 2017, Vol. 36
2017, Vol. 36扩展功能
文章信息
- 赵伊伦, 陈梦羽, 缪宁, Timothy Moermond
- ZHAO Yilun, CHEN Mengyu, MIAO Ning, MOERMOND Timothy
- 四川大学望江校区5种常见留鸟的食性及其季节差异
- A Study on Seasonal Variation Diet of Five Most Common Resident Bird Species in Wangjiang Campus of Sichuan University, Chengdu
- 四川动物, 2017, 36(5): 576-581
- Sichuan Journal of Zoology, 2017, 36(5): 576-581
- 10.11984/j.issn.1000-7083.20160218
-
文章历史
- 收稿日期: 2016-08-12
- 接受日期: 2017-04-10






随着城市化进程的发展及人们对鸟类保护意识的增强,越来越多的鸟类在城市化环境下成功定居和适应(陈水华等,2000;张琴,2014)。城市鸟类的组成及生态特点引起了较多的关注,并吸引了一些研究者展开研究(王彦平,2003;李鹏,2009;寿艺丹,2015)。关于城市人居环境下鸟类的食性研究大多针对单一季节(王维禹,郭延蜀,2005),例如:北京市区绿化带内鸟类食源树种研究(隋金玲等,2006),柳州城市公园冬季鸟类食源树种调查研究(谭丽凤,杨昌尚,2011)等,但对城市鸟类的食物组成与季节差异研究较少(赵伟等,2007;刘彬等,2009)。
通过对成都市典型的鸟类活动区域——四川大学望江校区的鸟类取食行为与季节变化进行研究,期望探讨:1) 四川大学望江校区优势度最大的鸟类是哪几种?2) 这些留鸟能够占据优势地位的生态策略是什么?3) 它们之间的生态位是否有重叠和分化?本研究有助于了解成都市常见留鸟的食性,研究结果可为城市化的鸟类行为学和鸟类保护研究提供借鉴和参考。
1 研究方法 1.1 望江校区植物种类调查辨认望江校区内的植物种类,了解乔木和草本的各取食部位特点,如花、果、种子的形状、颜色和大小等。
1.2 鸟类取食观察调查于2014年8月—2015年8月在四川大学望江校区进行,主要集中在日出后、日落前2 h。本研究共进行53 d,每个季度约13次,即每4 d在校园内进行一次观察。观察到鸟类的取食次数共803次,其中有效数据536组(无法辨识食物类别的数据视为无效数据)。调查采用样线法,根据地形和人为活动干扰程度,在兼顾湿地、绿化带、居住区等生境,随机选取多条调查线路进行调查。调查时以1~2 km·h-1的速度慢步前进,用双筒望远镜(Olympus 10×42和Bosma 8×42) 对视线范围内的鸟类进行观察,如遇到鸟多或其他特殊情况时,则稍停留片刻。记录所见鸟的种类和数量及其取食状况。
1.3 最常见留鸟的确定统计实验中记录到的所有鸟类出现的频次(某种鸟类出现1只或1群记为1),统计各季节中每种鸟类的出现频率及总数,选取出现频率最高且数量相对较多的留鸟(留鸟为四季都出现的鸟类)作为研究中的常见留鸟(Ottoni et al., 2009)。
1.4 各种食物出现频次百分率的统计通过直接观察难以统计鸟类对每种食物取食的量,但是可以统计观察到的食物被取食的次数(Ottoni et al., 2009),在实验中观察到一次鸟类取食某种食物记为1。计算每种食物被取食的频次百分率及分别被5种常见留鸟取食的频次百分率;将食物分为植物性食物(统计植物种类及被取食类型)、动物性食物(昆虫等)、人类提供的食物(馒头、米饭等食物碎屑)和其他食物(枯叶等),计算鸟类取食某种食物占总取食次数的百分率,得出鸟类取食某种食物的频次百分率。
1.5 生态位宽度计算以Levins指数来计算不同鸟类的生态位宽度(Levin,1968):
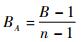
|
其中BA为Levins指数的值,B=1/Σpi2,pi为某种食物被取食占所有食物的比例(即食物的被取食频次百分率),n代表被取食次数。BA在0到1之间波动:该值越接近1代表取食生态位越宽,所有食物的取食频率相当,说明该鸟是食性广泛的物种;数值接近0代表这个物种专性取食一种或几种食物,说明该鸟是对食物专一的物种(Ottoni et al., 2009)。
1.6 生态位重叠分析使用相似性百分率指数计算5种常见留鸟的生态位重叠:
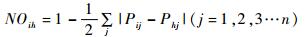
|
其中,NOih为种i和种h的生态位重叠值,Pij和Phj分别为种i和种h在资源状态j中的个体数占该种在群落中全部个体数的百分率,即分别取食某种食物的次数分别占总取食次数的百分率。相似性百分率可以反映不同留鸟之间食性生态位的重叠程度,值越大表明2种鸟的食性和行为越相似。
2 结果与分析 2.1 常见留鸟的基本情况共调查到鸟类19科32属42种,其中5种留鸟的数量和出现频次百分率最大,占总数的81.23%,分别为白颊噪鹛Garrulax sannio(20.14%)、麻雀Passer montanus(16.58%)、白头鹎Pycnonotus sinensis(16.30%)、珠颈斑鸠Streptopelia chinensis(15.21%)和乌鸫Turdus merula(13.01%)(表 1)。
| 物种Species | 春Spring | 夏Summer | 秋Autumn | 冬Winter | |||||||||||
| 频次 Time |
频率 Frequency/% |
总数 Number |
频次 Time |
频率 Frequency/% |
总数 Number |
频次 Time |
频率 Frequency/% |
总数 Number |
频次 Time |
频率 Frequency/% |
总数 Number |
||||
| 麻雀Passer montanus | 45 | 20.64 | 166 | 28 | 18.06 | 135 | 15 | 12.20 | 143 | 33 | 13.41 | 308 | |||
| 白头鹎 Pycnonotus sinensis |
22 | 10.09 | 37 | 16 | 10.32 | 59 | 28 | 23.58 | 157 | 53 | 21.54 | 218 | |||
| 白颊噪鹛 Garrulax sannio |
45 | 20.64 | 104 | 31 | 20.00 | 96 | 27 | 21.95 | 102 | 44 | 17.89 | 180 | |||
| 乌鸫Turdus merula | 30 | 13.76 | 45 | 30 | 19.35 | 88 | 8 | 7.32 | 24 | 27 | 10.98 | 49 | |||
| 珠颈斑鸠 Streptopelia chinensis |
35 | 16.06 | 54 | 35 | 22.58 | 75 | 19 | 15.45 | 61 | 22 | 8.94 | 43 | |||
| 戴胜Upupa epops | 4 | 1.83 | 5 | 1 | 0.65 | 1 | — | — | — | — | — | — | |||
| 普通翠鸟Alcedo atthis | 1 | 0.46 | 1 | 1 | 0.65 | 1 | 1 | 0.81 | 1 | — | — | — | |||
| 蚁Jynx torquilla | — | — | — | — | — | — | — | — | — | 2 | 0.81 | 2 | |||
| 棕腹啄木鸟 Dendrocopos hyperythrus |
— | — | — | — | — | — | 1 | 0.81 | 1 | — | — | — | |||
| 棕背伯劳Lanius schach | — | — | — | — | — | — | — | — | — | 2 | 0.81 | 3 | |||
| 喜鹊Pica pica | 1 | 0.46 | 1 | — | — | — | — | — | — | — | — | — | |||
| 红嘴蓝鹊 Urocissa erythrorhyncha |
— | — | — | — | — | — | 1 | 0.81 | 1 | — | — | — | |||
| 红耳鹎 Pycnonotus jocosus |
— | — | — | — | — | — | 1 | 0.81 | 1 | 2 | 0.81 | 3 | |||
| 棕腹大仙鹟 Niltava davidi |
— | — | — | — | — | — | — | — | — | 3 | 1.22 | 3 | |||
| 棕腹仙鹟 Niltava sundara |
— | — | — | — | — | — | — | — | — | 1 | 0.41 | 1 | |||
| 棕胸蓝姬鹟 Ficedula hyperythra |
— | 0.92 | 2 | — | — | — | — | — | — | — | — | — | |||
| 鹊鸲Copsychus saularis | 3 | 1.38 | 4 | 2 | 1.29 | 3 | 1 | 0.81 | 1 | 2 | 0.81 | 2 | |||
| 红胁蓝尾鸲 Tarsiger cyanurus |
4 | 1.83 | 4 | — | — | — | — | — | — | 9 | 3.66 | 10 | |||
| 金色林鸲 Tarsiger chrysaeus |
1 | 0.46 | 1 | — | — | — | — | — | — | — | — | — | |||
| 蓝额红尾鸲 Phoenicurus frontalis |
1 | 0.46 | 1 | — | — | — | — | — | — | — | — | — | |||
| 宝兴歌鸫 Turdus mupinensis |
— | — | — | — | — | — | — | — | — | 1 | 0.41 | 1 | |||
| 灰头鸫 Turdus rubrocanus |
— | — | — | — | — | — | — | — | — | 1 | 0.41 | 1 | |||
| 灰翅鸫 Turdus boulboul |
— | — | — | — | — | — | — | — | — | 1 | 0.41 | 1 | |||
| 八哥 Acridotheres cristatellus |
— | — | — | 4 | 2.59 | 5 | 3 | 2.44 | 6 | — | — | — | |||
| 丝光椋鸟 Sturnus sericeus |
— | — | — | — | — | — | 1 | 0.81 | 2 | — | — | — | |||
| 灰椋鸟 Sturnus cineraceus |
— | — | — | 3 | 1.94 | 7 | 2 | 1.63 | 6 | 8 | 3.25 | 85 | |||
| 白鹡鸰Motacilla alba | — | — | — | — | — | — | 2 | 1.63 | 2 | 1 | 0.41 | 2 | |||
| 黄腹山雀 Parus venustulus |
— | — | — | — | — | — | — | — | — | 1 | 0.41 | 2 | |||
| 大山雀Parus major | 1 | 0.46 | 1 | 1 | 0.65 | 2 | — | — | — | 1 | 0.41 | 1 | |||
| 红头长尾山雀 Aegithalos concinnus |
3 | — | 5 | 1 | 0.65 | 10 | 8 | 6.50 | 148 | 1 | 0.41 | 30 | |||
| 棕脸鹟莺 Abroscopus albogularis |
— | — | — | — | — | — | — | — | — | 1 | 0.41 | 4 | |||
| 白喉噪鹛 Garrulax albogularis |
1 | 0.46 | 2 | — | — | — | — | — | — | — | — | — | |||
| 红嘴相思鸟 Leiothrix lutea |
3 | 1.38 | 27 | — | — | — | — | — | — | 11 | 4.47 | 81 | |||
| 棕头鸦雀 Paradoxornis webbianus |
1 | 0.46 | 10 | — | — | — | — | — | — | 2 | 0.81 | 33 | |||
| 灰喉鸦雀Paradoxornisalphonsianus | 1 | 0.46 | 2 | — | — | — | — | — | — | — | — | — | |||
| 红胸啄花鸟 Dicaeum ignipectus |
— | — | — | — | — | — | — | — | — | 3 | 1.22 | 3 | |||
| 白腰文鸟 Lonchura striata |
1 | 0.46 | 3 | — | — | — | 1 | 0.81 | 4 | 2 | 0.81 | 6 | |||
| 红眉松雀 Pinicola subhimachala |
— | — | — | — | — | — | — | — | — | 1 | 0.41 | 1 | |||
| 燕雀 Fringilla montifringilla |
— | — | — | — | — | — | — | — | — | 1 | 0.41 | 3 | |||
| 金翅雀Carduelis sinica | 2 | 0.92 | 2 | 2 | 1.29 | 6 | 1 | 0.81 | 12 | 2 | 0.81 | 26 | |||
| 黑尾蜡嘴雀 Eophona migratoria |
5 | 2.75 | 16 | — | — | — | — | — | — | — | — | — | |||
| 黑头蜡嘴雀 Eophona personata |
1 | 0.46 | 1 | — | — | — | — | — | — | 6 | 2.44 | 18 | |||
5种常见留鸟的植物性食物共包含38科54属61种,其中,银木Cinnamomum septentrionale、早熟禾Poa annua、天竺桂Cinnamomum japonicum、君迁子Diospyros lotus等是常被取食的植物种类,被取食频率分别为16.79%、14.07%、9.63%、5.68%,其他植物种类占53.83%(表 2)。
| 物种Species | 频次Times | 频率Frequency/% |
| 银木Cinnamomum septentrionale | 68 | 16.79 |
| 早熟禾Poa annua | 57 | 14.07 |
| 天竺桂Cinnamomum japonicum | 39 | 9.63 |
| 君迁子Diospyros lotus | 23 | 5.68 |
| 其他 | 218 | 53.83 |
| 总计 | 592 | 100 |
鸟类对植物的果实、种子与其他部位的取食情况不同(表 3)。在5种常见留鸟中,乌鸫对果实与种子没有特别偏好,白头鹎偏好果实,珠颈斑鸠、麻雀与白颊噪鹛偏好种子。植物性食物被取食频率最高的部分为种子(55.33%)。
| 物种Species | 果实Fruit | 种子Seed | 其他Others |
| 白颊噪鹛Garrulax sannio | 166 | 214 | 16 |
| 白头鹎Pycnonotus sinensis | 234 | 33 | 27 |
| 麻雀Passer montanus | 76 | 452 | 54 |
| 乌鸫Turdus merula | 42 | 40 | 0 |
| 珠颈斑鸠 Streptopelia chinensis |
74 | 128 | 11 |
| 总计 | 592 | 867 | 108 |
| 频率/% | 37.78 | 55.33 | 6.89 |
在5种常见留鸟中,白颊噪鹛的Levins指数(0.315) 小于其他留鸟,且标准差最小(0.127),说明其生态位较窄且相对稳定。乌鸫在春季的Levins指数最大(0.71),夏季、秋季和冬季指数最大的分别为白头鹎(0.91)、珠颈斑鸠(1.00) 和麻雀(0.83)。白头鹎、麻雀和珠颈斑鸠在春季的Levins指数为各季最低,说明这3种鸟在春季的食性生态位较窄。白颊噪鹛和乌鸫的Levins指数在夏季最低,说明这2种鸟类在夏季的食性生态位最窄(表 4)。
| 白颊噪鹛 Garrulax sannio |
白头鹎 Pycnonotus sinensis |
麻雀 Passerm-ontanus |
乌鸫 Turdus merula |
珠颈斑鸠 Streptopelia chinensis |
|
| 春季 | 0.32 | 0.27 | 0.27 | 0.71 | 0.21 |
| 夏季 | 0.14 | 0.91 | 0.43 | 0.15 | 0.39 |
| 秋季 | 0.44 | 0.77 | 0.71 | 0.64 | 1.00 |
| 冬季 | 0.36 | 0.43 | 0.83 | 0.64 | 0.65 |
| 均值 | 0.315±0.127 | 0.595±0.296 | 0.560±0.266 | 0.535±0.259 | 0.563±0.343 |
相似性百分率指数最小的为麻雀与白颊噪鹛(49.7%),说明这2种鸟的食性差异最大,也表明这2种鸟的生态位分化较大。该指数最大的值为乌鸫与麻雀(79.9%),表明它们取食的对象、活动的时间和空间范围较一致(表 5)。
| 白颊噪鹛 | 白头鹎 | 麻雀 | 乌鸫 | 珠颈斑鸠 | |
| 白颊噪鹛 | — | ||||
| 白头鹎 | 52.9% | — | |||
| 麻雀 | 49.7% | 70.7% | — | ||
| 乌鸫 | 60.1% | 61.9% | 79.9% | — | |
| 珠颈斑鸠 | 76.0% | 67.0% | 65.8% | 64.9% | — |
四川大学望江校区有较丰富的植物种类(白洁,2011),为鸟类在不同季节提供了丰富的食物来源。校园内四季都有不同种类的植物提供果实或种子,比如春季的早熟禾与棒头草Polypogon fugax、夏季的狗牙根Cynodon dactylon、秋季的君迁子、冬季的小叶榕Ficus concinna。丰富的食物为鸟类的定居和繁衍提供基本的保障。5种常见留鸟的食性均较广泛,即使在不同季节对部分食物有所偏好,也会取食其他食物,这是它们成功定居并成为优势种的原因之一。这样的环境适应性有利于它们在人类活动较多的校园内获得更多的活动空间和食物,也较容易在人类活动区域获得额外的投食(王维禹,郭延蜀,2005;隋金玲等,2006;谭丽凤,杨昌尚,2011)。过去的研究认为,本地鸟类的种群密度、丰富度与植被的组成和覆盖程度密切相关(Mills et al., 1989),而现今城市化环境导致的环境均质性使杂食性和食果鸟类易成为环境的适应者(McKinney,2006)。表现为部分鸟类逐渐适应城市化的人居环境并成功定居和繁殖(隋金玲等,2006;Ottoni et al., 2009;Aronson et al., 2014),这也是城市化环境中鸟类种类日趋丰富并共存的重要原因。
一般而言,物种共存是由物种间的时间、空间和资源生态位的分化及其权衡所决定(Calcagno et al., 2006)。城市化环境下四川大学望江校区的鸟类以植物性食物为主,动物性食物和人类提供的食物为辅。随着季节的变化,这些优势留鸟的食物组成及其食性生态位变化明显。在望江校区中,这5种优势留鸟的出现频率占所有鸟类出现频率的79.91%,说明其数量占绝对优势,但这也意味着城市环境对鸟类种类影响很大,单一和均质的环境加上各种人类干扰减少了生态系统的初级生产力进而影响鸟类的物种数量(Lee et al., 2004),特别是稀有鸟类的生存(Aronson et al., 2014)。5种常见留鸟的食性都较为广泛,且重叠程度较大,但是它们的取食种类、活动区域与活动时间有一定差异。Levins指数分析表明,不同留鸟的食性生态位在各个季节都有差异。四川大学望江校区中5种常见的优势留鸟都是杂食性或者食果鸟类,这与McKinney(2006)的研究结果一致,这5种留鸟食性广泛且有取食时间和空间的分化,这是它们成为优势留鸟并共存的原因。
本研究通过对四川大学望江校区鸟类的取食行为与食性的季节变化进行研究,发现四川大学望江校区最常见的留鸟为白颊噪鹛、白头鹎、麻雀、乌鸫和珠颈斑鸠,它们都是杂食性鸟类,生态位存在重叠和分化。本研究是一个基于现状的描述和分析,而城市化环境下的鸟类组成及其动态,以及这些鸟类适应城市环境的生理生态学方面的机制都值得进一步研究和探索。从保护城市鸟类多样性的角度来看,在城市规划中,应尽可能保持多样化的植物和植被种类,这样既可以提供景观的多样性,也可以为潜在的城市鸟类定居者提供多样化的食物来源和栖息地。
致谢: 感谢参与并协助调查的陈俊橙、郭艳、梁思琪、张馨月和郑歆蕾同学!| 白洁. 2011. 四川大学校园植物现状调查与分析[J]. 四川林业科技, 32(2): 60–69. |
| 陈水华, 丁平, 郑光美, 等. 2000. 城市化对杭州市湿地水鸟群落的影响研究[J]. 动物学研究, 21(4): 279–285. |
| 李鹏. 2009. 杭州市鸟类区系与城市化对鸟类群落的影响[D]. 杭州: 浙江大学. http://cdmd.cnki.com.cn/Article/CDMD-10335-1011067079.htm |
| 刘彬, 周立志, 汪文革, 等. 2009. 大别山山地次生林鸟类群落集团结构的季节变化[J]. 动物学研究, 30(3): 277–287. |
| 寿艺丹. 2015. 上海城乡梯度上公园绿地鸟类物种多样性格局[D]. 上海: 华东师范大学. http://cdmd.cnki.com.cn/Article/CDMD-10269-1015345613.htm |
| 隋金玲, 张志翔, 胡德夫, 等. 2006. 北京市区绿化带内鸟类食源树种研究[J]. 林业科学, 42(12): 83–89. DOI:10.3321/j.issn:1001-7488.2006.12.015 |
| 谭丽凤, 杨昌尚. 2011. 柳州城市公园冬季鸟类食源树种调查研究[J]. 安徽农业科学, 38(34): 19428–19430. |
| 王维禹, 郭延蜀. 2005. 白头鹎春季食性及取食空间生态位的初步研究[J]. 四川动物, 24(4): 466–468. |
| 王彦平. 2003. 鸟类对城市化的适应性研究[D]. 杭州: 浙江大学. http://cdmd.cnki.com.cn/article/cdmd-10335-2003102296.htm |
| 徐玉梅. 2009. 乌鸫繁殖习性及食性的初步研究[J]. 生物学通报, 44(3): 31–33. |
| 张琴. 2014. 白头鹎生活史特征对城市化的响应[D]. 杭州: 浙江师范大学. http://cpfd.cnki.com.cn/article/cpfdtotal-zjkx201311002067.htm |
| 赵伟, 宋森, 邵明勤, 等. 2007. 甘肃民勤治沙站纵纹腹小鸮食性的季节变化[J]. 动物学报, 53(6): 953–958. |
| Aronson MF, LaSorte FA, Nilon CH, et al. 2014. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic driver[J]. Procedings Biological Sciences, 281(1780): 20133330. DOI:10.1098/rspb.2013.3330 |
| Calcagno V, Mouquet N, Jarne P, et al. 2006. Coexistence in a metacommunity:the competition-colonization trade-off is not dead[J]. Ecology Letters, 9(8): 897–907. DOI:10.1111/ele.2006.9.issue-8 |
| Lee PF, Ding TS, Hsu FH, et al. 2004. Breeding bird species richness in Taiwan:distribution on gradients of elevation, primary productivity and urbanization[J]. Journal of Biogeography, 31(2): 307–314. DOI:10.1046/j.0305-0270.2003.00988.x |
| Levin R. 1968. Evolution of diversity, efficiency, and community stability[J]. American Zoology, 10(1): 17–25. |
| McKinney ML. 2006. Urbanization as a major cause of biotic homogenization[J]. Biological Conservation, 127(3): 247–260. DOI:10.1016/j.biocon.2005.09.005 |
| Mills GS, Dunning JB, Bates JM. 1989. Effects of urbanization on breeding bird community structure in southwestern desert habitats[J]. Condor, 91(2): 416–428. DOI:10.2307/1368320 |
| Ottoni I, de Oliveira FFR, Young RJ. 2009. Estimating the diet of urban birds:the problems of anthropogenic food and food digestibility[J]. Applied Animal Behavior Science, 117(1-2): 42–46. DOI:10.1016/j.applanim.2008.11.002 |