 2019, Vol. 38
2019, Vol. 38扩展功能
文章信息
- 赵栋, 杨创明, 和梅香, 陈俪心, 何兴成, 冉江洪
- ZHAO Dong, YANG Chuangming, HE Meixiang, CHEN Lixin, HE Xingcheng, RAN Jianghong
- 贡嘎山国家级自然保护区兔狲的活动节律与适宜栖息地预测
- Habitat Suitability Assessment and Daily Activity Patterns of Otocolobus manul in the Gongga Mountain National Nature Reserve
- 四川动物, 2019, 38(3): 320-327
- Sichuan Journal of Zoology, 2019, 38(3): 320-327
- 10.11984/j.issn.1000-7083.20180410
-
文章历史
- 收稿日期: 2018-12-28
- 接受日期: 2019-04-08






2. 四川贡嘎山国家级自然保护区管理局, 四川康定 626000
2. Gongga Mountain National Nature Reserve Administration Bureau, Kangding, Sichuan Province 626000, China
兔狲Otocolobus manul隶属于哺乳纲Mammalia食肉目Carnivora猫科Felidae兔狲属Otocolobus,国家Ⅱ级重点保护野生动物,为世界自然保护联盟(IUCN)濒危物种红色名录近危(NT)物种,被列入濒危野生动植物国际贸易公约(CITES)附录Ⅱ(Clark et al., 2006;Ross,2009),分布在亚洲中部向东至西伯利亚海拔4 000 m左右的荒漠、沙漠、草原和戈壁(Ross et al., 2015),全球有50%以上的兔狲分布在中国,主要分布于西部和北部(Fox & Dorji,2007;Smith & Xie,2008)。在蒙古国和中国,兔狲在野外面临的最大威胁是偷猎(Nowell & Jackson,1996;Murdoch et al., 2006)。现有兔狲的研究主要集中于蒙古国的Daurskij生物圈保护区和俄罗斯境内的阿尔泰山脉一带,包括食性(Ross,2009)、栖息地(Mallon,2002;Ross et al., 2015)、生态和行为(Heptner et al., 1992;Ross,2009)等方面。研究表明兔狲是独居的夜行性动物,主要在晨昏活动,捕食达乌尔鼠兔Ochotona dauurica和小型啮齿动物(Ross,2009),分布于高山草甸草原、低山丘陵、荒漠和半荒漠区、高寒草甸等寒冷、贫瘠、干旱少雨、温差极大的环境(Murdoch et al., 2006)。国内关于兔狲的研究极少,仅见消化系统、生殖器官和毛绒纤维结构等的基础研究(朱安国, 1999, 2002;李维红,吴建平,2011),尚无该物种野外生态与生物学的研究报道。
栖息地是动物的栖息环境或生态环境,为某一动物个体、种群或群落提供生活所必需的空间场所(Momsen et al., 1992)。栖息地选择是野生动物管理研究的重要内容,动物在自然环境中,会偏好利于自身生存和繁殖的生境,使其广义适合度达到最大(陈良,2007)。活动节律是动物行为学研究的重要方面,直接与动物的代谢和能量收支相关,但随环境和时间的推移不断调整(Halles & Sthenseth,2000)。动物的日活动节律和活动时间分配不仅受环境因素(如光照、温度)和食物资源时空分布的影响,还受到种群因素(如大小和组成)和社会因素(如社会等级)的影响(周岐海等,2012)。
四川贡嘎山国家级自然保护区地理位置独特,与西北高原地区的自然环境差异较大。2016年6月—2017年5月,采用样线法和红外相机陷阱调查法在保护区调查和研究兔狲的活动海拔、活动节律,并结合最大熵(MaxEnt)模型进行栖息地分布预测等,以期丰富其基础生物学、生态学资料,为其保护提供科学参考。
1 研究区域概况四川省贡嘎山国家级自然保护区地处青藏高原的东南缘(101°29′~102°12′E,29°01′~30°05′N),总面积4 091.4 km2,区内山体高大,谷岭高差悬殊,主峰海拔7 556 m,是青藏高原东部的最高峰和东亚地区的第一高峰,由主峰山顶东至大渡河谷地,直线距离仅30 km,谷岭高差超6 000 m,巨大的海拔高差导致生物气候随海拔增加而变化,形成了区内复杂多样的植被垂直带谱,且东西坡差异明显。东坡植被类型多样,垂直带谱完整而复杂,海拔1 000~ 2 200 m为常绿阔叶林带(含河谷灌丛)、2 200~2 500 m为山地针阔叶混交林带、2 500~3 600 m为亚高山针叶林带、3 600~4 600 m为高山灌丛草甸带、4 600~5 100 m为高山流石滩稀疏植被带、5 100 m以上为永久冰雪带。西坡植被类型及垂直带谱均较简单,包括2 800~4 000 m亚高山针叶林带、4 000~ 4 600 m高山灌丛草甸带、4 600~5 200 m高山流石滩稀疏植被带以及5 200 m以上的永久冰雪带(沈泽昊,2004)。
2 研究方法 2.1 野外调查方法 2.1.1 样线调查以最短的距离尽可能地穿过保护区内兔狲的生境,使样线能够全面、有效地收集信息。在海拔3 500~5 100 m内按每一个5 km×5 km网格布设2条样线,每条样线长3~4 km,共52条样线(图 1)。采集样线上发现的疑似兔狲粪便,装入95%乙醇中,-20 ℃保存,同时记录伴生动物。

|
| 图 1 研究区域及样线分布 Fig. 1 Distribution of study area and transects |
| |
根据Murdoch等(2006)的研究,结合保护区地形地貌及本底资料,按5 km×5 km网格布设红外相机,每个网格放置3~4台,共设置177台,调查面积为650 km2。红外相机的布设及设置参照汪国海等(2015)的方法。
2.2 数据分析方法 2.2.1 兔狲粪便识别粪便样品提取DNA,选择线粒体cyt b基因的特异性引物进行物种鉴定和遗传结构分析,引物对为CAT-CytB-F/R。将PCR产物测序并导入GenBank中的BLAST进行比对,与数据库中的已知序列同源性在97%以上时,确定为相同物种(张于光等,2009)。
2.2.2 活动节律分析方法对收集的红外相机照片进行筛选,保留能够准确判断物种的照片和视频,记录所拍照片和视频的时间、数量等数据。由于动物在相机监测视野内进行觅食、行走或排泄等行为时会触发红外相机连续拍摄多张照片或视频,因此把同一相机位点拍摄的包含相同物种个体照片之间的时间间隔在30 min之内的视为1次独立捕捉,30 min内的多张照片视为1次独立照片,只记录1个活动时间数据(O'Brien et al., 2003)。
日活动节律:基于兔狲的红外相机独立捕捉数据,利用核密度估计法建立兔狲日活动节律的函数模型,公式如下:
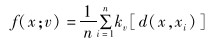
|
其中,kv为von Mises分布的概率密度函数,d(x,xi)为任意一点x与样本量n之间的角度距离(Ridout & Linkie,2009)。
日活动差异指数(α)是衡量兔狲在一天内的时间分配均匀程度。昼行性指数(β)用于衡量兔狲日活动主要在昼间还是在夜间。计算公式如下(汪国海等,2015):
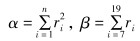
|
n为时间点,以1 h为时间间隔把一天分成连续的24个单位,其中i是任一时间点;ti为第i个时间点的时间值,00: 00—01: 00记为时间点0,01: 01—02: 00记为时间点1,02: 01—03: 00记为时间点2,依次类推;ri为ti时刻的相对活动强度,定义为ti时刻拍摄的兔狲数量(fi)占各时刻拍摄到兔狲总数(F)的比例,即ri=fi/F。若α≈0.042(即α=1/24时),则表示兔狲活动强度在一天各时间段内分配完全均匀;α越小,其在一天内的活动时间分配越均匀;α越大,其日活动时间分配越不均匀。
根据汪国海等(2015)的研究,动物是昼行性还是夜行性主要依据昼行性指数判断。β≈0.542 9(13/24),表明其日活动昼夜差别不大;β<0.542,表明其日活动主要在夜间发生;β>0.542,表明其日活动主要在昼间发生。
2.2.3 栖息地预测采用MaxEnt模型进行栖息地预测(余翔等,2017):将兔狲分布点位导入ArcGIS中,按1 km×1 km布设栅格,每个栅格只留最接近中心的1个分布点,最终选取38个兔狲分布点,将所有分布点的经纬度数据在Excel表格中汇总,保存CSV格式用于MaxEnt模型计算,用于建立适宜栖息地模型。
选择3类环境变量数据:(1)生物学变量,包括2年的、5年的增强型植被指数(EVI),该指数能反映地表植被的分布状况(崔绍朋等,2018);(2)生物气候变量,共19个,从WorldClim数据库中下载(Hijmans et al., 2005);(3)地形变量,包括海拔、坡度、坡向(余翔等,2017),对研究区域内的环境变量进行主成分分析,结合这些环境变量之间的相关性分析结果,剔除共线性高的变量(利用ArcGIS多元分析进行波段集统计,将Pearson相关系数绝对值大于0.7的剔除一个),综合考虑对模型贡献率高且彼此间相关性不高的环境变量(乔麦菊等,2017)。最终选择9个环境变量预测兔狲的潜在分布范围,包括平均日较差、气温年较差、EVI 2年最大值均值、海拔、坡度、坡向、到河流距离、到林地距离、植被类型(表 1)。
| 编码Code | 描述Description | 来源Source |
| vegetation1 | 植被类型 | Landsat 8 OLI_TIRS |
| slp | 坡度 | USGS's Hydro-1K dataset |
| river | 到河流距离 | Google Earth |
| forest | 到林地距离 | Landsat 8 OLI_TIRS |
| evi-2y-maxi | EVI 2年 最大值均值 |
Chinese Natural Resources Database |
| dem | 海拔 | USGS's Hydro-1K dataset |
| c0-bio-30s-07 | 气温年较差 | WorldClim database Version 1.4 |
| c0-bio-30s-02 | 平均日较差 | WorldClim database Version 1.4 |
| asp | 坡向 | USGS's Hydro-1K dataset |
将兔狲分布点和相应的环境变量导入MaxEnt模型(Phillips et al., 2006),随机选取75%的数据作为训练数据用于模型构建,其余25%作为检验数据,其余设置为默认(余翔等,2017),为保证模型结果的稳定性,进行10次自举法(bootstrap)重复,模型构建参照乔麦菊等(2017)和余翔等(2017)等。选取最大约登指数为阈值(周志美,2011),用ArcGIS中的重分类功能将此阈值以上的划分为兔狲适宜栖息地。
3 结果 3.1 海拔分布调查获得47个兔狲的活动位点(33个来自红外相机位点,14个通过粪便DNA barcoding鉴定),结果显示,保护区兔狲活动海拔为3 780~4 956 m,4 500 m以上的分布点最多,共有41个(87.2%),4 000~4 500 m有5个,低于4 000 m的只有1个。
3.2 活动节律33台红外相机共拍摄到50张独立照片,对所有独立照片进行核密度函数建模,计算得出α=0.071 2(>0.042),β=0.34(<0.542)。这表明兔狲日活动时间分配不均匀,活动高峰为05: 00—07: 00和20: 00—22: 00 (图 2)。

|
| 图 2 兔狲日活动节律 Fig. 2 Daily activity patterns of Otocolobus manul in the Gongga Mountain National Nature Reserve 横坐标上方的短竖线表示每一独立捕捉事件发生的时间 The short vertical lines above the abscissa indicate the time of individual independent captures |
| |
MaxEnt结果显示,经过10次重复交叉检验的平均训练集值为0.978,说明模型的拟合程度很高,预测结果较准确。其中,EVI 2年最大值均值、平均日较差、气温年较差、海拔和植被类型对兔狲分布的影响较大(图 3)。

主要环境因素响应曲线显示(图 4),兔狲在保护区偏好的植被类型为高山流石滩,偏好在海拔4 000 m以上的温差较大的地区活动。

|
| 图 4 环境因子对模型预测的平均响应曲线 Fig. 4 Response curves of environmental factors on model prediction 红色:10次重复的平均响应曲线,蓝色:平均值±标准差 Red: the mean response curves of 10 replicate MaxEnt runs, blue: mean±standard deviation |
| |
当训练数据的特异度和敏感度之和达到最大时,阈值为0.224 7,以此为分界点对保护区兔狲栖息地适宜度结果进行重分类,兔狲在保护区的适宜栖息地面积约187 km2,占保护区总面积的4.56%(图 5)。

|
| 图 5 贡嘎山国家级自然保护区兔狲适宜栖息地分布 Fig. 5 Suitable habitats of Otocolobus manul in the Gongga Mountain National Nature Reserve |
| |
全球各个地区都正在发生着不同程度的生物多样性危机(蒋志刚,马克平,2014),研究受胁物种适宜生境现状,了解影响其地理分布的环境因素,对开展保护工作有重要的指导作用。实际上很多物种的地理分布数据缺乏,这不利于研究和解决其生态学问题,特别是濒危物种。随着栖息地环境的退化和破碎化,兔狲的分布分散、种群密度低,种群分布趋势未知(Damdinsuren et al., 2008;Barashkova & Smelansky,2011;Ross et al., 2012)。
栖息地是动物生活和繁殖的场所,能够为动物提供充足的食物资源、适宜的繁殖地点、躲避天敌和不良气候的隐蔽和保护等,是保证其生存和繁衍的基本条件(张正旺,郑光美,1999)。MaxEnt结果显示,EVI 2年最大值、平均日较差、气温年较差、海拔和植被类型是影响兔狲生境选择的主要环境因子。兔狲活动海拔为3 780~4 956 m,主要分布于4 500 m以上,这与之前的报道一致(Fox & Dorji,2007;Ross et al., 2015)。四川贡嘎山国家级自然保护区高海拔地区人迹罕至,分布有裸岩坡和沟壑,为兔狲窝居和穴居创造了良好的条件,也有利于其躲避天敌。David(1994)、Sunquist和Sunquist(2002)的研究显示,兔狲分布于高山草甸草原、灌丛草甸、低山丘陵、荒漠与半荒漠地区,本研究显示兔狲主要分布于高山流石滩地带。造成这种差异的原因可能是物种生境选择与不同研究区的植被组成和异质性相关,某一地区不同植被的相对多度与组成或镶嵌格局可能会影响物种对不同植被的选择(Lorenzón et al., 2016)。保护区地形地貌复杂,海拔3 600~4 600 m为高山灌丛草甸带,海拔4 600 m以上为高山流石滩植被,这些植被中丰富的鼠兔等啮齿动物为兔狲提供了充足的食物。因此,建议从更大尺度(如家域尺度)深入研究兔狲对植被类型的选择,以及异质性对其选择的影响。
李明富等(2011)研究发现动物在野外被相机拍摄到的概率与其活动强度呈正比,相对活动强度指数越大,说明该动物在此时间段内活动强度越大。动物的日活动节律受自身代谢和能量需求、社群关系、种间关系,以及季节、天气状况等诸多因素的影响,是物种对周围环境适应的结果(金建丽等,2003)。Heptner等(1992)研究发现兔狲是夜行性动物,蒙古国境内的兔狲主要在晨昏活动,有时也在夜间活动(Murdoch et al., 2006),但没有明确报道其活动高峰及活动原因。保护区兔狲的活动高峰为05: 00—07: 00和20: 00—22: 00,推测这可能与其捕食相关。贡嘎山地区和蒙古国的气候和生物资源差异较大,贡嘎山高山灌丛草甸有高原鼠兔Ochotona curzoniae等的分布,要确定兔狲的食物组成以及与活动节律的关系还需进一步研究。
在蒙古国,偷猎、猎物种群被破坏、栖息地片段化和退化对兔狲的生存造成了负面影响(罗述金,2014)。本研究结果显示,保护区兔狲适宜栖息地被分成南北两部分,但从其分布区域来看,其栖息地基本是连续的。所以,保护区不仅应关注保护区内的状况,也需要关注与保护区相连接的保护区外的区域,以保障高山生态系统中的高寒动物个体交流,保护整个高山高寒生态系统的健康。
致谢: 感谢四川贡嘎山国家级自然保护区管理局提供的样品和野外数据,感谢李波、朱博伟、付焱文对本文的修改。| 陈良. 2007.黑麂栖息地特征和栖息地选择的季节变化[D].金华: 浙江师范大学. http://cdmd.cnki.com.cn/Article/CDMD-10345-2007184814.htm |
| 崔绍朋, 罗晓, 李春旺, 等. 2018. 基于MaxEnt模型预测白唇鹿的潜在分布区[J]. 生物多样性, 26(2): 171–176. |
| 蒋志刚, 马克平. 2014. 保护生物学原理[M]. 北京: 科学出版社. |
| 金建丽, 张春美, 杨春文. 2003. 棕背昼夜活动节律的研究[J]. 应用生态学报, 14(6): 1019–1022. DOI:10.3321/j.issn:1001-9332.2003.06.041 |
| 李明富, 李晟, 王大军, 等. 2011. 四川唐家河自然保护区扭角羚冬春季日活动模式研究[J]. 四川动物, 30(6): 850–855. DOI:10.3969/j.issn.1000-7083.2011.06.031 |
| 李维红, 吴建平. 2011. 旱獭、麝鼠、兔狲、青鼬、石貂毛绒纤维超微结构比较[J]. 畜牧兽医学报, 42(7): 994–999. |
| 罗述金. 2014. 中国猫科动物[M]. 北京: 中国林业出版社. |
| 乔麦菊, 唐卓, 施小刚, 等. 2017. 基于MaxEnt模型的卧龙国家级自然保护区雪豹(Panthera uncia)适宜栖息地预测[J]. 四川林业科技, 38(6): 1–4. |
| 沈泽昊, 刘增力, 伍杰. 2004. 贡嘎山东坡植物区系的垂直分布格局[J]. 生物多样性, 12(1): 89–98. DOI:10.3321/j.issn:1005-0094.2004.01.011 |
| 汪国海, 施泽攀, 李生强, 等. 2015. 基于红外相机技术的赤麂活动模式分析[J]. 广西师范大学学报(自然科学版), 33(3): 117–122. |
| 余翔, 陈俊橙, 王彬, 等. 2017. 四川小寨子沟国家级自然保护区绿尾虹雉种群密度调查与栖息地评价[J]. 四川动物, 36(4): 361–367. |
| 张于光, 何丽, 朵海瑞, 等. 2009. 基于粪便DNA的青海雪豹种群遗传结构初步研究[J]. 兽类学报, 29(3): 310–315. DOI:10.3969/j.issn.1000-1050.2009.03.011 |
| 张正旺, 郑光美. 1999. 鸟类栖息地选择研究进展[M]. 北京: 中国林业出版社. |
| 周岐海, 黄恒连, 唐小平, 等. 2012. 白头叶猴全雄群的日活动节律和活动时间分配[J]. 广西师范大学学报(自然科学版), 30(3): 282–287. DOI:10.3969/j.issn.1001-6600.2012.03.045 |
| 朱安国. 1999. 雄性兔狲生殖器官特征[J]. 上海畜牧兽医通讯, 5: 20–21. |
| 朱安国. 2002. 兔狲消化腺特征[J]. 青海大学学报, 18(1): 7–9. |
| Barashkova A, Smelansky I. 2011. Pallas's cat in the Altai Republic, Russia[J]. Cat News, 54: 1–7. |
| Clark EL, Munkhbat J, Dulamtseren S, et al. 2006. Summary conservation action plans for Mongolian mammals[M]. London: Zoological Society of London. |
| Damdinsuren B, Herrick JE, Pyke DA, et al. 2008. Is rangeland health relevant to Mongolia?[J]. Rangelands, 30: 25–29. |
| David B. 1994. Seasonal differences in habitat selection by black grouse in the northern Pennines[J]. IBIS, 136: 39–43. |
| Fox JL, Dorji T. 2007. High elevation record for occurrence of the manul or Pallas's cat on the northwestern Tibetan Plateau, China[J]. Cat News, 46: 35. |
| Halles S, Sthenseth NC. 2000. Activity patterns in small mammals:an ecological approach[M]. New York: Springer. |
| Heptner VG, Sludskii AA, Bannikov AG. 1992. Mammals of the Soviet Union. Volume Ⅱ, Part 2:Carnivora (hyaenas and cats)[M]. Washington DC: Smithsonian Institution and the National Science Foundation. |
| Hijmans RJ, Cameron SE, Parra JL, et al. 2005. Very high resolution interpolated climate surfaces for global land areas[J]. International Journal of Climatology, 25: 1965–1978. DOI:10.1002/(ISSN)1097-0088 |
| Lorenzón RE, Beltzer AH, Olguin PF, et al. 2016. Habitat heterogeneity drives bird species richness, nestedness and habitat selection by individual species in fluvial wetlands of the Paraná River, Argentina[J]. Austral Ecology, 41(7): 829–841. DOI:10.1111/aec.12375 |
| Mallon D. 2002. Manul sighting in Qinghai[J]. Cat News, 36: 18. |
| Momsen ML, Marcot BG, Mannan RW. 1992. Wildlife habitat relationships:concepts and application[M]. Wisconsin: The University of Wisconsin Press. |
| Murdoch JD, Munkhzul T, Reading RP. 2006. Pallas' cat ecology and conservation in the semidesert steppes of Mongolia[J]. Cat News, 45: 18–19. |
| Nowell K, Jackson P. 1996. Wild cats-status survey and conservation action plan[M]. Gland, Switzerland and Cambridge, UK: IUCN/SSC Cat Specialist Group. |
| O'Brien TG, Kinnaird MF, Wibisono HT. 2003. Crouching tigers, hidden prey:Sumatran tiger and prey populations in a tropical forest landscape[J]. Animal Conservation Forum, 6(2): 131–139. DOI:10.1017/S1367943003003172 |
| Phillips SJ, Anderson RP, Schapire RE. 2006. Maximum entropy modelling of species geographic distributions[J]. Ecological Modelling, 190: 231–259. DOI:10.1016/j.ecolmodel.2005.03.026 |
| Ridout MS, Linkie M. 2009. Estimating overlap of daily activity patterns from camera trap data[J]. Journal of Agricultural, Biological, and Environmental Statistics, 14(3): 322–337. DOI:10.1198/jabes.2009.08038 |
| Ross S. 2009. Providing an ecological basis for the conservation of the Pallas's cat Otocolobus manul[D]. Bristol, UK: University of Bristol. |
| Ross S, Barashkova Y, Farhadinia, et al. 2015. Otocolobus manul[J].[2018-12-20]. The IUCN Red List of Threatened Species 2015: e.T 15640A50657610. |
| Ross S, Munktsog B, Harris S. 2012. Determinants of mesocarnivore range use:relative effects of prey and habitat properties on Pallas's cat home-range size[J]. Journal of Mammalogy, 93(5): 1292–1300. DOI:10.1644/11-MAMM-A-060.1 |
| Smith AT, Xie Y. 2008. A guide to the mammals of China[M]. New Jersey: Princeton University Press. |
| Sunquist M, Sunquist F. 2002. Wild cats of the world[J]. Chicago:University of Chicago Press. |