 2017, Vol. 36
2017, Vol. 36扩展功能
文章信息
- 刁鲲鹏, 李明富, 潘世玥, 顾伟龙, 张晓鸥, 闻丞
- DIAO Kunpeng, LI Mingfu, PAN Shiyue, GU Weilong, ZHANG Xiaoou, WEN Cheng
- 基于红外相机研究脊椎动物在唐家河国家级自然保护区动物尸体分解过程中的作用
- Role of Vertebrates in Wild Animal Corpses Degradation Process by Camera Trap in Tangjiahe National Nature Reserve
- 四川动物, 2017, 36(6): 616-623
- Sichuan Journal of Zoology, 2017, 36(6): 616-623
- 10.11984/j.issn.1000-7083.20170199
-
文章历史
- 收稿日期: 2017-06-22
- 接受日期: 2017-09-20






2. 四川省唐家河国家级自然保护区, 四川青川 628100
2. Tangjiahe National Nature Reserve, Qingchuan, Sichuan Province 628100, China
分解过程是动物残骸或排泄物中的有机物依次分解的过程,由多种生物参与完成,包括食肉动物、食草动物、寄生生物等,参与这个过程的生物都可以被称为分解者(Watson & Carlton,2003)。在一个完整的生态系统中,分解者扮演着一个不可或缺的角色。动、植物的残骸(动物尸体、植物落叶)由分解者作用后将其中的各种物质归还到无机环境中,分解者起着维持生态系统物质循环的作用(黄福贞,1996)。
尸体分解是生态系统中不可缺少的一环,但是对大型动物尸体腐败分解过程的研究比较少。Shean(1993)使用家猪研究尸体在不同环境下的分解过程,结果表明不同的暴露环境能够影响尸体的分解速度,蝇蛆是影响尸体分解速度的主要因素。陈禄仕(2005)研究了人类尸体在自然条件下的腐败分解过程,也认为蝇蛆是尸体分解的主要因素,在没有蝇蛆参与的情况下,尸体主要靠组织自融和腐败细菌进行分解。陈禄仕(2011)对尸体腐败过程中滋生的尸食性蝇类等无脊椎动物进行了系统调查与分类,并对尸体的软组织分解过程进行了研究。彭倩宜等(2009)对嗜尸性甲虫对尸体的腐败、破坏、分解和姿势改变的作用进行了研究。少有描述脊椎动物参与尸体分解的研究。
以往的研究受条件所限,大部分局限于城市、乡村、近郊等人类活动频繁的地区(陈禄仕,2000),多使用家养动物尸体、人类尸体进行实验或者观察(张建刚,符德平,2012)。对于大型野生动物尸体在保护条件良好的保护区内的分解过程少有涉及,使以往研究认为尸体分解主要是由蝇蛆和腐败细菌参与,低估了食肉、食腐动物在尸体分解过程中的贡献,以及尸体对脊椎动物的作用。
唐家河国家级自然保护区近年来的巡护监测中时常发现有大型动物死亡,在2013—2014年巡护监测过程中遇见有蹄动物死亡个体达60头(保护区疫源疫病报告),数量众多的尸体在保护区生态系统中构成了一个不可忽略的因素。以往研究中曾有亚洲黑熊Ursus thibetanus进食动物尸体的记录(胡锦矗,1990),但没有对脊椎动物参与尸体分解的针对性研究。本文研究保护区内脊椎动物参与野生动物尸体分解过程,可以更全面地认识通过尸体连接的生态链,填补目前国内这方面研究的空缺,并为保护区管理提供一个更合理的方法。
1 研究区域概况研究地点为四川省青川县境内的唐家河国家级自然保护区,位于岷山山系龙门山西北段,摩天岭南麓,四川盆地向青藏高原过渡的高山峡谷地带,介于亚热带低地和西部及北部的温带高原之间。地理坐标为104°27′~104°53′E,32°32′~32°41′N,总面积40 000 hm2,白熊坪保护站区域为其核心区域,面积5 000 hm2。保护区在自然地理划分上属于亚热带季风气候区,年平均气温12 ℃,年均降水量1 021.7 mm。保护区内动植物资源丰富,兽类有7目29科101种,鸟类有14目51科266种,爬行类动物有4目8科27种,两栖动物有2目7科20种(四川省唐家河国家级自然保护区管理处等,2012)。
2 研究方法 2.1 死亡动物选取按日常巡护监测经验,大型动物死亡前多游荡在水源地附近。于2014年11月—2015年12月,在唐家河国家级自然保护区白熊坪保护站区域,采用样带法沿巡护监测样线搜寻动物尸体。搜寻频率为日常巡护样线4条,每月4次;季度监测样线11条,每季度1次。样带宽度50 m,4~5人一组均匀散开,沿河谷地带搜寻。在保护区内寻找到8具自然死亡的动物尸体(其中有1只家山羊于保护区的外围保护地带自然死亡,被移至实验区),每头动物尸体旁架设1台红外相机,作为实验组。8具尸体基本信息见表 1。
| 尸体种类 Corpse species |
尸体位置 Corpse position |
海拔 Altitude/m |
红外相机安放时间 Infrared camera placement time |
红外相机回收时间 Infrared camera recovery time |
分解时长 Decomposition time/min |
| 斑羚Naemorhedus goral | 104°70′E,32°59′N | 1 789 | 2015/4/21,11:21 | 2015/4/27,11:31 | 8 660 |
| 羚牛Budorcas taxicolor | 104°70′E,32°60′N | 1 965 | 2015/4/21,14:38 | 2015/6/11,12:24 | 73 306 |
| 羚牛 | 104°66′E,32°60′N | 1 989 | 2015/5/15,10:23 | 2015/7/12,10:48 | 86 425 |
| 羚牛 | 104°68′E,32°60′N | 1 859 | 2015/4/17,11:43 | 2015/7/15,11:08 | 129 565 |
| 山羊(家养) Capra aegagrus hircus (domestic) |
104°68′E,32°60′N | 1 871 | 2015/8/8,12:19 | 2015/10/6,17:13 | 86 694 |
| 斑羚 | 104°67′E,32°60′N | 1 959 | 2015/10/16,18:10 | 2015/11/19,18:39 | 50 429 |
| 野猪Sus scrofa | 104°68′E,32°60′N | 1 837 | 2015/7/26,10:43 | 2015/8/21,14:27 | 39 104 |
| 羚牛 | 104°70′E,32°60′N | 1 890 | 2015/12/11,15:24 | 2016/1/5,09:23 | 37 079 |
对照组:在每具尸体附近的相同生境中安放1台红外相机作为对照,相机点位距离尸体位置30~40 m。
2.2 红外相机设置红外相机型号为Ltl Acorn 6210MC,触发时间为0.8 s。标准设置为一次连续拍摄3张照片,录像30 s,连续2次工作时间间隔0 s。红外相机采取全天候监测,架设高度视尸体死亡地点的自然环境而定,以能清晰拍到整个动物尸体为准,且尽量记录到各种体型的动物。每周检查红外相机1次,如有尸体被大型动物拖离相机视野,则调整红外相机位置与角度继续拍摄。如果尸体没有被拖动,则不对尸体与红外相机进行操作。每2周收集1次红外相机数据,并检查电量,按需更换电池。
2.3 数据提取与分析照片中出现动物即为有效照片。人工识别并记录所拍摄动物的种类、数量、停留时间及行为(路过、进食、玩耍、未知)。2个机位在同一时间拍到的同一动物视为1条有效记录,同一机位对同一动物连续拍摄记为1条有效记录。如果仅有1个机位拍到动物,也视为1条有效记录。
停留时间为每个个体拍摄到第一张照片到最后一张照片的时间段。2张照片间隔不超过10 min的视为同一动物个体,记为1次拍摄事件。
2.4 尸体被完全利用的定义本次实验在于探索白熊坪保护站附近参与尸体能量流动的动物,及探索影响尸体被其他动物利用的原因,因此“完全利用”是一个衡量实验进度的重要指标。
在本次实验中,尸体样本出现以下情况时,被定义为完全利用:
(1) 当实验用尸体的物理破坏程度达到80%或以上,包括头部、躯干及四肢丢失,内脏完全被其他动物利用等,即可食用部分被消耗殆尽(刘哲铭,2016);
(2) 位于尸体点位的红外相机连续10 d不再拍到任何食腐动物前往取食尸体。
3 结果 3.1 参与食腐动物种类实验组8台相机共收集照片10 893张,其中,有效照片3 580张,有效记录326条。
研究发现,参与食腐的脊椎动物有12种,隶属5目8科,具体为亚洲黑熊、野猪Sus scrofa、大嘴乌鸦Corvus macrorhynchos、香鼬Mustela altaica、果子狸Paguma larvata、岩松鼠Sciurotamias davidianus、黄喉貂Martes flavigula、橙翅噪鹛Garrulax elliotii、绿背山雀Parus monticolus、红尾水鸲Rhyacornis fuliginosa、白顶溪鸲Chaimarrornis leucocephalus、铜蜓蜥Sphenomorphus indicus。
3.2 实验组相机与对照组相机物种拍摄率比较同域安放的8台对照组红外相机共收集照片4 960张,其中,有效照片3 286张,有效记录618条。
拍摄到动物18种,隶属5目12科,具体为亚洲黑熊、野猪、大嘴乌鸦、香鼬、果子狸、岩松鼠、小麂Muntiacus reevesi、羚牛Budorcas taxicolor、川金丝猴Rhinopithecus roxellana、藏酋猴Macaca thibetana、豪猪Hystrix hodgsoni、黄喉貂、金猫Catopuma temminckii、豹猫Prionailurus bengalensis、斑羚Naemorhedus griseus、毛冠鹿Elaphodus cephalophus、绿背山雀、橙翅噪鹛。
除去仅出现一、两次的林麝Moschus berezovskii,分别计算实验组相机与对照组相机中动物的拍摄率(capture rate,CR)(章书声等,2012):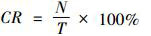
实验组相机与对照组相机拍摄率对比见图 1。2组中不同动物的拍摄率明显不同。部分动物被尸体吸引,如亚洲黑熊、野猪、大嘴乌鸦、果子狸、食虫鸟类、啮齿类、香鼬等。而部分动物则排斥接近尸体,如金猫、豹猫、川金丝猴、藏酋猴、黄喉貂、羚牛、小麂。

|
| 图 1 实验组相机与对照组相机中不同物种的拍摄率对比 Fig. 1 Comparison of the capture rate of animals between experimental and control group |
| |
在尸体分解过程中,不同动物前往参与食腐的时间量明显不同,以动物累积前往参与食腐的时间加和来表示该物种对尸体分解的相对贡献力(relative contribution,RC),即该物种在尸体分解过程中贡献的总时长。例如,当5只大嘴乌鸦共同进食同一尸体10 min,则大嘴乌鸦的相对贡献力为50 min。一般情况下,不同的脊椎动物不会在同一时间进食尸体。不同物种相对贡献力对比见图 2。

|
| 图 2 不同动物的相对贡献力对比 Fig. 2 Comparison of the relative contribution of different animals |
| |
在尸体分解过程中,大嘴乌鸦在数量和时间上占据了最重要的位置。其次为野猪、啮齿动物、亚洲黑熊、香鼬。由于本地的食虫鸟类、两栖爬行类明显不进食尸体,故不统计其相对贡献力。
3.2.2 实际贡献力由于不同动物体型存在差异,进食量也存在差异,相对贡献力并不能完全体现该物种对尸体分解的贡献程度,故引入实际贡献力(actual contribution,AC)和实际贡献率(actual contribution rate,ACR)(本文作者编写)。实际贡献率表示不同物种的实际贡献力占所有物种的实际贡献力的比值。
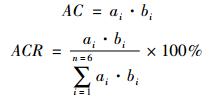
|
其中,a为本物种进食尸体时间总和;b为体质量系数,表示本物种的平均体质量(w)占所有物种的平均体质量加和的比值,即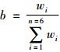
主要食腐动物的实际贡献率见表 2。野猪、亚洲黑熊、大嘴乌鸦在脊椎动物参与的大型动物尸体分解过程中占到的权重达99.9%以上。其中,大嘴乌鸦食腐时间量最大,即相对贡献力最大,但是因体型与食量限制,其实际贡献率明显低于野猪和亚洲黑熊。
| 物种 Species |
野猪 Sus scrofa |
亚洲黑熊 Ursus thibetanus |
大嘴乌鸦 Corvus macrorhynchos |
香鼬 Mustela altaica |
果子狸 Paguma larvata |
啮齿动物 Glires |
| 体质量/kg | 80 | 100 | 0.5 | 0.3 | 4.0 | 0.05 |
| 进食尸体的 时间总和/min |
984 | 82 | 6 726 | 53 | 6 | 117 |
| 实际贡献率/% | 87.15 | 9.08 | 3.72 | 0.02 | 0.03 | 0.01 |
依据统计所得数据,计算日活动强度指数(daily activity intensity index,DII),分析不同食腐动物食腐的日活动规律(武鹏峰等,2012):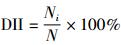
其中拍摄率最高的3种动物日活动节律见图 3。野猪全天有采食行为,在清晨与16: 00和21: 00呈现3个明显的采食高峰,以16: 00和21: 00最为明显。

|
| 图 3 不同动物的日活动强度分布 Fig. 3 Distribution of daily activity intensity of different animals |
| |
大嘴乌鸦仅在白天进食,每天有3个进食高峰,分别为08: 00、11: 00和14: 00。在12: 00停止取食。可能与正午光线过强、气温过高有关。
亚洲黑熊偏向于在夜间进食,并在20: 00左右形成一个进食高峰期,在06: 00—08: 00或16: 00左右也有进食行为。
野猪与亚洲黑熊在食腐时间上比较相似,但亚洲黑熊更倾向于避开白天的时间进食。大嘴乌鸦则为严格的昼行性进食。
3.4 部分参与尸体分解的动物红外相机照片实验期间利用红外相机陷阱技术拍摄到部分动物参与尸体分解(图 4)。

|
| 图 4 部分参与尸体分解的动物照片 Fig. 4 Part of the species involved in the decomposition of corpses |
| |
本研究发现,不同动物进食尸体的方式和行为有很大差异。
亚洲黑熊有拖动尸体的能力,但在取食过程中未发现该行为。本研究发现,第二年春季亚洲黑熊结束冬眠后,在一头因离水源过近而被人工掩埋的羚牛尸体附近长时间挖掘、进食,且进食过程中在尸体上长时间打滚,并表现出类似气味标记与掩盖的行为。
在以往研究中,黄喉貂以浆果、啮齿类甚至小型有蹄类为食(张建军,2000)。本研究发现黄喉貂不会啃食新鲜或者腐烂的动物尸体,但会取食尸体剖检过程中遗留在附近的新鲜碎肉。
有研究发现香鼬的食物由高原鼠兔Ochotona curzoniae为主的啮齿类动物组成,虽无长期固定巢穴,但雌雄均有拖动和贮存食物的行为(魏万红,1991;魏万红等,1994)。张逦嘉等(2011)通过分析粪便内容物(毛发、骨骼、鳞片等)发现香鼬的食物主要由鼠类、昆虫、植物类组成,不含有蹄类。本研究发现香鼬会啃食新鲜的有蹄类尸体,且进食时所有食物均被叼离原处,远离红外相机视野,故无法判断香鼬是否会储存尸体。同时拍摄记录也显示香鼬存在主动挖掘扩大洞口进洞猎杀因尸体腐烂滋生的啮齿动物的行为。
野猪会啃食高度腐烂的动物尸体,存在采食同类尸体的现象,且有将尸体拖离现场的行为。野猪通过拱、咬等方式将尸体拖离原位置进行食用,疑似为排斥其他竞争者,如大嘴乌鸦。
Shean(1993)用家猪尸体进行分解研究,发现蝇蛆是尸体分解的最主要因素,在用人类尸体进行分解研究中也证实了蝇蛆是尸体分解的主要因素(陈禄仕,2011)。但是本研究发现,有大量蝇蛆出现(高峰期覆盖满尸体1/2的表面积)的尸体仅有野猪一例,其他尸体虽有蝇蛆出现,但均未达到野猪尸体的蝇蛆量。之前的研究认为,猪的器官大小、行为生理与人最为接近,是人类实现异种器官移植的首选动物(兰宗宝等,2008),除研究地点可能缺乏中大型食腐动物外,人类与猪尸体生理生化结构相近可能是造成以往研究中认为蝇蛆是尸体分解的主要因素的原因之一。
研究发现,野猪尸体上蝇蛆量极大,同域分布的红腹锦鸡Chrysolophus pictus、血雉Ithaginis cruentus等鸡形目Galliformes鸟类不前往取食蝇蛆或者昆虫,食虫鸟类中仅有大嘴乌鸦前往采食蝇蛆。绿背山雀多次前往采集尸体腐败脱落的毛发。
在有尸体存在的研究区域内,果子狸会啃食高度腐烂的尸体。有蹄类动物如羚牛、小麂、斑羚等均没有接近同类尸体进行标记的行为。金丝猴、藏酋猴等灵长类动物在尸体处短暂停留,但不表现出其他行为。红外相机多次拍摄到岩松鼠,但未发现有啃食尸体的行为,其出现可能与取食尸体滋生的无脊椎动物有关。
5 讨论本研究中脊椎动物参与分解的时长累积达到7 968 min,在尸体的分解过程中起到了非常重要的作用。研究表明,野猪、亚洲黑熊、大嘴乌鸦在大型动物尸体分解过程中起到的作用最大,各权重分别为野猪87.15%、亚洲黑熊9.08%、大嘴乌鸦3.72%。大嘴乌鸦虽然参与分解的时间最长,但是考虑到体型因素,起到的作用并不如野猪和亚洲黑熊大。
与此同时,通过对照研究发现尸体的存在可以吸引也可以排斥不同的动物,这样对动物的分布格局会产生一定的影响。在保护区实际的食物链中,尸体的分解并不是物质由动物体直接转向微生物分解者的过程,而是流向了杂食类、啮齿类、食肉类等多个营养级。重达400 kg的羚牛尸体对体质量只有几十到几百克的啮齿类、鼬科Mustehdae等动物来说是一个巨大的食物资源,这与深海鲸落现象类似(Feldman et al., 1998)。尸体既为不同营养级的食肉、食腐动物提供了直接的食物来源,例如野猪、亚洲黑熊、大嘴乌鸦与啮齿动物,也间接地为鼬科、灵猫科Viverridae等小型食肉动物提供了啮齿类食物资源。同时,尸体也为多种鸟类提供了筑巢用的毛发。
对保护区内尸体分解过程的研究使我们对保护区内大型动物尸体物质再循环的过程有了新的认识,对动物尸体分解、净化过程和意义有了比较清晰的了解。研究建议:尸体是自然生态系统中不可缺少的一部分,在保护区管理过程中,除可能污染水源的尸体需要特殊处理外,其余的应让其进行自然分解,而不应该继续采用现行的深埋并喷洒石灰粉和消毒粉的方式。但鉴于研究中发现鸡形目鸟类拒绝前往尸体处进食蝇蛆,故建议保护区在开展鸡形目观测区域可以有选择地清理动物尸体。
致谢: 本研究过程中得到唐家河国家级自然保护区管理处的大力支持以及白熊坪保护站员工的密切配合。张平女士为数据处理提供了帮助。本研究过程中有来自全国各地30名志愿者的帮助,特此致谢!| 陈禄仕. 2005. 尸体腐败分解过程的观察与研究[J]. 法医学杂志, 21(4): 246–248. |
| 陈禄仕. 2011. 贵阳市郊嗜尸蝇类群落组成及生长观察[J]. 中国法医学杂志, 26(3): 204–206. |
| 陈禄仕. 2000. 嗜尸昆虫侵袭不同环境中尸体与死后经过时间的实验研究[J]. 中国法医学杂志, 3: 157–160. DOI:10.3969/j.issn.1001-5728.2000.03.013 |
| 胡锦矗. 1990. 唐家河自然保护区黑熊的觅食生态研究[J]. 四川师范学院学报(自然科学版), 11(3): 182–194. |
| 黄福贞. 1996. 分解者在生态系统物质循环中的作用[J]. 生物学通报, 31(12): 15–16. |
| 兰宗宝, 王修文, 许惠艳. 2008. 猪器官异种移植研究进展[J]. 广西农业科学, 39(3): 380–384. |
| 刘哲铭. 2016. 黑龙江省帽儿山林区小型鸟类尸体利用途径及效率分析[D]. 哈尔滨: 东北林业大学. http://cdmd.cnki.com.cn/Article/CDMD-10225-1016309572.htm |
| 彭倩宜, 叶鲁思, 马丽平, 等. 2009. 嗜尸性甲虫在尸体上的演替及其法医学应用前景[J]. 法医学杂志, 25(6): 447–450. |
| 四川省唐家河国家级自然保护区管理处, 四川省广元唐家河风景区管理局, 四川省地方志办公室. 2012. 四川唐家河国家级自然保护区志[M]. 成都: 四川科学技术出版社: 49-73. |
| 武鹏峰, 刘雪华, 蔡琼, 等. 2012. 红外相机技术在陕西观音山自然保护区兽类监测研究中的应用[J]. 兽类学报, 32(1): 67–71. |
| 魏万红. 1991. 香鼬行为生态研究[D]. 西宁: 中国科学院西北高原生物研究所. http://www.irgrid.ac.cn/handle/1471x/382180 |
| 魏万红, 周文扬, 樊乃昌. 1994. 香鼬的栖息地选择, 觅食和育幼行为[J]. 兽类学报, 14(3): 184–188. |
| 张建军. 2000. 黄喉貂生态特性的初步观察[J]. 河北林果研究, s1: 195–196. |
| 张逦嘉, 王安梦, 袁梨, 等. 2011. 内蒙古赛罕乌拉自然保护区4种小型食肉目动物的食性构成的初步分析[J]. 兽类学报, 31(1): 55–61. |
| 张建刚, 符德平. 2012. 36例腐败尸体的法医学检验及讨论[J]. 广东公安科技, 110(4): 50–54. |
| 章书声, 鲍毅新, 王艳妮, 等. 2012. 不同相机布放模式在古田山兽类资源监测中的比较[J]. 生态学杂志, 31(8): 2016–2022. |
| Feldman RA, Shank TM, Black MB, et al. 1998. Vestimentiferan on a whale fall[J]. The Biological Bulletin, 194(2): 116–119. DOI:10.2307/1543041 |
| Shean BS. 1993. Observations of differential decomposition on sun exposed v.shaded pig carrion in coastal Washington State[J]. Journal of Forensic Science, 38(4): 938–949. |
| Watson EJ, Carlton CE. 2003. Spring succession of necrophilous insects on wildlife carcasses in Louisiana[J]. Journal of Medical Entomology, 40(3): 338–347. DOI:10.1603/0022-2585-40.3.338 |