 2016, Vol. 35
2016, Vol. 35扩展功能
文章信息
- 王玲, 丁志锋, 胡君梅, 尹五元, 胡慧建
- WANG Ling, DING Zhifeng, HU Junmei, YIN Wuyuan, HU Huijian
- 广州城市绿地中鸟类对食源树种的偏好
- Bird Preference to Food Source Trees in Urban Green Space, Guangzhou, China
- 四川动物, 2016, 35(6): 838-844
- Sichuan Journal of Zoology, 2016, 35(6): 838-844
- 10.11984/j.issn.1000-7083.20160118
-
文章历史
- 收稿日期: 2016-05-09
- 接受日期: 2016-07-20






2. 广东省生物资源应用研究所, 广州 510260;
3. 广东省野生动物保护与利用公共实验室, 广州 510260;
4. 中南林业科技大学野生动植物保护研究所, 长沙 410004
2. Guangdong Institute of Applied Biological Resources, Guangzhou 510260, China;
3. Guangdong Public Laboratory of Wild Animal Conservation and Utilization, Guangzhou 510260, China;
4. Institute of Wildlife Conservation, Central South University of Forestry and Technology, Changsha 410004, China
世界范围内急剧的城市化进程所带来的生态问题,尤其是城市化对鸟类的影响引起了生态学家越来越多的关注(易兰等,2007)。城市化被认为是生物多样性丧失的主要原因(McKinney,2008),城市化带来的生态过程,如食物资源改变、人类干扰等,对鸟类群落结构产生了影响,导致城市生态系统失衡,呈现病态、无法自我调节的状态,将可能影响人类的可持续发展(Ren et al.,2007)。了解城市化对生物多样性的影响过程及其机理,提出有效的对策以减缓物种丧失速度和保护生物多样性,成为生态学家和保护生物学家高度关注的问题之一(毛齐正等,2013)。
绿地建设是现代城市建设的重要内容,是保护、恢复和提高城市生物多样性的主要举措。目前城市绿地建设常以大面积的草皮种植、乔木大树栽植等方式增加城市绿化量,运用彩色灌木、绿篱形成各种艳丽图案丰富色彩,香料植物沁人心脾,但还不太重视对食源树种的运用(姚月华,颜玉娟,2014)。随着国家大力推进“生态文明 美丽中国”建设,城市野生动物保护和恢复特别是城市鸟类的恢复成为其中的重要内容,这不仅是城市可持续发展的基本前提,也是城市生态系统稳定有序的保障。因此,城市绿地中食源树种的配置及应用对于促进城市鸟类的保护和恢复有重要的作用。
广州城市绿化率超过40%,在全国属领先水平:城市绿地纵横交错,各种居民公园、校园绿地、居住区绿地、交通绿地、生产防护绿地等随处可见(张钧挥,2011),为鸟类的生存繁衍提供了重要的前提条件。但在各类型的城市绿地中,鸟类食源树种的分布和鸟类群落的变化规律却少有关注。2014年10月—2015年9月对广州城市绿地中的鸟类食源树种及鸟类进行了调查,以期为广州城市鸟类的保护与恢复提供理论依据。
1 研究地概况广东省广州市(112°57′~114°03′E,22°26′~23°56′N)地处珠江三角洲北部,西江、北江、东江在此汇流入海,属南亚热带海洋性季风气候带,以春(3—5月)、夏(6—8月)、秋(9—11月)、冬(12月、1—2月)划分四季,年均气温22.8 ℃,1月最低,7—8月最高;雨水资源丰富,年均降水量1 800 mm(熊咏梅等,2013)。
根据绿地面积、生境类型、人为干扰等因素,选取广州市区具有代表性的7个城市绿地进行调查,即白云区麓湖公园、越秀区流花湖公园、荔湾区荔湾湖公园、天河区珠江公园、番禺区大学城湾咀头湿地公园、海珠区海珠湖公园和中山大学南校区绿地。
2 研究方法 2.1 鸟类调查采用样线法进行调查,每个绿地选取1~2条样线,共计9条,每条样线长3 km。2014年10月—2015年9月,对每条样线进行逐月调查,使用Bushnell 8×42倍双筒望远镜、数码照相机(Cannon,SX30IS)记录样线两侧25 m范围内出现及鸣叫的鸟类种类和数量,但不记录与行进同方向飞行的鸟类;同时对鸟类食性进行统计分析。调查时间为07∶ 00—10∶ 00(步行速度为1.5~2.0 km·h-1),仅在天气良好的情况下进行。
2.2 植被调查采用样方法对植物进行调查,样方数量根据绿地面积来确定,共设置24个20 m×20 m的样方,记录样方内植物和鸟类食源树种的种类和数量(张云霞等,2003)。其中,确定食源树种的主要原则是:1)野外直接观察鸟类取食;2)对鸟类粪便进行观察分析;3)参考有关资料(华南濒危动物研究所,1991;隋金玲等,2006a,2006b)中的方法。
2.3 数据处理优势度采用Howes(1989)的方法,按照鸟类不同种群数量占鸟类统计总数的百分比(Y)来确定优势种:Y≥10%为优势种(+++),1%<Y≤10%为常见种(++),0.1%<Y≤1%为稀有种(+),Y≤0.1%为罕见种(±)。
物种多样性采用Shannon-Wiener指数:H′=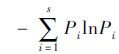
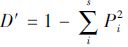
物种均匀度采用Pielou均匀度指数:J=H′/Hmax,式中,Hmax=lnS,Hmax为理论上最大多样性指数值。
物种丰富度采用Margalef丰富度指数(Margalef,1968):D=(S-1/lnN,其中,N为所有物种数之和。
采用卡方适合度检验按月统计鸟类多度和鸟类丰富度在全年的分布和变化情况(邓文洪,高玮,2005);使用单样本t检验分析鸟类多样性在全年的分布和变化情况。
3 结果 3.1 植物种类和食源树种共记录植物77科154属205种(含变种),其中,被子植物190种,裸子植物15种;乔木125种,灌木45种,草本34种,藤本1种。从生长型来看,乔木125种,占总种数的60.98%,其次是灌木(45种)、草本(34种)、藤本(1种)。桃金娘科、桑科、棕榈科植物最多,均为11种,其次是木兰科和樟科,分别为7种和6种。
能直接或间接为鸟类提供食物的食源树种有133种,它们的果实多数为蒴果、浆果和荚果。最多的是桃金娘科、桑科、棕榈科,均为11种,占总数的14%,其他均较少。鸟类取食的部位主要为嫩芽、嫩叶、花、花蜜、果实以及植物上面的昆虫。绿地中最常见、鸟类最喜欢取食的食源树种有18种,包括乔木13种:樟树Cinnamomum bodinieri、大花紫薇Lagerstroemia speciosa、白千层Melaleuca leucadendron、蒲桃Syzygium jambos、木棉Bombax malabaricum、刺桐Erythrina variegata、凤凰木Delonix regia、台湾相思Acacia confusa、羊蹄甲Bauhinia purpurea、小叶榕Ficus concinna、黄葛树Ficus virens var. sublanceolata、构树Brousso netia papyrifera、垂叶榕Ficus benjamina;灌木5种:海桐Pittosporum tobira、红千层Callistemon rigidus、朱槿Hibiscus rosa-sinensis、杜鹃Rhododendron simsii、木犀Osmanthus fragrans(表 1)。
| 植物物种 | 花期 | 果期 | 出现的绿地数 | 配置方式 |
| 樟树Cinnamomum bodinieri | 4—5月 | 8月—翌年2月 | 3 | SP |
| 大花紫薇Lagerstroemia speciosa | 5—7月 | 10—11月 | 7 | CP/LP |
| 白千层Melaleuca leucadendron | 全年 | 全年 | 5 | LP |
| 蒲桃Syzygium jambos | 3—4月 | 5—6月 | 4 | SP/CP |
| 木棉Bombax malabaricum | 3—5月 | 6—8月 | 7 | SP/LP |
| 刺桐Erythrina variegata | 3月 | 8月 | 4 | SP |
| 凤凰木Delonix regia | 6—7月 | 8—10月 | 3 | SP |
| 台湾相思Acacia confusa | 3—10月 | 8—12月 | 3 | SP |
| 羊蹄甲Bauhinia purpurea | 9—11月 | 2—3月 | 7 | SP/G |
| 小叶榕Ficus concinna | 3—6月 | 3—6月 | 7 | SP/G |
| 黄葛树Ficus virens var.sublanceolata | 4—7月 | 4—8月 | 4 | SP |
| 构树Broussonetia papyrifera | 4—5月 | 6—7月 | 5 | SP/GP |
| 垂叶榕Ficus benjamina | 8—11月 | 8—11月 | 5 | SP/LP |
| 海桐Pittosporum tobira | 5月 | 10月 | 5 | GP/H |
| 红千层Callistemon rigidus | 6—8月 | 6—8月 | 5 | CP |
| 朱槿Hibiscus rosa-sinensis | 全年 | 全年 | 5 | GP/H |
| 杜鹃Rhododendron simsii | 4—5月 | 6—8月 | 4 | GP/LP |
| 木犀Osmanthus fragrans | 9—10月 | 3月 | 4 | GP/H |
| 注: 1.物候信息参照《中国植物志》(吴征镒等,2004)并结合实际观察; 2.配置方式: SP.孤植,CP.丛植,LP.列植,GP.群植,H.绿篱。 | ||||
| Notes: 1.Phenological information of eighteen food source trees for birds was based onFlora of China (Wu et al.,2004) and field survey; 2.Configuration model: SP. solitary plant,CP. clump planting,LP. linear planting,GP. group planting,H. hedge. | ||||
共记录鸟类15 348只次,隶属于14目36科92种。其中,雀形目53种,鹳形目9种,雁形目7种,鸻形目4种,鸽形目、鹃形目和佛法僧目均为3种,鹤形目、雨燕目和形目均为2种,目、鹈形目、隼形目和鸡形目均为1种。按Howes优势度指数计算,并将水鸟去除,本次调查中共有16种林鸟优势种,按食性(尚玉昌,2007)划分为杂食性(8种)、植食性(1种)、食虫性(6种)和食肉性(1种)。为了便于统计鸟类的取食树种,未将食肉性的棕背伯劳Lanius schach纳入其中。各鸟类主要的取食树种各有不同,但杂食性鸟类取食的树种较食虫性和植食性的多(表 2)。
| 优势种 | 取食习性 | 主要取食树种 |
| 暗绿绣眼鸟Zosterops japonicus | Po | 小叶榕、羊蹄甲、大花紫薇、垂叶榕、凤凰木、蒲桃、台湾相思 |
| 白头鹎Pycnonotus sinensis | Po | 小叶榕、羊蹄甲、木棉、黄葛树、构树、樟树、大花紫薇、白千层 |
| 红耳鹎Pycnonotus jocosus | Po | 小叶榕、羊蹄甲、樟树、台湾相思、黄葛树、构树、大花紫薇、木棉 |
| 乌鸫Turdus merula | Po | 小叶榕、樟树、黄葛树、白千层、木棉、构树 |
| 白喉红臀鹎Pycnonotus aurigaste | Po | 小叶榕、樟树、羊蹄甲、白千层 |
| 鹊鸲Copsychus saularis | In | 小叶榕、黄葛树、樟树、大花紫薇 |
| 麻雀Passer montanus | Po | 小叶榕、樟树、凤凰木 |
| 长尾缝叶莺Orthotomus sutorius | In | 小叶榕、大花紫薇、羊蹄甲、垂叶榕 |
| 大山雀Parus major | In | 台湾相思、小叶榕、大花紫薇 |
| 白鹡鸰Motacilla alba | In | 台湾相思、垂叶榕 |
| 褐头鹪莺Prinia inornata | In | 小叶榕、羊蹄甲、大花紫薇 |
| 珠颈斑鸠Streptopelia chinensis | Po | 樟树、构树、柠檬桉 |
| 黄腹鹪莺Prinia flaviventris | In | 黄葛树、蒲桃、白千层 |
| 叉尾太阳鸟Aethopyga christinae | Ph | 羊蹄甲、朱槿、朱缨花、红千层、大花紫薇 |
| 红嘴蓝鹊Urocissa erythrorhyncha | Po | 长芒杜英、木棉、小叶榕、台湾相思、木麻黄 |
| 注: Po. 杂食性,Ph. 植食性,In. 食虫性。 | ||
| Notes: Po. polyphagia,Ph. phytophagous,In. insectivorous. | ||
鸟类对食源树种具有明显的偏好性,如叉尾太阳鸟Aethopyga christinae偏好在花朵艳丽的朱槿、大花紫薇、朱缨花Calliandra haematocephala上取食花蜜;而红嘴蓝鹊Urocissa erythrorhyncha则喜好在叶 片颜色较深的长芒杜英Elaeocarpus apiculatus上面 栖息。共记录暗绿绣眼鸟Zosterops japonicus出现363次,其中,小叶榕上出现67次(18.46%),羊蹄甲上出现50次(13.77%),其余树种皆为50次以下;共记录白头鹎Pycnonotus sinensis出现373次,其中,羊蹄甲上出现26次(6.97%),小叶榕上出现70次(18.82%);其他鸟类出现的频次见表 3。
| 优势种 | 出现频次/次 | ||||||||||||
| 羊蹄甲 | 小叶榕 | 大花紫薇 | 蒲桃 | 红千层 | 台湾相思 | 樟树 | 垂叶榕 | 白千层 | 木棉 | 凤凰木 | 黄葛树 | 构树 | |
| 暗绿绣眼鸟Zosterops japonicus | 50 | 67 | 20 | 14 | 12 | 13 | 13 | 15 | 12 | 10 | 17 | 6 | 2 |
| 白头鹎Pycnonotus sinensis | 26 | 70 | 17 | 2 | 2 | 6 | 18 | 12 | 10 | 20 | 7 | 25 | 19 |
| 红耳鹎Pycnonotus jocosus | 26 | 35 | 11 | 3 | 1 | 20 | 22 | 6 | 8 | 12 | 6 | 17 | 12 |
| 乌鸫Turdus merula | 7 | 38 | 2 | 5 | 0 | 9 | 36 | 6 | 17 | 15 | 9 | 18 | 10 |
| 白喉红臀鹎Pycnonotus aurigaste | 6 | 25 | 1 | 1 | 0 | 3 | 10 | 1 | 8 | 3 | 2 | 3 | 1 |
| 鹊鸲Copsychus saularis | 4 | 16 | 6 | 2 | 0 | 3 | 9 | 6 | 6 | 3 | 2 | 10 | 5 |
| 麻雀Passer montanus | 0 | 2 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 0 | 2 | 0 | 0 |
| 长尾缝叶莺Orthotomus sutorius | 4 | 11 | 6 | 0 | 0 | 3 | 1 | 4 | 2 | 0 | 0 | 1 | 1 |
| 大山雀Parus major | 5 | 8 | 7 | 4 | 2 | 10 | 6 | 3 | 2 | 3 | 5 | 2 | 0 |
| 白鹡鸰Motacilla alba | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 褐头鹪莺Prinia inornata | 4 | 8 | 3 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 3 | 0 |
| 珠颈斑鸠Streptopelia chinensis | 1 | 3 | 0 | 0 | 0 | 1 | 9 | 0 | 0 | 1 | 0 | 0 | 8 |
| 棕背伯劳Lanius schach | 3 | 10 | 3 | 0 | 0 | 1 | 3 | 1 | 2 | 3 | 6 | 2 | 5 |
| 黄腹鹪莺Prinia flaviventris | 2 | 1 | 1 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 1 |
| 叉尾太阳鸟Aethopyga christinae | 25 | 2 | 6 | 2 | 0 | 2 | 1 | 3 | 0 | 1 | 0 | 0 | 0 |
| 红嘴蓝鹊Urocissa erythrorhyncha | 3 | 6 | 0 | 0 | 0 | 6 | 1 | 0 | 1 | 10 | 0 | 3 | 1 |
| 注: 各行中最大值以加粗字体显示; 下表同。 | |||||||||||||
| Notes: Numbers in bold mean the maximum in each row; the same below. | |||||||||||||
共记录暗绿绣眼鸟3 165只次,其中,在小叶榕上出现430只次(13.58%),在羊蹄甲上出现226只次(7.14%),在大花紫薇上出现149只次(4.71%),在蒲桃、台湾相思上分别为113只次和106只次(3.57%和3.35%),其余树种皆为100只次以下;共记录白头鹎2 240只次,其中在小叶榕上出现298只次(13.30%),在黄葛树上出现101只次(4.51%),其余树种皆为100只次以下;红耳鹎P. jocosus和乌鸫Turdus merula在小叶榕上出现分别为137只次和136只次,其他鸟类均在100只次以下(表 4)。
| 优势种 | 数量分布/只次 | ||||||||||||
| 羊蹄甲 | 小叶榕 | 大花紫薇 | 蒲桃 | 红千层 | 台湾相思 | 樟树 | 垂叶榕 | 白千层 | 木棉 | 凤凰木 | 黄葛树 | 构树 | |
| 暗绿绣眼鸟Zosterops japonicus | 226 | 430 | 149 | 113 | 89 | 106 | 63 | 65 | 91 | 89 | 59 | 36 | 10 |
| 白头鹎Pycnonotus sinensis | 99 | 298 | 87 | 10 | 19 | 20 | 82 | 54 | 32 | 68 | 22 | 101 | 84 |
| 红耳鹎Pycnonotus jocosus | 100 | 137 | 49 | 14 | 2 | 85 | 78 | 19 | 31 | 59 | 18 | 60 | 51 |
| 乌鸫Turdus merula | 27 | 136 | 6 | 21 | 0 | 29 | 99 | 17 | 58 | 45 | 29 | 53 | 41 |
| 白喉红臀鹎Pycnonotus aurigaste | 17 | 98 | 2 | 5 | 0 | 13 | 33 | 2 | 31 | 16 | 10 | 12 | 2 |
| 鹊鸲Copsychus saularis | 11 | 26 | 10 | 5 | 0 | 6 | 20 | 10 | 12 | 5 | 3 | 16 | 7 |
| 麻雀Passer montanus | 0 | 6 | 0 | 1 | 0 | 0 | 9 | 0 | 0 | 0 | 13 | 0 | 0 |
| 长尾缝叶莺Orthotomus sutorius | 11 | 23 | 13 | 0 | 0 | 4 | 1 | 5 | 2 | 0 | 0 | 1 | 1 |
| 大山雀Parus major | 10 | 12 | 17 | 11 | 5 | 16 | 14 | 10 | 3 | 6 | 16 | 3 | 0 |
| 白鹡鸰Motacilla alba | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 |
| 褐头鹪莺Prinia inornata | 7 | 20 | 3 | 0 | 0 | 1 | 0 | 1 | 2 | 0 | 0 | 5 | 0 |
| 珠颈斑鸠Streptopelia chinensis | 2 | 8 | 0 | 0 | 0 | 2 | 21 | 0 | 0 | 1 | 0 | 0 | 31 |
| 棕背伯劳Lanius schach | 6 | 24 | 4 | 0 | 0 | 2 | 9 | 1 | 2 | 7 | 14 | 6 | 9 |
| 黄腹鹪莺Prinia flaviventris | 5 | 1 | 3 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 2 |
| 叉尾太阳鸟Aethopyga christinae | 50 | 3 | 8 | 2 | 0 | 2 | 1 | 6 | 0 | 1 | 0 | 0 | 0 |
| 红嘴蓝鹊Urocissa erythrorhyncha | 5 | 9 | 0 | 0 | 0 | 9 | 3 | 0 | 2 | 17 | 0 | 4 | 2 |
不同月份鸟类物种丰富度和多样性(Shannon-Wiener指数)的差异均有高度统计学意义(t=37.745,df=11,P<0.001;t=121.105,df=11,P<0.001),其中鸟类丰富度最高为3月(52种),最低为9月(39种)(图 1);鸟类多样性最高为4月(3.15),最低为9月(2.82)。

|
| 图 1 广州城市绿地中全年鸟类物种丰富度的变化 Fig. 1 Annual variation of bird species richness in urban green spaces,Guangzhou |
| |
不同月份鸟类多度差异有高度统计学意义(t=21.079,df=11,P<0.001),最高为4月(1 599只次),最低为7月(978只次)(图 2)。

|
| 图 2 广州城市绿地中全年鸟类多度的变化 Fig. 2 Annual variation of bird species abundance in urban green spaces,Guangzhou |
| |
不同城市绿地中鸟类多样性指数差异较大。海珠湖公园的Shannon-Wiener指数和Pielou均匀度指数最高,珠江公园的Shannon-Wiener指数最低,中山大学南校区绿地的Pielou均匀度指数最低;大学城湾咀头湿地公园的Margalef丰富度指数最高,荔湾湖公园和珠江公园的均为最低(表 5)。
| 调查区 | Shannon-Wiener指数(H′) | Pielou均匀度指数(J) | Margalef丰富度指数(D) |
| 麓湖公园 | 2.881 | 0.736 | 10.84 |
| 流花湖公园 | 2.556 | 0.686 | 9.07 |
| 荔湾湖公园 | 2.395 | 0.735 | 5.53 |
| 珠江公园 | 2.345 | 0.720 | 5.53 |
| 大学城湾咀头湿地公园 | 2.845 | 0.717 | 11.50 |
| 海珠湖公园 | 3.092 | 0.786 | 11.06 |
| 中山大学南校区 | 2.378 | 0.640 | 8.85 |
| 注: 加粗字体为各列最大值。 | |||
| Note: Bold numbers represent the maximum in each column. | |||
本次调查共记录食源树种133种,常见的仅18种,表明广州城市绿地中食源树种的空间分布不均匀,导致各个绿地中鸟类的多度、物种丰富度、物种多样性差异较大。在常见的18种食源树种中,大花紫薇、木棉、羊蹄甲和小叶榕在每个绿地中都有分布,它们不仅具有较好的绿化价值,同时也吸引了部分鸟类,因此,各城市绿地具有部分相同的食源树种优势种,其成为鸟类群落的重要组成部分。
4.2 鸟类优势种取食习性及主要取食树种食性和主要取食的树种对于鸟类的分布有较大影响。从生物链上看,植物既能为昆虫提供食物,也能为鸟类提供食物,同时昆虫又是食虫性鸟类的食物,因此,植物是鸟类的最终食物来源(郑光美,1995)。随着植物物候期的变化,鸟类的多度和物种丰富度也有较大变化,这是因为植物的花期主要集中在春季,果期主要在秋季,使得鸟类物种丰富度的2个峰值出现在春、秋季,这与食物可获得性决定鸟类分布有关(Ferger et al.,2014):春季Shannon-Wiener指数最高,植物的花和芽极大地满足了鸟类对食物的需求(葛振鸣等,2005);夏季天气炎热,鸟类食源相对匮乏,因此观察到的鸟类物种和数量都较春季少;秋季植物果实、种子成熟,食源大大增加,因此鸟类数量达到高峰;冬季只有极少的食源树种还挂果,且大部分的树叶开始凋落,因此Shannon-Wiener指数较低。
4.3 鸟类对食源树种的偏好本研究中鸟类最主要的食源树种有羊蹄甲、大花紫薇、小叶榕、木棉、台湾相思等。这类树种具有能吸引鸟类的特征,如木棉和台湾相思具有高大的树形、较大的树冠,羊蹄甲、大花紫薇、小叶榕具有较长的花果期,能为鸟类提供良好的栖息环境和躲避敌害的场所。一方面植被盖度影响鸟类的分布(陆祎玮等,2007),同时较高的树木提供了更好的安全性,有利于鸟类避开地面上的车辆和人流的干扰。此外,这几种食源树种花型大、气味芬芳,可吸引多种昆虫觅食,也是鸟类取食昆虫的良好场所(张晓明等,2015);同时,这些树种的果实多为浆果、梨果和蒴果,都是鸟类偏爱取食的类型。因此,广州城市鸟类倾向于选择这一类树种作为栖息繁衍的场所。
4.4 鸟类群落的时空变化番禺区大学城湾咀头湿地公园鸟类的Shannon-Wiener指数最高,这与该地区拥有大面积水域和丰富的无脊椎动物、鱼、虾、贝类等有关(Horváth,2012),其为鹭科和雁鸭类提供了适宜的栖息地和丰富的食物;岸堤的常绿阔叶林植物和灌丛亦为鹎科、椋鸟科等林鸟提供了良好的栖息环境。
人为干扰和生境异质性也是影响鸟类群落结构的重要因子(李敏等,2012;邓娇等,2014)。荔湾区荔湾湖公园和天河区珠江公园人流量大、植被单一,鸟类丰富度较低(26种),Shannon-Wiener指数和Margalef丰富度指数也较低。中山大学南校区为校园环境,植物种类较丰富,但由于该区域接近居民区,人为干扰强烈,因此该区域Pielou均匀度指数最低。
4.5 建议为保护和吸引更多鸟类,我们建议:(1)常见的食源树种仅18种,空间分布不均匀,因此在植物配置时,需增加食源树种的数量,提高食源植物配置的均匀度;(2)城市绿地中配置的植物应考虑其对鸟类招引的生态学效应,如在食物资源匮乏的夏季和冬季,应增加开花和结果的树种,如在夏季结果的桃Prunus persica、在冬季开花的光叶子花Bougainvillea glabra等(王勇等,2014);(3)在多样性较低的绿地内,如珠江公园,应减少大面积的纯林(如棕榈园、木兰园等),可以在考虑观赏的同时,适当增加植物垂直群落结构和生境异质性,从而提高生物多样性(隋金玲等,2006a,2006b);(4)为了给鸟类营造一个良好的栖息环境,还应当减轻人为干扰,在鸟类和人流之间设置缓冲区域。
致谢: 广东省昆虫研究所动物生态与恢复中心的袁喜才研究员、周智鑫博士、蓝道英博士对论文给予了大力指导;谢莉、曹宏芬、袁倩敏在野外调查中给予协助;西南林业大学的李苏雨和王倩同学在植物鉴别和植被参数的测定上给予帮助。在此一并致谢!| 邓娇, 晏玉莹, 张志强, 等. 2014. 城市化对长沙市城市公园繁殖期鸟类物种多样性的影响[J]. 生态学杂志 , 33(7) : 1853–1859. |
| 邓文洪, 高玮. 2005. 次生林不同类型森林边缘的鸟类物种丰富度及个体多度比较[J]. 生态学报 , 25(11) : 2804–2810. |
| 葛振鸣, 王天厚, 施文或, 等. 2005. 环境因子对上海城市园林春季鸟类群落结构特征的影响[J]. 动物学研究 , 26(1) : 17–24. |
| 华南濒危动物研究所. 1991. 广东鸟类彩色图鉴[M]. 广州: 广东科技出版社 . |
| 李敏, 陈文倩, 魏炜, 等. 2012. 内蒙古中部地区繁殖鸟类多样性调查[J]. 动物学杂志 , 4(3) : 102–108. |
| 陆祎玮, 唐思贤, 史慧玲, 等. 2007. 上海城市绿地冬季鸟类群落特征与生境的关系[J]. 动物学杂志 , 42(5) : 125–130. |
| 毛齐正, 马克明, 邬建国, 等. 2013. 城市生物多样性分布格局研究进展[J]. 生态学报 , 33(4) : 1051–1064. |
| 尚玉昌. 2007. 鸟类的食物多样性及其取食适应[J]. 生物学通报 , 42(3) : 9–13. |
| 隋金玲, 胡德夫, 李凯, 等. 2006a. 北京市区不同绿化带内夏季鸟类的群落特征[J]. 林业科学 , 42(7) : 66–72. |
| 隋金玲, 张志翔, 胡德夫, 等. 2006b. 北京市区绿化带内鸟类食源树种研究[J]. 林业科学 , 42(12) : 83–89. |
| 王勇, 许洁, 杨刚, 等. 2014. 城市公共绿地常见木本植物组成对鸟类群落的影响[J]. 生物多样性 , 22(2) : 196–207. |
| 吴征镒, 彭华, 李德铢, 等. 2004. 中国植物志(第一卷)[M]. 北京: 科学出版社 . |
| 熊咏梅, 叶凯图, 黄颖. 2013. 广州市居住区绿地小气候及影响因子分析[J]. 园林科技 , 2 : 22–24. |
| 姚月华, 颜玉娟. 2014. 柳州冬季观果树种应用与鸟类多样性调查研究[J]. 南方园艺 , 25(1) : 19–21. |
| 易兰, 程瑶, 邓军, 等. 2007. 武汉中山公园冬季鸟类群落研究[J]. 湖北林业科技 , 2 : 27–30. |
| 张钧挥. 2011. 城市绿地广场研究[D]. 西安:西安建筑科技大学. |
| 张云霞, 李晓兵, 陈云浩. 2003. 草地植被盖度的多尺度遥感与实地测量方法综述[J]. 地球科学进展 , 18(1) : 85–93. |
| 张晓明, 李强, 陈国华, 等. 2015. 花椒园昆虫群落结构及时间动态的聚类分析[J]. 应用昆虫学报 , 52(1) : 237–248. |
| 郑光美. 1995. 鸟类学[M]. 北京: 北京师范大学出版社: 402 -405. |
| Ferger SW, Schleuning M, Hemp A, et al. 2014. Food resources and vegetation structure mediate climatic effects on species richness of birds[J]. Global Ecology and Biogeography , 23(5) : 541–549. DOI:10.1111/geb.2014.23.issue-5 |
| Horváth Z, Ferenczi M, Móra A, et al. 2012. Invertebrate food sources for water birds provided by the reconstructed wetland of Nyirkai-Hany, northwestern Hungary[J]. Hydrobiologia , 697(1) : 59–72. DOI:10.1007/s10750-012-1170-5 |
| Howes J, Bakewell D, Bureau AW. 1989. Shorebird studies mannual[M]. Kuala Lumpur: Asian Wetland Bureau: 253 -572. |
| Margalef R. 1968. Perspective in ecological theory[M]. Chicago: University of Chicago Press . |
| McKinney ML. 2008. Effects of urbanization on species richness:a review of plants and animals[J]. Urban Ecosystems , 11(2) : 161–176. DOI:10.1007/s11252-007-0045-4 |
| Ren H, Shen WJ, Lu HF, et al. 2007. Degraded ecosystems in China:status, causes, and restoration efforts[J]. Landscape and Ecological Engineering , 3(1) : 1–13. DOI:10.1007/s11355-006-0018-4 |