln指数法、Schnabel法和Jolly-Seber法(孙儒泳,2001;张旭等,2012)。鉴于此,2019年7—9月,在海南吊罗山国家级自然保护区采用标记重捕法对海南睑虎种群密度开展了调查研究,以期为掌握该物种种群动态、评估资源现状和保护提供依据。
1 材料与方法 1.1 研究区域海南吊罗山国家级自然保护区地跨陵水、保亭和琼中3个县,地理坐标109°45′05″~109°57′07″E、18°40′08″~18°49′19″N,面积18 398 hm2,其中林地面积18 356.34 hm2,森林覆盖率99.82%,是海南四大林区之一。林区干湿季节分明,年均降水量为2 160 mm,每年的5月底到10月底为雨季,降水量占全年的四分之三;11月初至翌年5月初为旱季(邱治军等,2004;韩天宇等,2019)。
根据前期海南睑虎野外分布的预调查结果,在研究区域内选取了2个不同海拔区间的样区,分别标注为A样区(海拔551~688 m,长3 000 m,宽2 m)和B样区(海拔289~448 m,长3 000 m,宽2 m)。出于对物种的保护,调查样区具体地理位置不予公布。
1.2 调查方法根据海南睑虎的生活习性,采用样线法于每天20: 00—20: 30开始调查,从样线起点保持匀速前行,对沿途发现的海南睑虎进行捕捉标记。发现个体采用植入式动物电子标签(1.41 mm×8.26 mm,0.01 g;HT157,广州洪腾条码技术有限公司)进行标记:将植入式动物电子标签经专用标签注射器植入动物距泄殖腔前1~2 cm处的皮下,经前期预实验检测该标签可长期保存,且容易识别,对动物的日常活动影响较小。个体标记完毕后,记录被标记个体的标签编号和标记时间等信息后原地释放。在重捕调查过程中,对观察到的个体进行捕捉,并用读码扫描仪对其进行识别、记录编号后原地释放(陈一帆等,2019)。
1.3 数据处理分别采用Schnabel法和Lincoln指数法估算海南睑虎种群密度(孙儒泳,2001;张旭等,2012)。
1.3.1 Schnabel法中的定义与术语种群数量:
1/N方差:
1/N标准误:
1/N 95%置信区间为1/N±t0.05SE1/N
式中,a为重捕总次数;i为取样次数(i=1,2,3,……);Ci为第i次取样时发现个体的总数(其中包含未捕捉个体);Ri为第i次取样时捕获的个体数中已经标记个体的总数;Ui为第i次取样时新标记并释放的个体总数;Mi为第i次取样时种群中已标记的个体总数。
1.3.2 Lincoln指数法中的定义与术语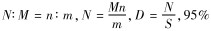

式中,D为种群密度;S为样区面积;N为该地段中全部个体数;M为标记个体数量;n为重捕个体数量;m为重捕个体中有标记个体数量。
2 结果 2.1 Schnabel法A样区共进行了6次标记9次重捕(表 1),共标记50只,种群数量为301只,种群数量95%置信区间为301只±36只,种群密度为501只/hm2;B样区共进行了6次标记9次重捕(表 2),共标记229只,种群数量为1 200只,种群数量95%置信区间为1 200只±332只,种群密度为1 999只/hm2。
| 重捕次数 | Ci/只 | Ri/只 | Ui/只 | Mi/只 | CiMi2 | RiMi | N±t0.05SE/只 | D/(只/hm2) |
| 0 | 16 | 0 | 11 | 0 | 0 | 0 | ||
| 1 | 7 | 0 | 6 | 11 | 847 | 0 | ||
| 2 | 6 | 1 | 5 | 17 | 1 734 | 17 | 152±80 | 253 |
| 3 | 16 | 1 | 15 | 22 | 7 744 | 22 | 265±63 | 441 |
| 4 | 6 | 1 | 4 | 37 | 8 214 | 37 | 244±42 | 407 |
| 5 | 8 | 2 | 5 | 41 | 13 448 | 82 | 202±43 | 337 |
| 6 | 5 | 0 | 4 | 46 | 10 580 | 0 | 269±38 | 499 |
| 7 | 4 | 1 | 0 | 50 | 10 000 | 50 | 253±35 | 421 |
| 8 | 5 | 1 | 0 | 50 | 12 500 | 50 | 252±35 | 420 |
| 9 | 5 | 0 | 0 | 50 | 12 500 | 0 | 301±36 | 501 |
| 重捕次数 | Ci/只 | Ri/只 | Ui/只 | Mi/只 | CiMi2 | RiMi | N±t0.05SE/只 | D/(只/hm2) |
| 0 | 27 | 0 | 25 | 25 | 16 875 | 0 | ||
| 1 | 36 | 2 | 29 | 54 | 104 976 | 108 | ||
| 2 | 42 | 5 | 26 | 80 | 268 800 | 400 | 769±496 | 1 282 |
| 3 | 52 | 6 | 34 | 114 | 675 792 | 684 | 895±280 | 1 491 |
| 4 | 81 | 14 | 53 | 152 | 1 871 424 | 2 128 | 895±336 | 1 475 |
| 5 | 56 | 5 | 38 | 206 | 2 376 416 | 1 030 | 1 222±325 | 2 036 |
| 6 | 45 | 13 | 24 | 229 | 2 359 845 | 2 977 | 1 047±306 | 1 746 |
| 7 | 47 | 10 | 0 | 229 | 2 464 727 | 2 290 | 1 054±309 | 1 757 |
| 8 | 53 | 7 | 0 | 229 | 2 779 373 | 1 603 | 1 151±324 | 1 919 |
| 9 | 47 | 7 | 0 | 229 | 2 464 727 | 1 603 | 1 200±332 | 1 999 |
A样区进行7次标记3次重捕,共标记个体50只,重捕个体共14只,重捕个体中有标记个体数量为2只,种群数量为350只,种群数量95%置信区间为350只±450只,种群密度为583只/hm2;B样区进行7次标记3次重捕,共标记个体229只,重捕个体共147只,重捕个体中有标记个体数量为40只,种群数量为842只,种群数量95%置信区间为842只±202只,种群密度为1 403只/hm2(表 3)。
| 样区 | 标记个体数量 M |
重捕个体数量 n |
重捕个体中有标记个体数量 m |
95%置信区间 N±2S·E |
种群密度D/(只/hm2) |
| A样区 | 50 | 14 | 2 | 350±450 | 583 |
| B样区 | 229 | 147 | 40 | 842±202 | 1 403 |
标记重捕法常用于爬行动物种群生态学研究(阳春生等,2017),对不同物种的种群数量及密度估算时采用的算法也有所不同(丁岩钦,1994;吴鹏飞等,2005;张旭等,2012)。在调查过程中发现,海南睑虎野外种群分布不均匀,这可能导致标记个体与未标记个体被捕的机会不同,且低密度区的重捕率较低。因此在采用Lincoln指数法估算A样区种群密度时置信区间范围过大,无法保证估算结果的精确性。而采用Schnabel法对海南睑虎种群密度进行估算时较Lincoln指数法估算的结果更高,在重捕到第6、7次时,估算的种群数量和置信区间结果开始趋于稳定。本研究结果认为,在海南睑虎种群密度估算中Schnabel法较为适用,标记重捕次数达到6次即能满足数据分析要求。
种群密度是种群生态学研究中的重要内容,是反应种群消长及资源现状的重要指标,对濒危物种科学评估、保护和管理具有重要意义(龚世平等,2006;Stephens et al., 2006)。本研究首次对海南吊罗山国家级自然保护区的海南睑虎种群密度进行了调查,结果显示,A样区种群密度为501只/hm2,B样区种群密度为1 999只/hm2。文陇英(2007)调查兰州郊区的丽斑麻蜥Eremias argus和草原沙蜥Phrynocephalus frontalis种群密度分别为129只/hm2和100只/hm2。而本研究中海南睑虎种群密度较高,可能与不同物种特性和栖息地环境差异有关。热带地区独特的气候特点,孕育了丰富而独特的生物多样性资源,植被类型也更为丰富(黄勇,王跃招,2011)。本次调查研究区域属于典型的热带雨林,雨水充足,动物食物丰富度较高,对于营穴居生活的睑虎属物种具有较高的适合性。
海拔综合了温度、降水等气候因子,也包含了植被类型、植物丰富度等生态因子,对动物的分布会产生较大的影响(McCain & Grytnes,2001;Barry,2008;王同亮等,2015;陶夏秋等,2018)。本研究结果显示,海南吊罗山国家级自然保护区海南睑虎种群密度随着海拔的增高而减少,与陈一帆等(2019)对霸王岭睑虎种群密度调查的结果(845只/hm2,海拔180~307 m;591只/hm2,海拔329~382 m)相似,但影响睑虎属物种种群密度海拔分布差异的因素尚有待进一步调查研究。
本研究发现,人为活动和干扰较强的低海拔区为海南睑虎的主要分布区,但槟榔Areca catechu和龙眼Dimocarpus longan等经济作物的种植一定程度上导致了栖息地质量下降和破碎化。此外,调查过程中也发现海南睑虎被车辆碾压致死的现象,海南睑虎主要活动时间在夜间,因此建议尽量减少夜间车辆行驶,降低人为干扰对该物种的影响。
致谢: 项目执行得到海南省野生动植物保护管理局莫燕妮局长、王世力科长和齐旭明科长等人的大力支持;实验得到海南师范大学博士研究生卜荣平、硕士研究生陈一帆、李汉东的协助;文章修改得到海南师范大学郭蕊老师的宝贵意见和建议;野外工作和生活得到海南吊罗山国家级自然保护区管理局工作人员的帮助和照顾,在此一并表示感谢。
陈一帆, 翟晓飞, 陶星宇, 等. 2019. 标记重捕法对模式产地霸王岭睑虎种群资源调查[J]. 四川动物, 38(4): 420-424. |
丁岩钦. 1994. 昆虫数学生态学[M]. 北京: 科学出版社.
|
龚世平, 史海涛, 陈川, 等. 2006. 海南岛黎母山四眼斑水龟种群密度与空间分布格局[J]. 动物学杂志, 41(6): 54-59. |
韩天宇, 沈燕, 王旭, 等. 2019. 海南吊罗山低地雨林群落特征分析[J]. 林业与环境科学, 35(3): 43-49. |
黄勇, 王跃招. 2011. 海南岛两栖爬行动物生物地理学研究进展[J]. 四川动物, 30(2): 304-309. |
齐旭明, 田蜜, 李小成, 等. 2011. 霸王岭睑虎种群及数量状况研究[J]. 热带林业, 39(1): 47-49. |
邱治军, 刘海伟, 李桂梅, 等. 2004. 海南省吊罗山自然保护区水文条件与水资源[J]. 热带林业, 32(2): 34-37. |
孙儒泳. 2001. 动物生态学原理(第三版)[M]. 北京: 北京师范大学出版社.
|
陶夏秋, 崔绍朋, 蒋志刚, 等. 2018. 新疆阿勒泰地区爬行动物区系及多样性海拔分布格局[J]. 生物多样性, 26(6): 578-589. |
汪松, 解焱. 2009. 中国物种红色名录(第二卷):脊椎动物(下册)[M]. 北京: 高等教育出版社.
|
王同亮, 程林, 兰文军, 等. 2015. 江西武夷山国家级自然保护区两栖动物多样性及海拔分布特点[J]. 生态学杂志, 34(7): 2009-2014. |
文陇英. 2007. 丽斑麻蜥和草原沙蜥的种群调查[J]. 甘肃科学学报, 19(1): 88-90. |
吴鹏飞, 曾宗永, 王跃招, 等. 2005. 青海沙蜥种群密度调查的一种新方法[J]. 生态学杂志, 24(10): 1241-1244. |
阳春生, 罗树毅, 李钰慧, 等. 2017. 样线法和标志重捕法在鳄蜥种群数量调查中的应用比较[J]. 野生动物学报, 8(2): 291-294. |
张汾, 李友邦. 2017. 我国睑虎属的分类和保护研究进展[J]. 安徽农业科学, 45(1): 1-3, 7. |
张洁, 庆宁, 易祖盛, 等. 2014. Goniurosaurus indet.的有效性及中国睑虎属(Squamata:Sauria:Eublepharidae)种间的亲缘关系[J]. 华南师范大学学报(自然科学版), 6(2): 92-98. |
张旭, 鲍毅新, 刘军, 等. 2012. 陆桥岛屿环境下社鼠种群数量的估算方法[J]. 生态学报, 32(5): 1562-1569. |
Barry RG. 2008. Mountain weather and climate[M]. New York: Cambridge University Press.
|
Blair C, Orlov NL, Shi HT, et al. 2009. A taxonomic re-evaluation of Goniurosaurus hainanensis (Squamata:Eublepharidae) from Hainan Island, China[J]. Russian Journal of Herpetology, 16(1): 35-40. |
Grismer LL, Shi HT, Orlov NL, et al. 2002. A new species of Goniurosaurus (Squamata:Eublepharidae) from Hainan Island, China[J]. Journal of Herpetology, 36(2): 217-224. |
Liang B, Zhou RB, Liu YL, et al. 2018. Renewed classification within Goniurosaurus (Squamata:Eublepharidae) uncovers the dual roles of a continental island (Hainan) in species evolution[J]. Molecular Phylogenetics and Evolution, 127: 646-654. |
McCain CM, Grytnes JA. 2001. Elevational gradients in species richness[M]. Chichester, UK: John Wiley & Sons.
|
Stephens PA, Zaumyslova OY, Miquelle DG, et al. 2006. Estimating population density from indirect sign:track counts and the Formozov-Malyshev-Pereleshin formula[J]. Animal Conservation, 9(3): 339-348. |
Wang JC, Chen YF, Zhai XF, et al. 2019. Complete mitochondrial genome of Goniurosaurus zhoui (Squamata:Sauria:Eublepharidae)[J]. Mitochondrial DNA Part B, 4(2): 3218-3219. |
Zhai XF, Chen YF, Wang TL, et al. 2019. Complete mitochondrial genome of Goniurosaurus bawanglingensis (Squamata:Sauria:Eublepharidae)[J]. Mitochondrial DNA Part B, 4(1): 844-845. |