 2018, Vol. 37
2018, Vol. 37扩展功能
文章信息
- 余艳萍, 张亢亢, 刘颖
- YU Yanping, ZHANG Kangkang, LIU Ying
- 动物交流声波句法结构研究进展
- Advanced in Syntactic Structures of Vocal Communications in Animals
- 四川动物, 2018, 37(1): 108-120
- Sichuan Journal of Zoology, 2018, 37(1): 108-120
- 10.11984/j.issn.1000-7083.20170301
-
文章历史
- 收稿日期: 2017-09-27
- 接受日期: 2017-11-29






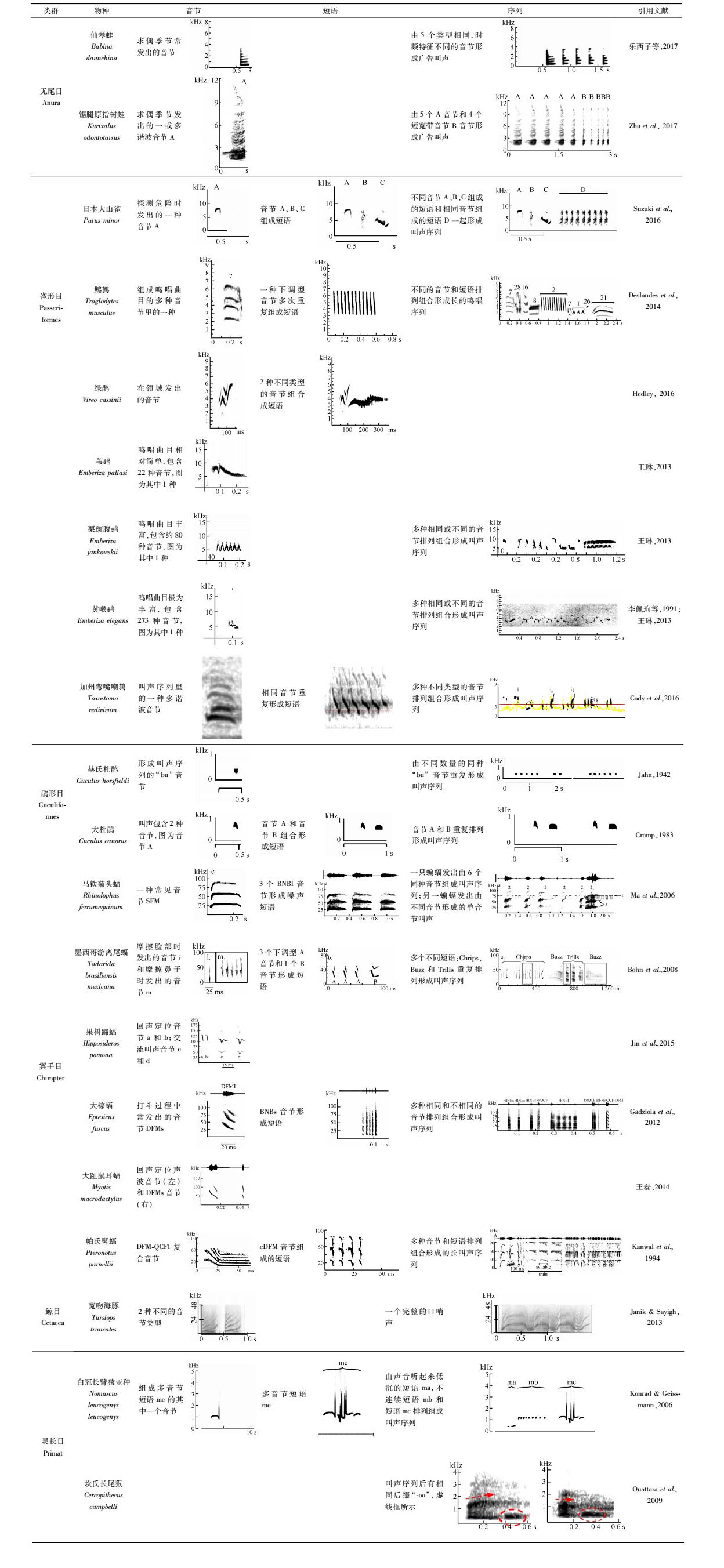
动物通过发出交流叫声传递丰富的信息,包括环境背景、发声者的身体状态、繁殖条件、情感表达,以及表现某种行为的可能性等。交流叫声主要用于求偶交配(Searcy et al., 1985)、保护领域和食物资源(Ma et al., 2006)、躲避天敌(Townsend et al., 2010)、维持母婴关系(Jin et al., 2015)、个体识别(Mates et al., 2015)等。因此,交流叫声在动物的整个生活史中都起着无可替代的作用(Chaverri et al., 2013)。针对动物交流叫声的研究已在多个动物类群中开展,如鸟类(Lemon et al., 1985;Hedley,2016;Taylor et al., 2017)、鲸目Cetacea(Janik,2000)、啮齿目Rodentia(Holy & Guo,2005)、非人灵长目Primates(Clarke et al., 2006;de la Torre & Snowdon,2009)、蝙蝠(翼手目Chiroptera)(Bohn et al., 2008)、蹄兔目Hyracoidea(Kershenbaum et al., 2012)和无尾目Anura (Tessarolo et al., 2016)等。
声波是叫声的物理表现形式,交流声波的基本组成单位是音节(syllable)。不同动物的声波复杂程度不一样,有的动物声波曲目(vocal repertoire)复杂多样,包含的音节类型极为丰富,可分为由单一组分构成的简单音节(simple syllable),如马铁菊头蝠Rhinolophus ferrumequinum的正弦调频(SFM)音节(附表);由2种或以上组分构成的复合音节(composite syllable),如噪音-正弦调频(NB-SFM)音节(Kanwal et al., 1994;Ma et al., 2006)(附表)。音节还可以通过不同的排列组合方式构成短语(phrase),并进一步构成更为复杂的声波序列(vocal sequence),如墨西哥游离尾蝠Tadarida brasiliensis mexicana的求偶叫声由3个短语“chrip-buzz-trill”组成(附表)。研究表明,越复杂的交流叫声信号携带的信息越多,能传达更丰富、完整的信息,有利于动物交流(Bohn et al., 2008)。例如,鸟类在繁殖季节的求偶叫声不仅鸣唱时间长,而且声波序列里包含很多相同或不同的音节,这些音节通过多样的方式聚合在一起形成复杂的求偶叫声。由于性选择压力,雌性个体倾向于选择具有复杂鸣唱曲目的雄性个体,这代表该雄性个体可能具有较好的身体质量(Searcy,1984;Searcy et al., 1985);而识别压力促使不同的动物个体发出复杂的叫声,有助于个体识别或者群体识别(Bohn et al., 2008);在领域竞争时,雄性个体也会尽量避开拥有更多叫声曲目的对手(Dobson & Lemon,1975)。因此,动物交流叫声信号的多样化和复杂化是其适应性进化的结果(Luo et al., 2017)。
动物只能发出有限的音节类型,但它们能够通过音节不同的排列组合方式,形成持续时间更长、组成类型更丰富的声波序列。而不同音节类型的排列组合规律就称为句法结构(syntactic structure)(Bohn et al., 2008;Weiss et al., 2014)。句法结构的存在使动物不用产生新音节,通过有限的音节类型传递“无限”的声学含义。如Jahn(1942)发现韩国赫氏杜鹃Cuculus horsfieldi有“bubu…”和“bububububu”2种鸣唱型(附表),说明赫氏杜鹃具有单音节重复的句法规则。此外,室内研究发现,非洲灰鹦鹉Psittacus erithacus、宽吻海豚Tursiops truncatus和倭黑猩猩Pan paniscus具备句法结构能力,能根据人的提示发出具有不同复杂程度的英文片段(Kako,1999)。在后续的声学研究中,鸟类、鲸类、非人灵长类存在句法结构的证据不断被发现,除此之外,啮齿目(Holy & Guo,2005)、翼手目(Bohn et al., 2008)、蹄兔目(Kershenbaum et al., 2012)和无尾目动物(乐西子等,2017;Yue et al., 2017)也已经被证明存在句法结构,且句法结构的分化受到不同选择压力的作用。研究动物交流声波句法结构对于探索动物交流声波的形成与维持机制具有重要意义;同时,由于动物交流声波的组成方式与人类语言的组成方式有相同之处,所以对动物交流声波句法结构的研究有助于揭示人类语言的形成与进化机制。因此,本文对目前动物声学领域已有的动物交流叫声句法研究进行综述,并在此基础上提出未来动物交流声波及句法结构研究的重点与方向,以期对深入研究动物声学行为提供帮助与指导。
1 动物交流声波的类型及多样性 1.1 交流声波的分类交流声波类型多样,对交流声波的分类主要采取以下3种方法:首先,根据人耳听到的声音类型将交流声波描述为:尖叫声(scream)、咔咔声(twitter)和口哨声(whistle)(Janik,2000)等。然而这种主观分类存在不足之处,因为有一些交流声波可能是超声波,人耳无法听到。所以,另一种分类方法是根据发声的行为背景进行划分:打斗发出的交流声波被称为激进叫声(aggressive calls)或警示叫声(warning calls)(Fernandez et al., 2014)、交配季节里吸引异性时发出的求偶叫声一般称为鸣唱叫声(song calls)(Bohn et al., 2008)、遇险时发出的求救叫声(distress calls)(Townsend et al., 2010)、捍卫领域时发出的领域叫声(territory calls)(Ma et al., 2006)以及母蝠寻找幼蝠的指导叫声(director calls)和幼蝠的隔离叫声(isolation calls)(附表:果树蹄蝠Hipposideros pomona)(Jin et al., 2015)等。虽然可以根据发声背景确定交流叫声类型,但这种分类方法依然存在不足,如在不同背景下,同一个体可能会发出结构相似的交流声波。因此,比较客观的划分方法是根据声波组成元素和结构进行划分。大部分交流声波都可看作由3种简单类型的声音元素组成,即调频声波(frequency modulation,FM)、恒频声波(constant frequency,CF)和噪声串(noise burst,NB)。根据这些简单声波元素的组分,可以对复杂交流声波命名。如马铁菊头蝠的长宽带噪音(BNBl)音节就是根据NB再加上描述带宽的broadband和长持续时间long的首字母分别作为前缀和后缀组成(附表)。
1.2 交流声波的多样性不同动物由于长期适应不同的生态和社会环境,发声器官有所差异,发出的交流声波也不同;同一物种的不同性别、不同个体之间,交流声波也会存在差异;甚至同一个体在同一背景的不同表现程度下发出的叫声都存在差异。
在叫声复杂的众多动物中,由于幼鸟学习鸣唱(song)的过程与人类婴儿学习说话的过程存在行为、神经、基因等相似性,鸟类能够发出比其他物种更复杂的鸣声,因此,鸟类一直是研究人类语言起源的模式物种。鸟类的鸣唱是一种非常重要的行为学特征。Catchpole(2008)把鸣唱定义为:一般在繁殖期由雄鸟发出的长且复杂的鸣声。对于鸣禽来讲,不同物种具有不同的曲目大小,这反映了鸟类鸣唱的复杂性和多样性。同为鹀属Emberiza物种,苇鹀E. pallasi鸣唱语句相对简单,包含22种音节,且语句中音节个数少,平均为3.86个±0.17个(n=82);栗斑腹鹀E. jankowskii的鸣唱序列更加丰富,包含音节更多,约80种音节类型;而黄喉鹀E. elegans的鸣唱曲目则复杂得多,共有87种鸣唱语句,包含的音节多达273个(n=215)(李佩珣等,1991;王琳,2013)(附表)。
在哺乳动物类群中,鲸目宽吻海豚的同一个群体内不同个体之间,就存在超过20种共享音节,并且共享音节在频率、结构上存在个体差异。除共享音节外,不同个体还有特有音节,有利于个体的识别过程(Janik & Sayigh,2013)。非人类灵长目动物具有高度社会化的社群结构,不同的家庭成员会发出具有不同声谱结构的叫声。如在白颊长臂猿亚种Nomascus leucogenys leucogenys家庭里,成年雌性发出由不同短语组成的声波序列,成年雄性发出的叫声在短语组成上跟雌性差异很大(Konrad & Geissmann,2006)。翼手目帕氏髭蝠Pteronotus parnellii的交流声波里存在33种音节类型,包括19种简单音节,14种复合音节(Kanwal et al., 1994)(附表)。大棕蝠Eptesicus fuscus甚至能在同一背景的不同程度下发出不同的叫声,如在低激进和中激进程度下发出以短的下调型调频(DFMs)音节为主的叫声,而在高激进程度下发出以其他音节为主的叫声(Gadziola et al., 2012)(附表)。
综上,动物叫声包含多种音节类型,从几种到几百种不等,然而很多动物的声波序列并不是随机发出,音节也不是随机组合,而是具有一定的句法规律。现总结已有研究,对动物中出现的句法结构进行归类。
2 动物的句法结构 2.1 单音节重复的叫声或鸣唱动物句法结构的类型多种多样,体现在音节的多样性和排列组合的多样性上。有些鸟类的鸣唱曲目只有简单的类型,鸣唱表现为单调的音节重复(Catchpole,2008)。如韩国赫氏杜鹃的鸣唱类型为同种音节的重复,“A”表示音节类型,赫氏杜鹃的鸣唱表现为“AA…”型重复,或者“AAAAA…”型和“AAA…”型,鸣唱曲目越长,音节重复次数越高,传达的感情越强烈。美洲红尾鸲Setophaga ruticilla也有类似的单音节重复鸣唱型,在一个长的鸣唱回合中可以唱出3种或者更多的鸣唱类型(Lemon et al., 1985),其中,一种为“…AAAAA…”的类型也是单音节的重复。音节重复的次数跟物种本身所具有的感情状态有关,当情感比较强烈时,可能会发出音节重复次数更多、声波序列更长的鸣唱型。蝙蝠也存在单音节重复的现象。Gadziola等(2012)对大棕蝠在不同激进程度下的打斗叫声进行研究,发现大棕蝠在低等程度的打斗过程中发出的声波序列由4~6个音节组成,其中94%是DFMS音节,声波序列显示为DFMS音节的简单重复(附表)。马铁菊头蝠的领域叫声也有包含多个单音节重复的现象,比如BNBl音节的重复,重复数量一般是2~4个音节(附表);还有SFM的重复,重复数量一般为3~5个音节。但是,马铁菊头蝠的领域叫声更为复杂,多是由重复音节组成短语,然后再由短语组成声波序列(Ma et al., 2006)。
2.2 双音节重复的叫声或鸣唱除单音节重复的句法结构外,动物还具有双音节重复的句法结构。大杜鹃Cuculus canorus夏季在我国的分布非常广泛,几乎遍布全国(雷富民等,1997)。Cramp(1983)将大杜鹃的领域鸣唱描述为“cu-coo”,其中“cu”和“coo”是2个不同的音节,分别用“A”和“B”表示。其鸣唱型表现为双音节的重复,如“AB…”型、“ABB…”型和“ABBBBB”型,后2种可以看做是“AB…”型的变体。同单音节重复一样,当双音节重复次数越多时,鸣唱序列就越长,曲目多样性越高,表达的情感可能越强烈,更能达到求偶成功的目的。在另一些物种中,比如墨西哥游离尾蝠的领域叫声和求偶叫声都是由不同的短语组成,而短语是由不同或者相同的音节排列组合而成,比如3个音节A和1个音节B按顺序排列形成一个短语Chirp,这种短语形成方式也是双音节重复排列的句法结构(Bohn et al., 2008, 2013)。雄性锯腿原指树蛙Kurixalus odontotarsus的求偶鸣唱采用2种音节重复排列组合的方式,如5个音节A和4个音节B组成的声波序列(Zhu et al., 2017)(附表)。
2.3 由固定短语组合形成的叫声或鸣唱有些鸟类的句法结构相对单一,表现为几个简单音节的重复,另一些鸟类的句法结构则更加复杂。如对鹪鹩Troglodytes musculus的领域叫声进行研究,发现其能发出42种不同的音节,由这些音节形成短语的过程复杂多样,短语可以由同种或不同种音节组合形成(Deslandes et al., 2014)。对绿鹃Vireo cassinii的研究发现,每只绿鹃能发出40~60条短语,其中,每2~7条短语又可组合成一段有意义的声波序列,并且绿鹃对这些短语的使用具有一定规则,有些短语使用频率极高,而另一些短语的使用频率极低,说明绿鹃具有非随机的句法结构规律(Hedley,2016)。加州弯嘴嘲鸫Toxostoma redivivum具有大量的鸣唱曲目,能发出多种短语类型,其中2~3个短语一起发出,表达一个鸣唱主题;之后,另外2~3个短语组合在一起,表达第二个主题;该主题结束之后,又由另外的短语组合在一起,表达第三个主题。该研究表明,加州弯嘴嘲鸫在鸣唱阶段有3个鸣唱主题,且每个主题都具有不同的中心语句,这些中心语句由相对固定的2~3个短语组成,展现了高度结构化和复杂化的叫声组合规律(Cody et al., 2016)。
2.4 由固定音节或短语起始的叫声或鸣唱动物叫声包含起始音节(或短语)、中间组分、结束组分。由于每个组分音节和短语的数量和类型变化多样,导致整个叫声复杂多变,序列长度也不尽相同,但整个序列总是围绕核心不变的组分展开。如大趾鼠耳蝠Myotis macrodactylus的交流叫声具有多种不同组分,包括4种单音节和37种多音节。虽然每条声波序列里使用的音节类型和数量存在差异,但所有叫声类型都遵循一个基本规则,即所有声波序列均由DFM音节开头,然后与其他音节组合形成复杂的多音节叫声(王磊,2014)。大棕蝠在较激烈的打斗过程中会发出以2种音节起始的叫声,一种是以torQCF音节起始,然后转化成rBNBs音节;另一种是以rBNBs音节起始,然后转化成rBNBl音节或rBNBs音节的重复(Gadziola et al., 2012)。求偶季节,墨西哥游离尾蝠在当陌生个体接近领域时会发出特定的叫声,这些叫声都是由固定短语Chirp开始,随后展示出完整的Chirp-Buzz-Trill-Buzz结构,该研究揭示了墨西哥游离尾蝠的求偶叫声句法结构(Bohn et al., 2008)。随后,Bohn等(2013)又进行了回放实验,实验结果表明,在面对不同的回放刺激时,墨西哥游离尾蝠会发出不同的交流叫声回应刺激,但是所有的回应叫声都是以短语Chirp为起始组分,然后再与其他短语一起组成声波序列。由于动物叫声各组分的时频特征和声谱结构具有明显差异,动物对不同组分表现出不同的行为响应,推测叫声的不同组分具有不同的意义(Williams & Staples,1992;Tang et al., 2001;Suzuki et al., 2016)。乐西子等(2017)通过采集仙琴蛙Babina daunchina的回放刺激和脑电信号,证明了蛙类叫声的第一个音节在声音通讯系统中起至关重要的作用(Yue et al., 2017)。
2.5 由固定音节结束的叫声或鸣唱灵长类动物除了具有复杂的音节排列组合方式外,还会在叫声的最后加上一个固定的声学元素。最具有代表性的是坎氏长尾猴Cercopithecus campbelli在领域范围内发出的警示叫声。当有捕食者接近或进入它们的领域范围时,雄性坎氏长尾猴会在领域叫声后面加上一个类似于人类尾音的“-oo”后缀,虽然警示叫声的其他声学组分都是可变的,但这个后缀却是固定的(Williams & Staples,1992),这样做是为了改变原来声波序列的意思以警示同伴,最终达到保护同伴的目的(Ouattara et al., 2009;Schlenker et al., 2014)。
3 动物交流声波句法结构的地理进化及进化驱动力不同种群的动物长期适应于各自的生活环境,声学表型也发生了适应性进化。地理距离和地理隔离导致同一物种不同种群之间的生态环境存在差异,比如温度、湿度、气压、植被结构等,而这些因素直接影响了声音信号的传播与扩散。此外,遗传因素、社会选择压力、文化漂变等因素会直接或者间接影响动物的句法结构。同一物种不同地理种群间的声音信号差异被称为声学分歧(acoustic divergence),当种群间存在基因流时,这种声学分歧被称为方言(dialect);当有长的地理距离或隔离阻碍种群间基因流时,这种声学分歧则被称为声学地理变化(geographic variation)(Marler & Tamura,1962;Nottebohm,1969;Conner,1982;Lameira et al., 2010)。关于动物交流声波进化的解释,有3种常见互不排斥的经典假说。
3.1 生态复杂性假说生态复杂性假说认为很多生态因素通过多种方式定向地影响动物交流声波复杂性的进化(Ord & Garcia-Porta,2012)。由于地理距离和地理隔离的存在,不同种群的栖息地可能具有不同的地形、地貌和海拔等特征,从而直接影响了不同种群的生态环境,如温度、湿度、植被结构等(Janes et al., 2017)。这些生态环境可能通过影响动物的形态结构(发声器官)来影响发声方式,导致不同句法结构的产生。同时,生态环境的不同也会直接影响发声者声音信号的传播与接收者声音信号的接收,因此,生态环境压力促使不同种群的动物句法结构分化。温、湿度等生态因子也直接影响猎物和捕食者的丰富度,导致发声者面临不同的捕食和被捕食压力,促使它们发出具有不同结构和功能的叫声。如Davidson和Wilkinson(2002)对特立尼达拉岛4个种群大银线蝠Saccopteryx bilineata的研究发现,这4个种群在叫声曲目上存在很大差异,其中一个种群的音节种类和数量远超过其他种群,说明大银线蝠至少在音节水平产生了句法差异。Bjørgesæter等(2004)对6个种群的雄性海豹Phoca vitulina叫声进行研究,发现这6个种群的叫声类型存在地理差异,并且这种叫声差异是长期适应繁殖地环境条件的结果。灵长类的狐猴Lepilemur ssp.被证明叫声差异跟地理距离存在显著关系,即地理距离越远,叫声差异越大(Mendez-Cardenas et al., 2008)。Ortiz-Ramírez等(2016)对存在地理隔离的不同种群的红顶夜鸫Catharus frantzii叫声进行研究,不仅证明地理隔离是句法结构分化的主要驱动力,还暗示社会选择压力也是驱动句法分化的主要因素。
3.2 社群复杂性假说社群复杂性假说认为动物交流通常与社群结构相关,即随着社群个体间相互作用的频次和行为背景的增加,调节这些相互作用的交流信号趋向于更加复杂(Freeberg et al., 2012)。自然种群中,雄性的繁殖成功率很大程度上取决于性选择,表现为雄性竞争(同性选择)和雌性选择(异性选择)。在无尾类、鸟类、哺乳类等动物类群中,雌性会根据雄性的声音信号作为判断雄性身体甚至是巢穴质量的依据,从而选择适合度高的配偶繁育后代。受性选择压力的作用,雄性会发出重复率更高、持续时间更长、结构更复杂的叫声或鸣唱序列(Russ & Racey,2007)。并且不同种群往往具有不同的性别比,性选择压力会刺激不同种群句法结构分化,导致动物方言和声学地理变化的产生(Podos & Warren,2007;Keighley et al., 2017)。对新疆歌鸲Luscinia megarhynchos的叫声回放实验发现,在回放具有正确音节和短语顺序的鸣唱序列时,实验雄鸟会发出具有其他音节和短语顺序的叫声,目的是避免跟随回放叫声,从而发出不同的叫声吸引雌鸟;而当回放更改过音节、短语顺序的鸣唱序列时,实验雄鸟会发出正确顺序的声波序列,目的在于成为其他雄鸟的领导者,以获得交配权(Weiss et al., 2014)。对日本大山雀Parus minor的组合音节回放实验发现,当日本大山雀听到正常的“ABC-D”音节组合时能做出探测、扫描和接近声源等行为,而当听到人为调换过顺序的“ABC-D”音节组合时则很少做出反应,该实验说明日本大山雀拥有严格的组合句法能力,并能够识别同伴叫声的准确性,从而做出相应行为反应,提高交配成功率(Suzuki et al., 2016)。鸟类严格的句法结构是长期适应性选择压力的结果,为了提高其适合度,获得繁殖权利,雄鸟进化出了复杂的句法结构模式,拥有复杂的声学交流体系(Weiss et al., 2014;Suzuki et al., 2016)。
除性选择压力外,个体或者群体的识别压力也会影响句法分化。个体数量多的种群,个体之间、个体与群体之间面临更大的识别压力(Luo et al., 2017)。在这种识别压力的驱使下,不同个体或者群体之间会进行发声学习以获得更多发声曲目,从而提高叫声多样性与复杂性,促进识别作用。比如宽吻海豚会学习其他个体的叫声,丰富自己的发声曲目,从而在识别过程中占据优势(Janik & Sayigh,2013)。同一种群内的不同个体可能选择性地保留或遗弃本身拥有的部分交流声音信号,学习来自邻近群体的其他个体的声音信号,并在种群内保留下来,长此以往,导致不同地理种群的方言和声学地理变化的产生(Irwin et al., 2008)。
3.3 中性假说基于选择,社群复杂性和生态复杂性假说解释了交流声波句法的适应性进化;基于漂变,中性假说认为在缺乏选择作用的情况下,随机过程也可能促进交流声波句法的分化(Grant & Grant,2009)。遗传漂变在进化过程中可能附带地增加了交流声波句法的复杂性,从而导致亲缘关系较近的物种或种群间具有相似的交流声波,反之亦然(Irwin et al., 2008;Benedict & Bowie,2009;Campbell & Phelps,2010;Jiang et al., 2010)。Wright和Wilkinson(2001)对已经被证明存在方言的亚马逊鹦鹉进行遗传结构和声学分歧研究,发现遗传距离近的种群间声学结构类似,如在黄颈亚马逊鹦鹉Amazona auropalliata中发现的方言,在其姐妹种黄冠亚马逊鹦鹉Amazona ochrocephala中也被发现。Luo等(2017)对不同种蝙蝠的交流声波差异和进化驱动力进行研究,发现系统发育成分在蝙蝠声学差异上起至关重要的作用。在灵长类侏儒狨猴Callithrix (Cebuella) pygmaea pygmaea的方言形成过程中,遗传漂变与其他因子一起对句法结构分化起重要作用(de la Torre & Snowdon,2009)。这些研究结果表明,句法结构的差异性或相似性与种群间遗传结构具有密切相关性,遗传结构和其他影响因子一起介导句法结构分化。
在长期的社会适应过程中,每个种群的动物由于所处文化背景的不同,形成了各个种群特有的声文化结构(Leader et al., 2008)。与遗传漂变一样,动物的叫声也会发生随机的改变,称为文化漂变(Sun et al., 2013)。这种声音信号的随机漂变可能会提高或者降低动物的适合度,当发生漂变后的声音信号有利于提高个体和群体的适合度时,该声音信号便在种群内保留下来,长此以往,可能导致不同种群间的声学结构,包括句法结构差异越来越大(Janes et al., 2017)。研究者对不同地理种群岩兔Procavia capensis的句法结构进行研究,实验结果表明,在一定范围内,句法结构差异跟地理距离呈正相关,随着地理距离的增加,不同岩兔种群之间的句法结构差异越大,该研究不仅证明了蹄兔目动物方言的存在,同时还讨论了岩兔句法结构差异的原因可能是相邻种群之间不精确的声音信号复制的结果。如岩兔在学习临近种群其他个体的叫声时,发生了声音复制上的错误,使声波序列里的一些声学组分缺失、增加,甚至是改变,这种“错误”的声音学习会加大相邻种群间句法结构的差异(Kershenbaum et al., 2012)。
4 结论与展望关于动物句法的研究,本文总结分析了已有的文献报导,得到以下结论:1)动物句法研究对象:目前句法结构研究已在无尾类、鸟类和哺乳类动物中开展,而在鸟类中开展的句法研究最为丰富;2)研究内容:目前动物句法研究内容多围绕句法结构的剖析、句法功能的探究、句法结构的分化以及分化驱动力开展,缺乏对句法形成机制的研究,如神经机制,生理机制等;3)研究水平:句法研究水平多为音节水平和短语水平,缺乏在完整声波序列上的句法研究。
综上,未来的动物交流声波句法结构研究可以集中于以下科学问题:1)在声波序列水平开展动物句法研究,从整体出发,揭示动物叫声的规律以及句法的整体变化;2)对于已经被证明存在交流声波句法的动物,应该着力于句法的形成机制,加强对句法的神经调控和生理调节机制的研究;3)应该拓宽句法结构的研究对象,开展更多动物句法的研究,丰富句法结构,提供更多动物句法的证据。
在已有研究对象中,针对其开展句法研究的必要性和优缺点,建议在各动物类群中开展以下研究:1)雄蛙在繁殖季节靠鸣声吸引雌性个体,所以,蛙类的鸣唱是其求偶繁殖过程中最主要的生物学表型。虽然蛙类适于开展声学实验,但是其句法也是近年来才被报导,且限于少数物种(乐西子等,2017),因此,建议未来开展不同种蛙类句法证据的相关研究;2)鸟类由于叫声的复杂性以及跟人类语言独特的相似性,一直以来都被作为研究动物交流声波的关键模型,在句法结构方面更是做出了突出贡献,因此,应继续深入开展鸟类句法研究,尤其是句法形成机制的相关研究;3)哺乳类是动物界进化地位最高的自然类群,现已发现鲸目、啮齿目、蹄兔目、翼手目、灵长目存在句法结构。在哺乳动物类群中,翼手目动物相比于其他动物具有体型小、种类丰富、数量多、易于驯养、便于进行野外实验和室内实验等优点,重要的是翼手目动物长期适应夜空生态位,基本依靠交流声音信号进行母婴交流、求偶交配、维持社群关系等,从而进化出了丰富的交流叫声,但是目前仅证明蝙蝠科的大棕蝠、犬吻蝠科的墨西哥游离尾蝠和菊头蝠科的马铁菊头蝠存在句法结构现象,因此,翼手目应该作为未来句法研究的重要对象。综上,动物句法研究虽已取得显著进展,但仍有很多问题需要不断探索,今后的研究应更全方位、多层次、多角度揭示动物句法的形成机制和分化方向,进而为人类语言的起源与进化提供一定参考。
| 雷富民, 郑作新, 尹祚华. 1997. 纵纹腹小鸮在中国的分布、栖息地及各亚种的梯度变异(鸮形目:鸱鸮科)[J]. Zoological Systematics, 3: 327–334. |
| 李佩珣, 于学锋, 李方满. 1991. 繁殖期黄喉鹀的领域鸣唱及其种内个体识别[J]. 动物学研究, 2: 163–168. |
| 王磊. 2014. 大趾鼠耳蝠声波发育、多样性及其生态可塑性研究[D]. 长春: 东北师范大学. http://cdmd.cnki.com.cn/Article/CDMD-10200-1014375248.htm |
| 王琳. 2013. 八种鹀属鸟类鸣声特征及其适应性进化研究[D]. 长春: 东北师范大学. http://cdmd.cnki.com.cn/Article/CDMD-10200-1013363531.htm |
| 乐西子, 江帆, 薛飞, 等. 2017. 仙琴蛙广告鸣叫中不同音节生物学意义的差异研究[J]. 四川动物, 36(3): 241–248. DOI:10.11984/j.issn.1000-7083.20160313 |
| Benedict L, Bowie RC. 2009. Macrogeographical variation in the song of a widely distributed African warbler[J]. Biology Letters, 5(4): 484–487. |
| Bjørgesæter A, Ugland KI, Bjørge A. 2004. Geographic variation and acoustic structure of the underwater vocalization of harbor seal (Phoca vitulina) in Norway, Sweden and Scotland[J]. Journal of the Acoustical Society of America, 116(1): 2459–2468. |
| Bohn KM, Schmidtfrench B, Ma ST, et al. 2008. Syllable acoustics, temporal patterns, and call composition vary with behavioral context in Mexican free-tailed bats[J]. Journal of the Acoustical Society of America, 124(3): 1838–1848. |
| Bohn KM, Smarsh GC, Smotherman M. 2013. Social context evokes rapid changes in bat song syntax[J]. Animal Behaviour, 85(6): 1485–1491. |
| Campbell P, Phelps SM. 2010. Geographic variation in the songs of neotropical singing mice:testing the relative importance of drift and local adaptation[J]. Evolution, 64(7): 1955–1972. |
| Catchpole C. 2008. Bird song:biological themes and variations[M]. Hong Kong: Cambridge University Press. |
| Chaverri G, Gillam EH, Kunz TH. 2013. A call-and-response system facilitates group cohesion among disc-winged bats[J]. Behavioral Ecology, 24(2): 481–487. DOI:10.1093/beheco/ars188 |
| Clarke E, Reichard UH, Zuberbühler K. 2006. The syntax and meaning of wild gibbon songs[J]. PLoS ONE, 1(1): e73. DOI:10.1371/journal.pone.0000073 |
| Cody ML, Stabler E, Taylor CE. 2016. Structure, syntax and "small-world" organization in the complex songs of California thrashers (Toxostoma redivivum)[J]. Bioacoustics, 25(1): 41–54. DOI:10.1080/09524622.2015.1089418 |
| Conner DA. 1982. Dialects versus geographic variation in mammalian vocalizations[J]. Animal Behaviour, 30(1): 297–298. |
| Cramp S. 1983. Handbook of the birds of Europe, the middle east and north Africa[M]. Oxford: Oxford University Press: 482-489. |
| Davidson SM, Wilkinson GS. 2002. Geographic and individual variation in vocalizations by male Saccopteryx bilineata (Chiroptera:Emballonuridae)[J]. Journal of Mammalogy, 83(2): 526–535. DOI:10.1644/1545-1542(2002)083<0526:GAIVIV>2.0.CO;2 |
| de la Torre S, Snowdon CT. 2009. Dialects in pygmy marmosets? Population variation in call structure[J]. American Journal of Primatology, 71(4): 333–342. |
| Deslandes V, Faria LR, Borges ME, et al. 2014. The structure of an avian syllable syntax network[J]. Behavioural Processes, 106(3): 53–59. |
| Dobson CW, Lemon RE. 1975. Re-examination of monotony threshold hypothesis in bird song[J]. Nature, 257(5522): 126–128. |
| Fernandez AA, Fasel N, Knörnschild M, et al. 2014. When bats are boxing:aggressive behaviour and communication in male Seba's short-tailed fruit bat[J]. Animal Behaviour, 98(98): 149–156. |
| Freeberg TM, Dunbar RI, Ord TJ. 2012. Social complexity as a proximate and ultimate factor in communicative complexity[J]. Philosophical Transactions of the Royal Society of London, 367(1597): 1785–1801. |
| Gadziola MA, Grimsley JM, Faure PA, et al. 2012. Social vocalizations of big brown bats vary with behavioral context[J]. PLoS ONE, 7(9): e44550. DOI:10.1371/journal.pone.0044550 |
| Grant PR, Grant BR. 2009. The secondary contact phase of allopatric speciation in Darwin's finches[J]. Proceedings of the National Academy of Sciences of the United States of America, 106(48): 20141–20148. |
| Hedley RW. 2016. Composition and sequential organization of song repertoires in Cassin's vireo (Vireo cassinii)[J]. Journal of Ornithology, 157(1): 13–22. DOI:10.1007/s10336-015-1238-x |
| Holy TE, Guo Z. 2005. Ultrasonic songs of male mice[J]. PLoS Biology, 3(12): e386. DOI:10.1371/journal.pbio.0030386 |
| Irwin D, Thimgan M, Irwin J. 2008. Call divergence is correlated with geographic and genetic distance in greenish warblers (Phylloscopus trochiloides):a strong role for stochasticity in signal evolution?[J]. Journal of Evolutionary Biology, 21(2): 435–448. DOI:10.1111/jeb.2008.21.issue-2 |
| Jahn H. 1942. Zur Oekologie und Biologie der Vögel Japans[J]. Journal of Ornithology, 90(1-2): 7–302. |
| Janes SW, Ryker L, Ryan RM. 2017. Geographic variation in type Ⅰ dialects of hermit warblers:does fragmented habitat promote variation in song?[J]. Journal of Ornithology, 158(2): 421–430. DOI:10.1007/s10336-016-1420-9 |
| Janik VM, Sayigh LS. 2013. Communication in bottlenose dolphins:50 years of signature whistle research[J]. Journal of Comparative Physiology A Neuroethology Sensory Neural & Behavioral Physiology, 199(6): 479–489. |
| Janik VM. 2000. Whistle matching in wild bottlenose dolphins (Tursiops truncatus)[J]. Science, 289(5483): 1355–1357. |
| Jiang T, Liu R, Metzner W, et al. 2010. Geographical and individual variation in echolocation calls of the intermediate leaf-nosed bat, Hipposideros larvatus[J]. Ethology, 116(8): 691–703. |
| Jin L, Yang S, Kimball RT, et al. 2015. Do pups recognize maternal calls in pomona leaf-nosed bats, Hipposideros pomona?[J]. Animal Behaviour, 100: 200–207. |
| Kako E. 1999. Elements of syntax in the systems of three language-trained animals[J]. Animal Learning & Behavior, 27(1): 1–14. |
| Kanwal JS, Matsumura S, Ohlemiller K, et al. 1994. Analysis of acoustic elements and syntax in communication sounds emitted by mustached bats[J]. Journal of the Acoustical Society of America, 96(3): 1229–1254. |
| Keighley MV, Langmore NE, Zdenek CN, et al. 2017. Geographic variation in the vocalizations of Australian palm cockatoos (Probosciger aterrimus)[J]. Bioacoustics, 26(1): 91–108. |
| Kershenbaum A, Ilany A, Blaustein L, et al. 2012. Syntactic structure and geographical dialects in the songs of male rock hyraxes[J]. Proceedings:Biological Sciences, 279(1740): 2974–2981. DOI:10.1098/rspb.2012.0322 |
| Konrad R, Geissmann T. 2006. Vocal diversity and taxonomy of Nomascus in Cambodia[J]. International Journal of Primatology, 27(3): 713–745. |
| Lameira AR, Delgado RA, Wich SA. 2010. Review of geographic variation in terrestrial mammalian acoustic signals:human speech variation in a comparative perspective[J]. Journal of Evolutionary Psychology, 8(4): 309–332. |
| Leader N, Geffen E, Mokady O, et al. 2008. Song dialects do not restrict gene flow in an urban population of the orange-tufted sunbird, Nectarinia osea[J]. Behavioral Ecology & Sociobiology, 62(8): 1299–1305. |
| Lemon RE, Cotter R, Macnally RC, et al. 1985. Song repertoires and song sharing by American redstarts[J]. Condor, 87(4): 457–470. |
| Luo B, Huang X, Li Y, et al. 2017. Social call divergence in bats:a comparative analysis[J]. Behavioral Ecology, 28(2): 533–540. |
| Ma J, Kobayasi K, Zhang S, et al. 2006. Vocal communication in adult greater horseshoe bats, Rhinolophus ferrumequinum[J]. Journal of Comparative Physiology A, 192(5): 535–550. |
| Marler P, Tamura M. 1962. Song "dialects" in three populations of white-crowned sparrows[J]. Condor, 64(5): 368–377. DOI:10.2307/1365545 |
| Mates EA, Tarter RR, Ha JC, et al. 2015. Acoustic profiling in a complexly social species, the American crow:caws encode information on caller sex, identity, and behavioural context[J]. Bioacoustics, 24(1): 63–80. DOI:10.1080/09524622.2014.933446 |
| Méndez-Cárdenas M, Randrianambinina B, Rabesandratana A, et al. 2008. Geographic variation in loud calls of sportive lemurs (Lepilemur ssp.) and their implications for conservation[J]. American Journal of Primatology, 70(9): 828–838. |
| Nottebohm F. 1969. The song of the chingolo, Zonotrichia capensis, in Argentina:description and evaluation of a system of dialects[J]. Condor, 71(3): 299–315. |
| Ord TJ, Garcia-Porta J. 2012. Is sociality required for the evolution of communicative complexity? Evidence weighed against alternative hypotheses in diverse taxonomic groups[J]. Philosophical Transactions of the Royal Society of London, 367(1597): 1811–1828. DOI:10.1098/rstb.2011.0215 |
| Ortiz-Ramírez MF, Andersen MJ, Zaldívar-Riverón A, et al. 2016. Geographic isolation drives divergence of uncorrelated genetic and song variation in the ruddy-capped nightingale-thrush (Catharus frantzii; Aves:Turdidae)[J]. Molecular Phylogenetics and Evolution, 94(Pt A): 74–86. |
| Ouattara K, Lemasson A, Zuberbühler K. 2009. Campbell's monkeys use affixation to alter call meaning[J]. PLoS ONE, 4(11): e7808. DOI:10.1371/journal.pone.0007808 |
| Podos J, Warren PS. 2007. The evolution of geographic variation in birdsong[J]. Advances in the Study of Behavior, 37(7): 403–458. |
| Russ JM, Racey PA. 2007. Species-specificity and individual variation in the song of male Nathusius' pipistrelles (Pipistrellus nathusii)[J]. Behavioral Ecology and Sociobiology, 61(5): 669–677. |
| Schlenker P, Chemla E, Arnold K, et al. 2014. Monkey semantics:two 'dialects' of Campbell's monkey alarm calls[J]. Linguistics and Philosophy, 37(6): 439–501. DOI:10.1007/s10988-014-9155-7 |
| Searcy WA, Mcarthur PD, Yasukawa K. 1985. Song repertoire size and male quality in song sparrows[J]. Condor, 87(2): 222–228. |
| Searcy WA. 1984. Song repertoire size and female preferences in song sparrows[J]. Behavioral Ecology & Sociobiology, 14(4): 281–286. |
| Sun K, Luo L, Kimball RT, et al. 2013. Geographic variation in the acoustic traits of greater horseshoe bats:testing the importance of drift and ecological selection in evolutionary processes[J]. PLoS ONE, 8(8): e70368. DOI:10.1371/journal.pone.0070368 |
| Suzuki TN, Wheatcroft D, Griesser M. 2016. Experimental evidence for compositional syntax in bird calls[J]. Nature Communications, 7: 10986. DOI:10.1038/ncomms10986 |
| Tang YZ, Zhuang LZ, Wang ZW. 2001. Advertisement calls and their relation to reproductive cycles in Gekko gecko (Reptilia, Lacertilia)[J]. Copeia, 430(1): 248–253. |
| Taylor CE, Brumley JT, Hedley RW, et al. 2017. Sensitivity of California thrashers (Toxostoma redivivum) to song syntax[J]. Bioacoustics, 26(3): 1–12. |
| Tessarolo G, Maciel NM, Ribeiro MA, et al. 2016. Geographic variation in advertisement calls among populations of Dendropsophus cruzi (Anura:Hylidae)[J]. The Herpetological Journal, 26(3): 219–227. |
| Townsend SW, Hollén LI, Manser MB. 2010. Meerkat close calls encode group-specific signatures, but receivers fail to discriminate[J]. Animal Behaviour, 80(1): 133–138. |
| Weiss M, Hultsch H, Adam I, et al. 2014. The use of network analysis to study complex animal communication systems:a study on nightingale song[J]. Proceedings of the Royal Society of London B:Biological Sciences, 281: 20140460. DOI:10.1098/rspb.2014.0460 |
| Williams H, Staples K. 1992. Syllable chunking in zebra finch (Taeniopygia guttata) song[J]. Journal of Comparative Psychology, 106(3): 278–286. DOI:10.1037/0735-7036.106.3.278 |
| Wright TF, Wilkinson GS. 2001. Population genetic structure and vocal dialects in an amazon parrot[J]. Proceedings of the Royal Society of London B:Biological Sciences, 268(1467): 609–616. |
| Yue X, Fan Y, Xue F, et al. 2017. The first call note plays a crucial role in frog vocal communication[J]. Scientific Reports, 7(1): 10128. DOI:10.1038/s41598-017-09870-2 |
| Zhu B, Wang J, Zhao L, et al. 2017. Male-male competition and female choice are differentially affected by male call acoustics in the serrate-legged small treefrog, Kurixalus odontotarsus[J]. PeerJ, 5: e3980. DOI:10.7717/peerj.3980 |