 2019, Vol. 38
2019, Vol. 38扩展功能
文章信息
- 游卫云, 高欣, 梅爱君, 徐卫南, 张先福, 鲁庆斌
- YOU Weiyun, GAO Xin, MEI Aijun, XU Weinan, ZHANG Xianfu, LU Qingbin
- 基于红外触发相机技术的白鹇栖息地选择研究
- Habitat Selection Research of Lophura nycthemera Using Infrared-Triggered Camera Technology
- 四川动物, 2019, 38(5): 564-570
- Sichuan Journal of Zoology, 2019, 38(5): 564-570
- 10.11984/j.issn.1000-7083.20190078
-
文章历史
- 收稿日期: 2019-03-04
- 接受日期: 2019-07-08






2. 杭州市临安区林业局, 杭州 311300
2. Forestry Bureau of Lin'an District, Hangzhou 311300, China
白鹇Lophura nycthemera主要分布于中国、东南亚以及南亚的部分地区,在我国分布于南方各省的山地林区,为国家Ⅱ级重点保护野生鸟类(Delacour,1977;谭耀匡,吴至康,1981;郑作新,1987)。白鹇面临栖息地的破坏、丧失和片段化等威胁(郑家文等,2006),种群数量下降(卢汰春,1991;高育仁,余德群,1995),因此栖息地保护是关键。有关白鹇种群的数量分布、食性、行为和繁殖等方面的研究已有报道(丁平等,1992;高育仁,1993;尹祚华,1995;熊志斌等,2002;刘选珍等,2013);栖息地选择也有相应的研究报道(邵晨,胡一中,2005;郑家文等,2006;张娟茹等,2010;黄清山,2015),但传统方法在获取数据时可能会对白鹇的活动行为产生不同程度的干扰,研究结果的可靠性值得商榷。红外触发相机对动物几无干扰,有连续性及记录时间长等特点,应用于野生动物研究具有明显的优势,包括调查时间和人力的限制少、可获取全天候数据、对动物影响小等(李晟等,2014)。红外触发相机技术已被应用于物种多样性、行为及生境利用等野生动物研究方面(Schipper,2007;张履冰等,2014;赵玉泽等,2016),但栖息地选择方面的报道较少。鉴于此,笔者利用红外触发相机技术,以浙西山区分布的白鹇作为研究对象,不仅补充和完善白鹇栖息地选择的研究成果,也探讨该技术在野生动物栖息地选择研究中的应用,促进野生动物保护研究工作的发展。
1 研究地区研究地区位于浙江省西北部临安区北部(118°51′~119°52′E,29°56′~30°23′N),东西宽约80 km,南北长约40 km,总面积约3 126.8 km2。境内低山丘陵与河谷盆地相间排列,交错分布,西北多崇山峻岭,深沟幽谷,整体称为天目山系,向北绵延至安徽省境内,西部的清凉峰为最高峰,海拔1 787 m;属季风型气候,温暖湿润,光照充足,雨量充沛,四季分明;年均降水量1 390~1 870 mm,年均无霜期209~237 d,年均气温9~16 ℃;主要溪流有西坑溪、东关溪、丰陵溪和昌北溪,分别注入太湖水系和钱塘江水系;森林覆盖率高,有复杂的动植物区系成分和独特的环境资源(程渭山,2002;吕建中,2009)。
2 研究方法 2.1 相机布设2015年8月—2016年9月,根据预调查确定有白鹇活动的山区,选择其中7个地点(太湖源镇的东坑和阳山坪、天目山国家级自然保护区的东关和西关、清凉峰国家级自然保护区的千顷塘和童玉、大峡谷镇的叶家)作为调查样地(图 1),样地之间的间距不低于5 km。由于白鹇是留鸟,且边食边移动,常早晨朝上坡走(吴名川,1988),因此由山脚到山顶设置样线,样线直线长度为1 500~2 000 m。沿样线在有粪便、爪印等痕迹的位置(可能是白鹇活动或适合白鹇活动的场所)安放红外触发相机(Bestguarder SG-990V,1 200万像素),固定在距地面30~60 cm的树干或其他固定物上,相机镜头与坡面平行,以取景最佳为原则,设置为3次连拍,拍摄间隔时间为3 s。相邻2台相机之间的距离大于100 m。

|
| 图 1 调查样地 Fig. 1 The location of study area |
| |
相机数量和安放时间:天目山国家级自然保护区36台,2015年8月25日—10月15日在东关安放了18台,2015年10月25日—12月12日在西关安放了18台;大峡谷镇20台,2016年1月19日—2月27日安放在叶家;太湖源镇36台,2016年2月20日—4月13日在东坑安放了18台,2016年3月27日—5月13日在阳山坪安放了18台;清凉峰国家级自然保护区35台,2016年5月21日—6月23日在千顷塘安放了18台,2016年7月16日—8月23日在童玉安放了17台。
2.2 数据采集以相机为中心,设置1个10 m×10 m白鹇可利用栖息地的大样方,正方形的一边处于相同海拔;在样地中心和四角位置分别各设1个2 m×2 m的中样方和1个1 m×1 m的小样方,前者用于测量灌木指标,后者用于测量草本指标。参照有关资料(石建斌,郑光美,1997;Skarin et al., 2004;孟秀祥等,2010)和实际情况测定生态因子:
海拔(m):相机所处地的海拔;
植被类型:根据大样方中优势种的种类判定,分为:阔叶林、针阔混交林、针叶林、灌丛、竹林;
郁闭度(%):大样方中心点4个方向的植被上层林冠对地面覆盖百分比的平均值;
乔木密度:大样方内乔木(株高≥6 m)的数量;
灌木密度:5个中样方内灌木(株高<6 m)的平均数量,分为:少(<20株)、中(20~40株)、多(≥40株);
草本密度:5个小样方内草本的平均数量,分为:少(<50株)、中(50~100株)、多(≥100株);
地面落叶盖度:5个小样方内落叶的平均量(以落叶覆盖地面的百分率计算),分为:少(<20%)、中(20%~80%)、多(≥80%);
坡向:东北坡(22.5~67.5°)、东坡(67.5~112.5°)、东南坡(112.5~157.5°)、南坡(157.5~202.5°)、西南坡(202.5~247.5°)、西坡(247.5~292.5°)、西北坡(292.5~337.5°)、北坡(337.5~22.5°);
坡度(°):大样方所在坡面与水平地面所成的角度;
坡位:坡下位(含山谷)、坡中位(含山腰)、坡上位(含山脊);
离水源距离(m):大样方中心到最近水源(泉水及河溪等水体,不含积雪)的直线距离;
离公路距离(m):大样方中心到最近道路(供机动车行驶)的直线距离;
人为干扰程度:大样方所在区域人为活动情况,包括足迹(人和家畜的足迹)、废物和噪声(人声、车辆声等),分为3个等级:小(以上三者均无)、中(三者不全有)、大(三者全有)。
2.3 数据处理采用非配对设计2个样本平均数的t检验比较白鹇选择的栖息地与可利用的栖息地之间的差异性。可利用的栖息地是指白鹇可能活动、采食等的生活场所,即所有相机的安放点。白鹇选择的栖息地是指其实际利用的生活场所,即相机记录到白鹇活动的安放点。首先进行正态分布检验,符合正态分布的数据进行方差齐性检验,按方差相等或不等的条件进行不同的t检验。显著性水平设置为α=0.05。统计分析采用SPSS 19进行。
用χ2适合度检验比较各类生态因子的实际观察值与理论值的差异性,当差异有统计学意义或高度统计学意义时,再以下式进行区间分析。在置信水平为α时,动物对某生态因子其中一个分类等级的实际利用比率pi的置信区间为(Koehler & Larntz,1980):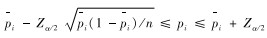
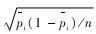
127台相机中有27台拍到了白鹇,共180张相片。对白鹇栖息地中的生态因子进行t检验,白鹇对坡度(P<0.05)和离水源距离(P≤0.01)有选择性,而对海拔(P>0.05)、乔木密度(P>0.05)、离公路距离(P>0.05)和郁闭度(P>0.05)无选择性(表 1)。白鹇选择坡度适中(26.30 ℃±4.20 ℃)、离水源距离较近(48.44 m±15.64 m)的栖息地。
| 生态因子 Ecological factors |
可利用栖息地 Available habitats |
选择的栖息地 Selected habitats |
t | df | P |
| 海拔/m | 772.74 | 697.81 | 1.414 | 45 | >0.05 |
| 乔木密度 | 8.80 | 8.33 | 0.540 | 152 | >0.05 |
| 坡度/° | 21.30 | 26.30 | -1.992 | 152 | <0.05 |
| 离公路距离/m | 931.54 | 996.30 | -0.939 | 52 | >0.05 |
| 离水源距离/m | 97.43 | 48.44 | 2.558 | 152 | ≤0.01 |
| 郁闭度/% | 0.69 | 0.71 | -0.473 | 152 | >0.05 |
对白鹇栖息地中的计数资料进行χ2检验,白鹇对植被类型(P<0.01)、坡位(P<0.01)、坡向(P<0.01)、灌木密度(P<0.01)、草本密度(P<0.01)、地面落叶盖度(P<0.01)和人为干扰程度(P<0.01)等均有极显著的选择性(表 2)。因此,对这些生态因子继续进行适合度分析。
| 生态因子 Ecological factors |
χ2 | df | P |
| 植被类型 | 82.14 | 4 | <0.01 |
| 坡位 | 79.45 | 2 | <0.01 |
| 坡向 | 79.08 | 7 | <0.01 |
| 灌木密度 | 80.11 | 2 | <0.01 |
| 草本密度 | 80.07 | 2 | <0.01 |
| 地面落叶盖度 | 79.95 | 2 | <0.01 |
| 人为干扰程度 | 79.47 | 2 | <0.01 |
结果表明,白鹇对植被类型的选择性,表现为选择针阔混交林,回避针叶林和阔叶林,而对灌丛林和竹林随机选择。在坡位方面,白鹇回避下坡位,而对上坡位和中坡位随机选择。在坡向方面,白鹇回避西坡,而其他坡向随机选择。在灌丛密度方面,白鹇选择灌丛密度小的栖息地,回避灌丛密度大的栖息地。在草本密度方面,白鹇选择草本密度适中的栖息地,回避草本密度大的栖息地。在地面落叶盖度方面,白鹇选择地面落叶盖度大的栖息地。在人为干扰程度方面,白鹇选择选择人为干扰小的栖息地,回避人为干扰大的栖息地,对人为干扰中等的栖息地随机选择。
综上所述,白鹇栖息地选择主要与食物、水源和人为干扰等密切相关,结合实际调查数据发现,它们主要选择坡度较小、离水源较近的针阔混交林活动,且这些区域灌木较少(<20株/4m2)、草本适中(50~100株/m2)和落叶较多(≥80%)。
4 讨论适宜的栖息地是野生动物赖以生存的基本条件,可满足动物的基本生理需求,如食物提供、反捕食及保温等,从而使其存活适合度及繁殖适合度最大化(Rosenzweig,1985;Beest et al., 2010)。利用红外触发相机技术开展白鹇栖息地选择研究与传统方法比较(表 4),在可比较的8个生态因子中,植被类型、坡度、离水源距离、灌木密度、地面落叶盖度、人为干扰程度6个相同,占75%;而坡位和郁闭度略有不同,占25%。由于红外触发相机技术具有无干扰性,可以更为真实客观地获得第一手资料(Wegge et al., 2004;Schipper,2007),因此研究结果的可信性高。如本研究发现,有95%白鹇偏好的坡度为22~30.5°,与邵晨和胡一中(2005)的研究结果(30~45°)不同,可能是因为夜栖地只是栖息地的一部分。
| 生态因子 Ecological factor |
分类等级 Classification level |
可利用栖息地占比 Available habitat proportion |
选择的栖息地占比 Selected habitat proportion |
置信区间 Confidence interval |
| 植被类型 | 阔叶林 | 0.283 | 0.074 | 0≤P1≤0.173* |
| 针阔混交林 | 0.346 | 0.556 | 0.368≤P2≤0.743* | |
| 针叶林 | 0.283 | 0.148 | 0.014≤P3≤0.282* | |
| 灌木林 | 0.047 | 0.148 | 0.014≤P4≤0.282 | |
| 竹林 | 0.039 | 0.074 | 0≤P5≤0.173 | |
| 坡位 | 下坡位 | 0.299 | 0.148 | 0.014≤P1≤0.282* |
| 中坡位 | 0.354 | 0.481 | 0.293≤P2≤0.670 | |
| 上坡位 | 0.346 | 0.370 | 0.188≤P3≤0.553 | |
| 坡向 | 北 | 0.039 | 0.074 | 0≤P1≤0.173 |
| 东北 | 0.063 | 0.037 | 0≤P2≤0.108 | |
| 东 | 0.315 | 0.370 | 0.188≤P3≤0.553 | |
| 东南 | 0.102 | 0.185 | 0.039≤P4≤0.332 | |
| 南 | 0.157 | 0.148 | 0.014≤P5≤0.282 | |
| 西南 | 0.142 | 0.148 | 0.014≤P6≤0.282 | |
| 西 | 0.214 | 0.037 | 0≤P7≤0.108* | |
| 西北 | 0.010 | 0 | — | |
| 灌木密度 | 小 | 0.323 | 0.519 | 0.330≤P1≤0.707* |
| 中 | 0.323 | 0.333 | 0.156≤P2≤0.511 | |
| 大 | 0.354 | 0.148 | 0.014≤P3≤0.282* | |
| 草本密度 | 小 | 0.496 | 0.444 | 0.257≤P1≤0.632 |
| 中 | 0.252 | 0.444 | 0.257≤P2≤0.632* | |
| 大 | 0.252 | 0.111 | 0≤P3≤0.230* | |
| 地面落叶盖度 | 小 | 0.354 | 0.259 | 0.094≤P1≤0.425 |
| 中 | 0.339 | 0.222 | 0.065≤P2≤0.379 | |
| 大 | 0.307 | 0.519 | 0.330≤P3≤0.707* | |
| 人为干扰程度 | 小 | 0.732 | 0.889 | 0.770≤P1≤1.000* |
| 中 | 0.150 | 0.074 | 0≤P2≤0.173 | |
| 大 | 0.118 | 0.037 | 0≤P3≤0.108* | |
| 注:Pi为动物对某生态因子其中一个分类等级实际利用比率的置信区间 Note:Pi is the confidence interval of the actual utilization ratio at one classification of an ecological factor | ||||
| 数据来源 Resource |
生境类型 Habitat type |
植被类型 Vegetation type |
坡位 Slope |
坡度 Gradient |
离水源距离 Water distance/m |
灌木密度 Shrub density |
地面落叶盖度 Fallen leaf density |
郁闭度 Canopy density/% |
人为干扰程度 Human disturbance |
| 丁平等,1992 | 5种生境 | 针阔混交林 | — | — | — | 小 | — | ≥80 | — |
| 熊志斌等,2003 | 栖息地 | 针阔混交林 | 随季节变化 | — | 附近 | 稀少 | 大 | ≥80 | 小 |
| 邵晨,胡一中,2005 | 夜栖地 | 阔叶林 | 沟谷 | 30~45° | — | — | — | — | — |
| 张娟茹等,2010 | 巢址 | — | — | 较小 | 较近(15.53 m) | — | — | 81.90 | 小 |
| 黄清山,2015 | 栖息地 | 针叶林和灌木林 | 沟谷 | 较平缓(<45°) | — | 小 | — | — | — |
| 本研究 | 栖息地 | 针阔混交林 | 回避下坡位 | 26.30°±4.20° | 较近(48.44 m±15.64 m) | 小 | 大 | 无选择性 | 小 |
邵晨和胡一中(2005)、黄清山(2015)的研究认为,白鹇选择沟谷为夜栖地或栖息地;熊志斌等(2003)认为,白鹇对坡位的选择随季节而变化,春季主要选择距离山脊的栖息地,冬季偏向于选择坡下位的栖息地。本研究发现,白鹇回避下坡位的栖息地。导致这一差异的原因可能是环境因素的影响。本研究区域内有2个国家级自然保护区,该地区是驰名中外的风景名胜区,具有优美的自然风光和历史文化传承,所以白鹇的栖息地成为当地政府大力发展生态旅游的重点区域。这对生态系统产生了一定程度的负面影响(曹敏鲁等,1999;鲁庆彬等,2001)。实地调查发现,当地的餐饮经营存在采集野菜、偷捕野生动物等行为,自驾游很普遍,并带来一定的噪声和交通拥挤,驴友组团开展森林观光和探险等活动也很常见,但这些事件主要发生在山谷、低山等人为活动比较频繁的区域。因此,上述情况对白鹇栖息地选择可能形成干扰,从而影响白鹇对坡位的选择性。此外,对“沟谷”和“坡位”的涵义有待考证,本文认为沟谷并不限于下坡位,因为山区溪流总是从山上蜿蜒曲折流向山下,流经区域似乎均可称为沟谷;从另一方面来说,沟谷包括山溪和谷地,而“沟”者显然包括所有坡位,“谷”者仅仅包括下坡位。本研究证明,白鹇选择离水源较近的栖息地,间接说明白鹇对山溪有一定的选择性。因此,本文的坡位选择研究结果似乎与前述作者的研究结果部分相同。
除此之外,食物因子也是动物生境选择最重要的影响因素(马建章,1990;Anoop & Hussain,2004)。白鹇为杂食性,主要以植物的嫩叶、幼芽、花、茎、浆果、种子、根和苔藓等为食,也取食昆虫、蜗牛等动物性食物(赵正阶,2001)。从植物性食物看,果实产量与光照强度存在线性回归关系,但最大光照强度不超过70%(Palmer,1989;Wagenmakers & Tazelaar,1999)。这说明光照强度太高或郁闭度太低的环境,果实产量不高,所以白鹇最丰富的植物性食物资源需要较高的郁闭度;但是,郁闭度过高不利于林下植物生长,其嫩叶、幼芽、花、茎、浆果、种子均不丰富。从动物性食物看,土壤动物(蚁类、蚯蚓、蜗牛等)对生存环境表现为负趋光性(程量,1987),故较高的郁闭度和植物密度才能满足条件。因此,白鹇对食物因子的选择性应该表现为较高的郁闭度和植物密度。基于以上分析,白鹇选择郁闭度大于80%的栖息地(丁平等,1992;熊志斌等,2003)的结论似乎过于笼统。本研究发现,白鹇对郁闭度无显著的选择性,但从被选择的栖息地看,郁闭度为50%~85%,与上述结论较为一致。
综上所述,保护与利用是相互矛盾的问题,资源的利用始终会对野生动物造成干扰,解决该问题的关键在于开展资源的可持续利用。区域的可持续发展与生态保护紧密关联(Potschin & Haines-Young,2013;Turner et al., 2013),故白鹇种群及其栖息地保护是当地可持续发展的前提和基础条件之一。由于白鹇强烈回避人为干扰强度大的栖息地,因此,有关部门在进行旅游开发时,应专门针对白鹇及其栖息地进行保护管理。可在熟悉其生态习性的基础上,规划旅游开发的开放和非开放区域,制定政策措施限制非开放区域内的旅游活动,使其种群及核心栖息地免受人为干扰的影响。
| 曹敏鲁, 安琪林, 毅红. 1999. 生态旅游指南[M]. 北京: 经济日报出版社. |
| 程量. 1987. 蚁类在不同生境中摄食活动规律的研究[J]. 动物学研究, 8(2): 133–141. |
| 程渭山. 2002. 浙江林业自然资源[M]. 北京: 中国农业科学技术出版社. |
| 丁平, 姜仕仁, 石斌山, 等. 1992. 浙江西部山区白鹇生态的初步研究[J]. 动物学杂志, 27(3): 20–23. |
| 高育仁, 余德群. 1995. 白鹇海南亚种的生态和现状[J]. 动物学研究, 16(4): 353–358. |
| 高育仁. 1993. 白鹇食性研究[J]. 动物学研究, 14(2): 194–196. |
| 黄清山. 2015. 福建茫荡山自然保护区白鹇栖息地研究[J]. 现代农业科技(6): 260–261. DOI:10.3969/j.issn.1007-5739.2015.06.157 |
| 李晟, 王大军, 肖治术, 等. 2014. 红外相机技术在我国野生动物研究与保护中的应用与前景[J]. 生物多样性, 22(6): 685–695. |
| 刘选珍, 李洪文, 毛杰. 2013. 蓝鹇和白鹇繁殖与非繁殖期间的日常行为比较[J]. 经济动物学报, 17(1): 27–31. |
| 卢汰春. 1991. 中国珍稀濒危野生鸡类[M]. 福州: 福建科学技术出版社. |
| 鲁庆彬, 游卫云, 赵昌杰, 等. 2011. 旅游干扰对青山湖风景区植物多样性的影响[J]. 应用生态学报, 22(2): 295–302. |
| 吕建中. 2009. 重修西天目山志[M]. 北京: 方志出版社. |
| 马建章. 1990. 野生动物管理学[M]. 哈尔滨: 东北林业大学出版社. |
| 孟秀祥, 潘世秀, 栾晓峰, 等. 2010. 兴隆山自然保护区马麝春季生境选择[J]. 生态学报, 30(20): 5509–5517. |
| 邵晨, 胡一中. 2005. 白鹇的夜栖息地选择及夜栖地行为[J]. 浙江林学院学报, 22(5): 562–565. DOI:10.3969/j.issn.2095-0756.2005.05.018 |
| 石建斌, 郑光美. 1997. 白颈长尾雉栖息地的季节变化[J]. 动物学研究, 18(3): 275–283. |
| 谭耀匡, 吴至康. 1981. 贵州白鹇的一新亚种——白鹇榕江亚种[J]. 动物学研究, 2(4): 301–306. |
| 吴名川. 1988. 白鹇鸡的生态及其在广西的分布[J]. 广西林业(2): 19–20. |
| 熊志斌, 谭成江, 余登利, 等. 2002. 贵州茂兰自然保护区白鹇种群数量与保护对策[J]. 四川动物, 21(4): 244–247. DOI:10.3969/j.issn.1000-7083.2002.04.012 |
| 熊志斌, 余登利, 谭成江, 等. 2003. 茂兰自然保护区白鹇种群数量与栖息地保护[J]. 贵州大学学报, 20(2): 200–204. DOI:10.3969/j.issn.1000-5269.2003.02.021 |
| 尹祚华. 1995. 白鹇饲养繁殖行为的观察[J]. 动物学杂志, 30(1): 50–51. |
| 张娟茹, 谢春雨, 倪丽. 2010. 半散养状态下白鹇的巢址选择性[J]. 野生动物, 31(4): 182–184. DOI:10.3969/j.issn.1000-0127.2010.04.005 |
| 张履冰, 崔绍朋, 黄元骏, 等. 2014. 红外相机技术在我国野生动物监测中的应用:问题与限制[J]. 生物多样性, 22(6): 696–703. |
| 赵玉泽, 曹婉露, 余进, 等. 2016. 基于红外相机技术的白冠长尾雉集群行为研究[J]. 生态学报, 36(9): 2513–2520. |
| 赵正阶. 2001. 中国鸟类志上卷非雀形目[M]. 长春: 吉林科学技术出版社. |
| 郑家文, 丁平, 徐肖江, 等. 2006. 白鹇种群分布与栖息地斑块特征的关系[J]. 应用生态学报, 17(5): 951–953. DOI:10.3321/j.issn:1001-9332.2006.05.040 |
| 郑作新. 1987. 中国鸟类区系纲要[M]. 北京: 科学出版社. |
| Anoop KR, Hussain SA. 2004. Factors affecting habitat selection by smooth-coated otters (Lutra perspicillata) in Kerala, India[J]. Journal of Zoology (London), 263: 417–423. DOI:10.1017/S0952836904005461 |
| Beest FMV, Mysterud A, Loe LE, et al. 2010. Forage quantity, quality and depletion as scale-dependent mechanisms driving habitat selection of a large browser herbivore[J]. Journal of Animal Ecology, 79(4): 910–922. |
| Delacour J. 1977. The pheasants of the world 2nd ed[M]. Hindhead: Saiga Publishing Co. LTD. |
| Koehler KJ, Larntz K. 1980. An empirical investigation of goodness-of-fit statistics for sparse multinomials[J]. Journal of the American Statistical Association, 75: 336–344. DOI:10.1080/01621459.1980.10477473 |
| Palmer JW. 1989. Canopy manipulation for optimum utilization of light[M]//Wright CJ. Manipulation of fruiting. London: Butterworths. |
| Potschin M, Haines-Young R. 2013. Landscapes, sustainability and the place-based analysis of ecosystem services[J]. Landscape Ecology, 28(6): 1053–1065. DOI:10.1007/s10980-012-9756-x |
| Rosenzweig ML. 1985. Some theoretical aspects of habitat selection[M]//Cody ML. Habitat selection in birds. New York: Academic Press: 135-140. |
| Schipper J. 2007. Camera-trap avoidance by Kinkajous Potos flavus:rethinking the "non-invasive" paradigm[J]. Small Carnivore Conservation, 36: 38–41. |
| Skarin A, Danell O, Bergstrom R, et al. 2004. Insect avoidance may override human disturbances in reindeer habitat selection[J]. Rangifer, 24(2): 95–103. DOI:10.7557/2.24.2.306 |
| Turner M, Donato D, Romme W. 2013. Consequences of spatial heterogeneity for ecosystem services in changing forest landscapes:priorities for future research[J]. Landscape Ecology, 28(6): 1081–1097. DOI:10.1007/s10980-012-9741-4 |
| Wagenmakers PS, Tazelaar M. 1999. Modelling light interception on the basis of sunfleck measurements[J]. Acta Horticulturae, 499: 297–301. |
| Wegge P, Pokheral CP, Jnawali SR. 2004. Effects of trapping effort and trap shyness on estimates of tiger abundance from camera trap studies[J]. Animal Conservation, 7(3): 251–256. DOI:10.1017/S1367943004001441 |