 2018, Vol. 37
2018, Vol. 37扩展功能
文章信息
- 陈祯, 周成理, 陈晓鸣, 石雷
- CHEN Zhen, ZHOU Chengli, CHEN Xiaoming, SHI Lei
- 温度和光周期对虎斑蝶幼期生长发育的影响
- Effects of Temperature and Photoperiod on the Development of Immature Stages of Danaus genutia
- 四川动物, 2018, 37(4): 420-425
- Sichuan Journal of Zoology, 2018, 37(4): 420-425
- 10.11984/j.issn.1000-7083.20170379
-
文章历史
- 收稿日期: 2017-11-28
- 接受日期: 2018-04-20






2. 中国林业科学研究院资源昆虫研究所, 昆明 650224
2. Research Institute of Resource Insects, Chinese Academy of Forestry, Kunming 650224, China
虎斑蝶Danaus genutia隶属蛱蝶科Nymphalidae斑蝶亚科Danainae斑蝶属Danaus,又名粗脉棕斑蝶、黑脉桦斑蝶、拟阿檀蝶、虎纹青斑蝶和青红条红斑蝶等。国内分布于河南、四川、云南、西藏、江西、浙江、福建、广东、广西、台湾和海南等地,国外分布于中南半岛、西太平洋诸岛和澳大利亚(周尧,1998)。成虫中大体型,访花,翅膀正面色彩鲜艳,飞行缓慢,姿态优美,是生态观赏、喜庆放飞和工艺制作的优良蝶种,无危害经济和园林植物记录。该种蝴蝶在国内蝴蝶观赏园及庆典蝴蝶放飞中被广泛利用,但其主要生物学、生理生态学特征等鲜见报道。在国内,陈晓鸣等(2008)首次对虎斑蝶的主要寄主、幼期形态特征及成虫和幼虫习性进行了初步记述;陈祯等(2017)对云南元江虎斑蝶种群的全虫期形态特征、幼期发育历期、在云南昆明温室中的代生活史和生殖力、寄主范围、成虫及幼虫的行为习性进行了详细报道。在国外,Meyer(1995)简略描述了澳大利亚虎斑蝶D. genutia alexis的1龄、5龄幼虫及蛹的形态,幼虫期和蛹期在自然条件下的发育历期;Bhuyan等(1999)研究了虎斑蝶成虫对4种蜜源植物的选择性;Atluri等(2010)报道了印度Visakhapatnam地区虎斑蝶的幼期形态、幼期发育历期及幼虫取食寄主植物Pergularia daemia后的生长发育情况;此外,学者也对虎斑蝶的寄主植物进行了报道(Meyer,1995;Igarashi & Fukuda,2000;Robinson et al., 2001;Atluri et al., 2010)。
近10多年来,野生虎斑蝶因美丽的颜色斑纹及轻缓的飞行方式被大量采集,用于商业放飞和工艺品制作,野生种群数量急剧下降,野生资源在数量和质量上都无法满足国内迅猛发展的市场需要,开展人工繁育势在必行。为此,本文作者近年来对云南省元江县虎斑蝶的主要生物学、生理生态学特征进行了较为系统的持续研究,以期筛选出优良寄主植物,找出最佳幼期发育条件和成虫繁殖条件,研发出高效的规模化人工繁育技术。本文报道温度和光周期对虎斑蝶幼期生长发育的影响。
1 材料与方法 1.1 实验材料虎斑蝶:成虫种源采自云南省元江县野外,将采回的野生雌雄成虫置于云南省昆明市盘龙区中国林业科学研究院资源昆虫研究所蝴蝶试验大棚内放养,建立实验种群。采集新产的卵供实验用。
饲养幼虫的寄主植物:青羊参Cynanchum otophyllum(萝藦科Asclepiadaceae)。
成虫补充营养:10%蜂糖水。
1.2 实验方法 1.2.1 温度和光周期对虎斑蝶幼期生长发育的影响在人工气候箱(型号:SPX-400;上海博讯医疗设备厂)内设置2个光照处理[长光照(LD=15: 9)和短光照(LD=9: 15),依据Goehring和Oberhauser(2002)对虎斑蝶近缘种君主斑蝶Danaus plexippus相关研究中的光周期,并结合元江冬、夏季最短和最长光照时数考虑]。长光照下设7个温度梯度:17.5 ℃、20.0 ℃、22.5 ℃、25.0 ℃、27.5 ℃、30.0 ℃和35.0 ℃;短光照下设5个温度梯度:17.5 ℃、20.0 ℃、25.0 ℃、30.0 ℃和35.0 ℃。以上处理组的湿度均恒定为70%。
由于卵的孵化率和初龄幼虫成活率存在差异,各处理中使用不同初始卵量以保证每个处理中至少有60头幼虫存活至2龄初。故每个处理自80~150粒新产卵开始,幼虫孵化后以保鲜枝叶单虫饲养,直至化蛹、羽化。1~3龄期每2日更换1次食料,此后每日更换食料。每日记录卵孵化数、幼虫蜕皮数、化蛹数、羽化数,计算各虫态发育历期。
1.2.2 幼期各虫态发育起点温度和有效积温测定长光照下幼期各虫态发育起点温度(C)和有效积温(K),设置了17.5 ℃、20.0 ℃、22.5 ℃、25.0 ℃、27.5 ℃、30.0 ℃ 6个恒定温度梯度,根据公式T=C+KV,式中,T为温度,V为各温度下的发育速率,采用“直线回归法”计算虎斑蝶各个虫态的发育起点温度与有效积温(张孝羲,2011)。
1.3 数据分析利用SPSS 19.0对数据进行方差分析,平均数进行Duncan’s检验,显著性水平α=0.05;多元相关分析采用偏相关分析。
2 结果 2.1 温度和光周期对卵发育历期的影响不同温度条件下,长光照组虎斑蝶卵的发育历期为2~11 d,短光照组为2~10 d(表 1)。在相同光照和设定温度范围内,随着温度上升,卵的发育加快,发育历期缩短。无论是在长光照还是短光照条件下,同一温度条件下实验卵的发育均较为整齐。在长、短光照处理中,当温度设定为35 ℃时,卵均不能孵化。偏相关分析表明,在固定温度条件下,光照与卵的发育历期呈不显著正相关(R2=0.376,P=0.406);在固定光照条件下,温度与卵的发育历期呈极显著负相关(R2=-0.950,P=0.005)。
| 光周期 Photoperiod |
温度 Temperature/℃ |
发育历期 Developmental duration/d |
样本量 Sample size/粒 |
| 长光照 (15:9) |
17.5 | 9.29±1.12a | 113 |
| 20.0 | 4.90±0.90b | 120 | |
| 22.5 | 4.91±0.29b | 93 | |
| 25.0 | 3.97±0.17c | 100 | |
| 27.5 | 3.54±0.83d | 146 | |
| 30.0 | 3.20±0.41c | 136 | |
| 35.0 | 0 | 95 | |
| 短光照 (9:15) |
17.5 | 9.04±0.87a | 88 |
| 20.0 | 8.20±1.15b | 85 | |
| 25.0 | 4.11±0.50c | 108 | |
| 30.0 | 3.04±0.55d | 120 | |
| 35.0 | 0 | 92 | |
| 注:同列数据(平均数±标准误)后标有不同字母表示差异有统计学意义;下同 Notes:The different letters in the same column indicate there is a significant difference;the same below | |||
不同温度条件下,长光照组虎斑蝶幼虫的发育历期为9~50 d,短光照组为8~40 d(表 2)。在相同光照和设定温度范围内,随着温度上升,幼虫的生长发育加快,发育历期缩短。无论是在长光照还是短光照条件下,同一温度下幼虫不同龄期的发育历期为:5龄>1龄>4龄>2龄和3龄。偏相关分析表明,在固定温度条件下,光照与幼虫的发育历期呈不显著负相关(R2=-0.110,P=0.814);在固定光照条件下,温度与幼虫的发育历期呈显著负相关(R2=-0.847,P=0.016)。
| 处理 Treatments | 发育历期 Developmental duration/d | ||||||
| 光周期 Photoperiod |
温度 Temperature/℃ |
1龄幼虫 1st instar |
2龄幼虫 2nd instar |
3龄幼虫 3rd instar |
4龄幼虫 4th instar |
5龄幼虫 5th instar |
幼虫全期 1st-5th instar |
| 长光照 (15:9) |
17.5 | 7.29±1.27a (72) | 6.17±0.96a (49) | 6.71±1.40a (38) | 7.17±1.49a (37) | 10.71±1.73a (30) | 38.04±3.65a (30) |
| 20.0 | 3.83±1.04b (69) | 2.90±0.67b (41) | 2.83±0.60b (35) | 3.31±0.71b (33) | 5.03±0.72b (31) | 17.89±1.33b (31) | |
| 22.5 | 3.17±0.78c (52) | 2.70±0.63b (47) | 2.91±0.60b (39) | 3.00±0.52b (32) | 5.35±0.81b (30) | 17.13±1.50b (30) | |
| 25.0 | 1.91±0.38d (75) | 1.70±0.68c (66) | 1.58±0.61c (47) | 2.03±0.53c (42) | 3.60±1.15c (34) | 10.82±1.55c (34) | |
| 27.5 | 2.25±0.61d (76) | 1.58±0.50c (42) | 1.42±0.50c (27) | 2.00±0.72c (26) | 2.67±0.48d (23) | 9.92±0.88c (23) | |
| 30.0 | 2.33±0.62d (84) | 1.67±0.62c (50) | 1.33±0.62c (27) | 1.73±0.88c (18) | 3.13±0.35cd (15) | 10.20±1.57c (15) | |
| 短光照(9:15) | 17.5 | 5.81±1.06a (76) | 4.96±1.22a (54) | 5.04±1.22a (34) | 5.19±1.10a (30) | 9.08±0.99a (30) | 30.08±2.96a (30) |
| 20.0 | 5.07±0.70b (45) | 2.93±0.46b (27) | 2.67±0.49b (17) | 2.80±0.56b (17) | 5.07±0.57b (17) | 18.54±0.96b (17) | |
| 25.0 | 3.21±0.74c (71) | 1.89±0.63c (53) | 1.96±0.58c (31) | 2.32±0.67b (28) | 3.90±0.32c (28) | 13.29±1.08c (28) | |
| 30.0 | 2.29±0.62d (77) | 1.38±0.58d (43) | 1.38±0.58d (33) | 1.50±0.59c (22) | 3.00±0.20d (22) | 9.54±1.10d (22) | |
| 注 Note:括号内数字为样本量 Number in the bracket is the sample size | |||||||
不同温度条件下,长光照组虎斑蝶蛹的发育历期为6~28 d,短光照组为5~24 d(表 3)。在相同光照和设定温度范围内,随着温度上升,蛹的发育加快,发育历期缩短。方差分析表明,无论是在长光照还是短光照条件下,不同温度条件下蛹的发育历期差异均有高度统计学意义(P<0.01)。偏相关分析表明,在固定温度条件下,光照与蛹的发育历期呈不显著负相关(R2=-0.005,P=0.991);在固定光照条件下,温度与蛹的发育历期呈极显著负相关(R2=-0.911,P=0.004)。
| 光周期 Photoperiod |
温度 Temperature/℃ |
发育历期 Developmental duration/d |
样本量 Sample size |
| 长光照 (15:9) |
17.5 | 22.71±2.35a | 29 |
| 20.0 | 13.72±1.00b | 30 | |
| 22.5 | 12.00±0.80c | 29 | |
| 25.0 | 7.73±0.45d | 34 | |
| 27.5 | 7.46±0.59de | 23 | |
| 30.0 | 6.80±0.56e | 15 | |
| 短光照 (9:15) |
17.5 | 21.19±1.86a | 29 |
| 20.0 | 13.40±0.51b | 17 | |
| 25.0 | 9.64±0.56c | 28 | |
| 30.0 | 6.38±0.58d | 22 |
不同温度条件下,长光照组虎斑蝶从卵到成虫羽化整个幼期的发育历期为18~82 d,短光照组为17~70 d(表 4)。在相同光照和设定温度范围内,随着温度上升,幼期的生长发育加快,发育历期缩短。偏相关分析表明,在固定温度条件下,光照与幼期的发育历期呈不显著负相关(R2=-0.018,P=0.969);在固定光照条件下,温度与幼期的发育历期呈极显著负相关(R2=-0.885,P=0.008)。
| 光周期 Photoperiod |
温度 Temperature/℃ |
发育历期 Developmental duration/d |
样本量 Sample size |
| 长光照 (15:9) |
17.5 | 70.04±3.69a | 30 |
| 20.0 | 36.52±1.79b | 31 | |
| 22.5 | 34.04±1.64c | 30 | |
| 25.0 | 22.52±1.68d | 34 | |
| 27.5 | 20.92±1.50e | 23 | |
| 30.0 | 20.20±2.21e | 15 | |
| 短光照 (9:15) |
17.5 | 60.31±3.95a | 29 |
| 20.0 | 40.13±1.73b | 17 | |
| 25.0 | 27.04±1.43c | 28 | |
| 30.0 | 18.96±1.33d | 22 |
虎斑蝶完成整个幼期发育需要的有效积温为286.28 d·℃±33.62 d·℃。其中,卵的发育起点温度和有效积温最低,分别为11.10 ℃±1.56 ℃和55.15 d·℃±6.68 d·℃;其次是蛹,分别为13.46 ℃±1.01 ℃和98.95 d·℃±9.27 d·℃;幼虫的最高,分别为14.06 ℃±1.58 ℃和131.62 d·℃±20.49 d·℃,其有效积温约占幼期各虫态积温总和的45%。幼虫不同龄期的发育起点温度由大到小依次是:3龄>5龄>1龄>2龄>4龄;幼虫在不同龄期的有效积温由大到小依次是:5龄>1龄>4龄>2龄>3龄(表 5)。
| 虫期 Developmental stage |
发育起点温度 Developmental threshold temperature/℃ |
有效积温 Effective accumulated temperature/(d·℃) |
发育历期的预测式 Forecast formula of developmental duration |
| 卵 | 11.01±1.56 | 55.15±6.68 | 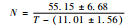 |
| 1龄幼虫 | 14.51±2.60 | 25.65±6.68 |  |
| 2龄幼虫 | 14.18±1.72 | 20.82±3.57 |  |
| 3龄幼虫 | 15.45±1.13 | 16.53±2.11 |  |
| 4龄幼虫 | 13.75±0.87 | 25.02±2.08 | 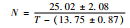 |
| 5龄幼虫 | 14.65±2.19 | 36.72±8.47 | 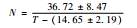 |
| 幼虫全期 | 14.06±1.58 | 131.62±20.49 |  |
| 蛹 | 13.46±1.01 | 98.95±9.27 |  |
| 卵~成虫羽化 | 13.38±1.27 | 286.28±33.62 |  |
本研究初步探讨了温度和光周期对虎斑蝶幼期虫态发育的影响,基本确定了其适宜温度范围,为该重要观赏蝶种的人工繁育提供了重要参数。结果表明,在17.5~30.0 ℃内,虎斑蝶卵、幼虫和蛹均能正常发育;随着温度上升,各虫态的生长发育加快,发育历期显著缩短;当温度为15 ℃和35 ℃时,卵均不能孵化;温度是影响幼期发育速率的主导因素。温度不仅影响发育速率,也影响幼虫成活率。初步观察表明,在17.5 ℃和30.0 ℃下,虎斑蝶幼虫的存活率较20.0~27.5 ℃下明显降低(陈祯,2010)。Goehring和Oberhauser(2002)认为,30 ℃以上的高温对君主斑蝶Danaus plexippus幼虫可能有亚致死作用,并大幅延长了其发育历期。吕龙石等(2004)对麝凤蝶Byasa alcinous的研究表明,30 ℃以上的温度不利于幼虫的生长与化蛹,34 ℃已经超出卵孵化与越冬蛹羽化的上限温度。
本研究中,光照与虎斑蝶幼期各虫态的发育历期相关性均不显著。但Goehring和Oberhauser(2002)发现,较短及渐短的光照都能加快君主斑蝶幼期发育,并诱发其成虫进入生殖休眠。这种差异可能是2个物种间的光周期反应性不同所致,也可能与本研究中光周期设置较少有关。在其他蝶类中,易传辉等(2007)发现凤蝶科Papilionidae美凤蝶Papilio memnon在12~15 h光照内,幼虫最长和最短发育历期相差9.2 d,25 ℃时相差达10.6 d,差异十分显著。与此类似,光周期对凤蝶科柑橘凤蝶Papilio xuthus幼期生长发育也有明显影响,不同光周期下幼虫发育历期、幼虫化蛹的发育历期都存在明显差异(易传辉等,2009)。而Yoshio和Ishii(2004)的结果表明,光周期对马来西亚沙巴美凤蝶幼虫发育历期的影响并不明显。这些差别可能是同一物种不同地理种群间的遗传差异所致。
本研究还计算了幼期各虫态的发育起点温度和有效积温。在虎斑蝶幼期虫态中,卵的发育起点温度最低(11.10 ℃±1.56 ℃),这在我国绝大多数地方的晚春至初秋都能满足。整个幼期发育需要的有效积温为286.28 d·℃±33.62 d·℃,据此初步估算在我国南方低海拔地区可年养殖5代以上,而在热带和部分南亚热河谷,年养殖更可高达10代以上。这种生物学特性赋予了虎斑蝶巨大的养殖利用潜力。在养殖生产上,建议将幼期养殖温度控制在20.0~27.5 ℃,同时可调整养殖温度以适时满足市场对活体成虫的需要。光照并非养殖生产中需要重点管理的条件,采用自然光照即可。
在蝴蝶的人工繁育中,温度是关键的技术指标之一,但相关研究并不多见,且主要集中在凤蝶科和蛱蝶科蛱蝶亚科种类中(易传辉等, 2007, 2009)。斑蝶亚科Nymphalinae的大多数种类色彩艳丽、飞行缓慢,是当前国内蝴蝶生态观赏园中的主流蝶种。这类蝴蝶年世代数多、繁殖力强,极具开发利用潜力。有关这个重要蝴蝶类群的生理生态研究国外已有一些报道(Malcolm et al., 1987;Goehring & Oberhauser,2002),国内这方面的研究工作还需要进一步加强。
致谢: 西南林业大学保护生物学学院曹永同学和蒋创同学参与了部分实验观察,昆明中林观赏昆虫科技开发有限公司赵灿繁协助采集实验虎斑蝶种源。在此一并致谢!| 陈晓鸣, 周成理, 史军义, 等. 2008. 中国观赏蝴蝶[M]. 北京: 中国林业出版社: 207-209. |
| 陈祯, 曹永, 周元清, 等. 2017. 虎斑蝶实验种群生物学特征研究[J]. 应用昆虫学报, 54(2): 279–291. |
| 陈祯. 2010. 虎斑蝶生理生态学研究[D]. 昆明: 西南林业大学. |
| 吕龙石, 金大勇, 朴锦. 2004. 温度和光周期对麝凤蝶生长发育的影响[J]. 昆虫知识, 41(6): 572–574. DOI:10.7679/j.issn.2095-1353.2004.145 |
| 易传辉, 陈晓鸣, 史军义, 等. 2007. 光周期和温度对美凤蝶幼虫发育历期的影响[J]. 林业科学研究, 20(4): 547–550. |
| 易传辉, 陈晓鸣, 史军义, 等. 2009. 光周期对柑橘凤蝶幼虫生长发育及存活的影响[J]. 西北农业学报, 18(1): 162–164, 169. DOI:10.7606/j.issn.1004-1389.2009.01.035 |
| 张孝羲. 2011. 昆虫生态及预测预报(第三版)[M]. 北京: 中国农业出版社: 35-36. |
| 周尧. 1998. 中国蝶类志(修订版)[M]. 郑州: 河南科学技术出版社: 382-383. |
| Atluri JB, Rayalu MB, Deepika DS. 2010. Life history and larval performance of the common tiger butterfly, Danaus genutia Cramer (Lepidoptera; Rhopalocera; Danaidae)[J]. The Bioscan, 5(3): 511–515. |
| Bhuyan M, Kataki D, Deka M, et al. 1999. Nectar host plant selection and floral probing by the India butterfly Danaus genutia (Nympahlidae)[J]. Journal of Research on the Lepidoptera, 38(1): 79–84. |
| Goehring L, Oberhauser KS. 2002. Effects of photoperiod, temperature, and host plant age on induction of reproductive diapause and development time in Danaus plexippus[J]. Ecological Entomology, 27(6): 674–685. DOI:10.1046/j.1365-2311.2002.00454.x |
| Igarashi S, Fukuda H. 2000. The life histories of Asian butterflies[M]. Tokyo: Tokai University Press: 244-245. |
| Malcolm SB, Cockrell BJ, Brower LP. 1987. Monarch butterflyvoltinism:effects of temperature constraints at different latitudes[J]. Oikos, 49: 77–82. DOI:10.2307/3565556 |
| Meyer CE. 1995. Notes on the life history of Danaus genutia alexis (Waterhouse and Lyell)(Lepidoptera:Nymphalidae)[J]. Australian Entomologist, 22(4): 137–139. |
| Robinson GS, Ackery PR, Kitching IJ, et al. 2001. Hostplants of the moth and butterfly caterpillars of the Oriental Region[M]. London: The Natural History Museum. |
| Yoshio M, Ishii M. 2004. Photoperiod effects on pupal development in two tropical Papilio butterflies (Lepidopera, Papilionidae)[J]. Transactions of the Lepidopterological Society of Japan, 55(4): 275–279. |