 2016, Vol. 35
2016, Vol. 35扩展功能
文章信息
- 原宝东, 闫永峰
- YUAN Baodong, YAN Yongfeng
- 鹊鸲冬季和春季取食生态位初步研究
- Feeding Niche of Copsychus saularis between Spring and Winter
- 四川动物, 2016, 35(3): 426-430
- Sichuan Journal of Zoology, 2016, 35(3): 426-430
- 10.11984/j.issn.1000-7083.20150352
-
文章历史
- 收稿日期: 2015-11-09
- 接受日期: 2016-01-15






2. 河池学院化学与生物工程学院, 广西 宜州 546300
2. Department of Chemistry and Life Sciences, Hechi University, Yizhou, Guangxi Zhuang Autonomous Region 546300, China
鹊鸲Copsychus saularis是主要分布于我国长江以南地区的常见留鸟,共有2个亚种分化(赵正阶, 2001)。国外见于欧亚大陆、印度次大陆、中南半岛、华莱士区(Wallace)和太平洋诸岛(蒋爱伍等, 2009)。鹊鸲主要栖息于海拔2 000 m以下的低山、丘陵和山脚平原地带的次生林、竹林、林缘疏林灌丛和小块丛林等开阔地,尤其喜欢村寨和居民点附近的小块丛林、灌丛、果园以及耕地、路边和房前屋后的树林与竹林,甚至出现在城市公园和庭院树上(蒋仕仁, 2003)。目前关于鹊鸲的研究相对较少,有学者在杭州对鹊鸲的声行为进行了观察和记录,并与印度哈里瓦地区的鹊鸲进行了比较分析,有学者分析了四川省东部峨城山区鹊鸲数量不断减少的原因(王协义, 1999;刘绘, 2003)。乱砍滥伐、猎捕和农药的使用以及土地使用性质的改变使鹊鸲的栖息地丧失,造成了鹊鸲的数量锐减(郑光美, 王岐山, 1998)。
生态位是一个生物单位(个体、种群或物种)生存条件的总集合体,由于食物资源的质量直接影响鸟类的分布模式、种群密度、繁殖成功率和雏鸟存活率等,因而食物生态位研究至今仍然是生态位研究中非常活跃的一个领域(Thomas, 2008)。动物的取食生态位研究对评估生境质量、预测栖息地负载量、制定合理的保护策略和资源管理方案等均具有重要意义。通过对鸟类在不同季节取食生态位的重叠度、相似性及宽度的计算,可深入认识鸟类对资源波动性的行为适应机制(杨晓君等, 1994;王维禹等, 2005;赵欣如等, 2010)。因此,通过对广西宜州地区鹊鸲的取食行为进行直接观察,初步分析其春季和冬季取食生态位的宽度和重叠度,为以后开展鹊鸲的行为研究工作提供基础性资料,并对今后的保护和管理工作提出合理建议。
1 研究地区概况研究地区位于广西壮族自治区宜州市龙江中游,总面积4.0×105 m2,地理坐标108°4'11″~109°2'44″E、24°0'10″~24°52'5″N,属桂北九万大山余脉南端与凤凰山山脉交接地带。地处北回归线略偏北,属南亚热带季风气候区与中亚热带季风气候区的过渡带。根据气候特征划分季节:3—5月为春季,6—9月为夏季,10—11月为秋季,12月—次年2月为冬季。区内雨量充沛,干湿明显,年均降雨量1 300~1 550 mm,年均气温19.6 ℃~20.2 ℃,夏秋日照多,春季少。植物种类丰富,共有野生和栽培植物400多种,优势种有阴香Cinnamomum burmanii、大叶榕Ficus altissima、假槟榔Archontophoenix alexandrae、朴树Celtis sinensis、马尾松Pinus massoniana、构树Broussonetia papyrifera、香樟Cinnamomum camphora、金樱子Rosa laevigata、枫香Liquidambar formosana 等喀斯特地区常见植物。动物资源丰富,其中国家Ⅰ级保护动物有林麝Moschus berezovskii,国家Ⅱ级保护动物有穿山甲Manis pentadactyla、黑鸢Milvus migrans、普通Buteo buteo、燕隼Falco subbuteo等5种;自治区保护动物有眼镜蛇Naja naja、赤腹松鼠Callosciurus erythraeus等26种(覃勇荣, 覃宝山, 2008)。
2 研究方法 2.1 研究方法2013年3—5月,2014年12月—2015年2月在野外调查采用样线法和样方法,发现鹊鸲后利用8×42倍的双筒望远镜采用随机取样法和扫描取样法定点观察鹊鸲的数量和取食行为,每2 d观察一次,观察时间段为:06∶00—09∶00、09∶00—12∶00、12∶00—15∶00、15∶00—18∶00。记录食物种类、取食方式、取食生态位及取食方向,每次观察可选取不同的时间段,也可以进行连续观察或全天观察。其中取食方式分为整吞、啄食、拾取及其他;取食生态位分为地面、灌丛及乔木树冠,其中乔木树冠的垂直取食生态位由上至下依次分为上层、中层及下层,水平取食生态位由外向内依次分为外侧、中侧及内侧;取食方向以正北方向为0°,正东方向为90°,正南方向为180°,正西方向为270°,共分为4个范围:315°~45°、45°~135°、135°~225°、225°~315°。将春季、冬季的各生态位参数进行卡方独立性检验。
所有数据处理均在Excel 2010和SPSS 13.0中进行。
2.2 种群分布模式鹊鸲种群的个体在其生活空间中的位置状态和布局,采用统计样方结果中的方差/平均数(S2/m)来判断。S2/m=0为均匀分布,S2/m=1为随机分布,S2/m>1为集群分布。其中:

|
式中x为样本中含有的鹊鸲数量,f为出现频率(含有不同个体数样本的出现频率),N为样本总数。方差反映了样本与平均数的差异程度,方差越大,样方间鹊鸲种群密度的差异越大。
2.3 数据处理方法 2.3.1 取食生态位宽度取食生态位宽度(feeding niche breadth)用Simpson多样性指数(B)表示(吴诗宝等, 2005)。
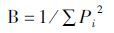
|
式中,Pi为实际利用的第i种食物在所有被利用的食物中所占的比例。
2.3.2 取食生态位重叠度取食生态位重叠度(feeding niche overlap)用Levins(Ojk或Okj)指数表示。
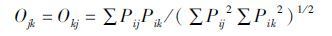
|
式中,Pij和Pik分别为被j和k组鹊鸲利用的第i种食物在所有被利用的食物中所占比例。
3 结果及分析 3.1 鹊鸲种群的分布型鹊鸲以单独觅食为主,偶尔有雌雄2只共同觅食,超过3只共同觅食时,它们之间会发生食物竞争的冲突,春季2只共同觅食发现比例明显超过冬季。将数据运用格局指数进行计算,结果表明,研究地区冬季的鹊鸲种群分布格局指数为0.99,为随机分布;春季的为1.13,为集群分布(表 1)。
| 参数 | 冬季 | 春季 | |||||
| 平均值±标准差( | 方差(S2) | 格局指数(S2/m) | 平均值±标准差( | 方差(S2) | 格局指数(S2/m) | ||
| 结果 | 1.15±0.14 | 0.47 | 0.99 | 1.37±0.26 | 0.63 | 1.13 | |
根据观察记录的数据,鹊鸲的取食行为数据见表 2,并对不同季节的取食生态位宽度和重叠度进行分析。从结果可以看出鹊鸲为杂食性鸟类,以果实和昆虫为主,冬季取食的植物有23种,取食生态位宽度为11.24;春季取食的植物有22种,取食生态位宽度为12.19;春季和冬季的取食生态位重叠度为0.359。取食都是用喙啄食。鹊鸲喜欢在高大乔木、灌丛、草地和地面上独自取食,取食植物上的果实及地面上的落果,还取食植物枝条上和地面上的昆虫,有时也会在生活垃圾中寻找食物。鹊鸲春季和冬季的取食生态位宽度都较高,这与不同季节可供取食的果实的种类不同有关。
| 植物种类 | 食物组成 | 取食频次比例/% | |
| 冬季 | 春季 | ||
| 香樟Cinnamomum camphora | 果实 | 1.88 | 13.08 |
| 苦楝Melia azedarach | 果实 | 8.22 | 13.43 |
| 大叶榕Ficus altissima | 果实 | 11.07 | 9.13 |
| 雅榕Ficus concinna | 果实 | 7.80 | 6.67 |
| 构树Broussonetia papyrifera | 果实、花 | 3.22 | 9.75 |
| 阴香Cinnamomum burmanii | 果实 | 13.90 | 2.55 |
| 佛肚竹Bambusa ventricosa McCluore | 昆虫 | 1.41 | 0 |
| 桂花Osmanthus fragrans | 果实、花 | 15.68 | 2.19 |
| 桃Amygdalus persica L. | 昆虫 | 4.22 | 3.25 |
| 台湾相思Acacia confuse Merr. | 昆虫、花 | 1.63 | 2.72 |
| 柿Diospyros kaki Thunb. | 果实 | 3.22 | 0.61 |
| 三角梅Bougainvillea spectabilis Wild. | 花 | 1.88 | 2.90 |
| 小叶紫薇Lobular crape myrtle | 昆虫、花 | 1.10 | 0 |
| 鱼尾葵Caryota ochlandra Hance | 果实 | 0.39 | 0 |
| 白玉兰Magnolia denudate | 昆虫 | 10.39 | 0 |
| 侧柏Platycladus orientalis(L.)France | 昆虫 | 3.19 | 0 |
| 女贞Ligustrum lucidum | 果实 | 3.22 | 0 |
| 芒果Mangifera indica L. | 果实 | 2.90 | 0 |
| 枇杷Eriobotrya japonica(Thunb.)Lindl | 果实 | 3.68 | 0 |
| 桉树Eucalyptus robusta Smith | 昆虫 | 0.66 | 0 |
| 芭蕉Musa basjoo Sieb. et Zucc. | 果实 | 0.56 | 0 |
| 合欢Albizia julibrissin Durazz. | 花、昆虫 | 0.24 | 0 |
| 重阳木Bischofia polycarpa | 昆虫 | 2.02 | 0 |
| 秋枫Bischofia javanica | 昆虫 | 0 | 4.04 |
| 木棉Bombax ceiba | 花 | 0 | 3.34 |
| 撑蒿竹Bambusa pervariabilis McClure | 昆虫 | 0 | 12.12 |
| 青皮竹Bambusa textilis McClure | 昆虫 | 0 | 4.57 |
| 枫香Liquidambar formosana | 昆虫 | 0 | 2.99 |
| 枫杨Pterocarya stenoptera | 昆虫 | 0 | 0.61 |
| 乌桕Sapium sebiferum(L.)Roxb. | 果实 | 0 | 0.70 |
| 羊蹄甲Bauhinia purpurea L. | 花 | 0 | 2.28 |
| 九里香Murraya exotica | 果实 | 0 | 0.88 |
| 假连翘Duranta repens L. | 果实 | 0 | 1.76 |
| 杏李Prunus simonii | 昆虫 | 0 | 0.44 |
| 取食生态位宽度 | 11.24 | 12.19 | |
| 取食生态位重叠度 | 0.359 | ||
通过对鹊鸲春季和冬季的取食时间、食物种类、取食方式、取食高度、取食水平和取食方向等进行分析,结果见表 3。
| 参数 | 食物种类 | 取食时间 | 取食水平 | 取食高度 | 取食方向 |
| P值 | 0.019 | 0.038 | 0.564 | 0.513 | 0.999 |
卡方独立性检验表明:鹊鸲春季和冬季的取食时间和食物种类的差异有统计学意义(P<0.05),取食高度、取食水平和取食方向等的差异无统计学意义(P>0.05),取食方式为啄食。取食果实种类不同可能与不同季节果实的成熟期不同有关;春季的主要取食时间在06∶00—09∶00,比例为50.13%,冬季的主要取食时间在09∶00—12∶00,比例为32.12%,取食时间受季节的影响较大,冬季早上寒冷,鹊鸲为了减少能量的散失会推迟取食时间,这是对环境的行为适应。鹊鸲春季和冬季在取食高度、取食水平和取食方向等的差异无统计学意义,这可能与研究区植物种类组成单一和挂果在外侧并朝阳有关。
4 讨论 4.1 鹊鸲的取食生态位食物是影响鸟类生境利用和选择的主要因素,鸟类取食行为的适应机制是对食物丰富程度的响应。鸟类取食地选择的问题,不仅与鸟类本身的食性相关,也与栖息地结构以及食物资源状况相关。它可以揭示鸟类在其生存环境中的地位和作用,以及鸟类与环境之间的关系(杨晓君等, 1994)。鹊鸲不同时段取食频次的差异可能与以下几方面原因有关:经过一晚的休息,鹊鸲消化道内的食物已经消化完毕;早晨人类的活动较少,对鸟类的干扰较少,这是造成早上取食频次相对较多的主要原因。以往的研究指出,大多数昼行性动物都具有早上进食的行为,以补充夜晚能量的消耗(庞秉璋, 1981;韦明思等, 2004)。鸟类的取食行为并不是一成不变的,它随着环境条件、季节、个体发育阶段不同以及新情况的出现而发生变化,是对环境适应的一个重要方面。然而随着季节的变化,冬季与春季相比温度要低一些,鹊鸲会推迟外出活动取食的时间,在其他鸟类的研究中也得出了类似的结果(Yagihashi et al., 1998)。
生态位宽度是物种利用或趋于利用所有可利用资源状态,一个物种的生态位宽度越宽,其特化程度越小,也就是说该物种更倾向于一个泛化种;相反,一个物种的生态位宽度越窄,该种的特化程度越大,即更倾向于一个特化种。生态位重叠是个体与生态因子联系上的相似性,当物种利用同一资源(食物、营养成分、空间等)时,就会出现生态位重叠现象,生态位重叠程度能体现种群对共同资源的利用状况(van Den Wyngaert et al., 2003)。在研究地区,冬季挂果的植物相对较少,春季植物开始冒出新芽,有些春季结果实的植物为鹊鸲提供了更多的食物选择,这使得春季的取食生态位宽度略大于冬季。春季和冬季的取食生态位重叠度很低可能与植物果实的成熟季节不同有关(Kwit et al., 2004)。
在进化过程中,动物选择了适应本身条件发展的最佳取食策略。鹊鸲属于小型鸟类,2个季节能够取食的植物种类达到30多种,这与早期报道中小型鸟类食物选择性较小,通过提高觅食活动率、增加食物量、拓宽取食生态位相符(Barnea et al., 1991)。研究过程中发现,由于研究地区冬、春季昆虫资源相对较少(邓维安, 2003),可以提供的肉食性食物有限,鹊鸲会更多地取食果实,这与早期报道杭州地区鹊鸲以肉食性为主的结果有差异(蒋仕仁, 2003),同时从另一方面表明鹊鸲是一种适应能力较强的鸟类,能够根据食物资源的可利用性调整取食策略。一年之中夏季是昆虫多样性丰富的季节,秋季是果实成熟的主要季节,下一步开展季节变化对鹊鸲取食行为的影响研究是理解鹊鸲生活史策略的关键。
4.2 鹊鸲对种子的潜在传播作用通过对2个季节鹊鸲的取食可以看出,鹊鸲主要以果实为主,也会取食昆虫、垃圾中的食物,这对抑制虫害起到了一定的作用。鹊鸲喜欢在树冠高处取食,这与植物树冠层采光充足、果实发育相对饱满、食物营养更多有关。许多植物通过鸟类传播果实,Barnea等(1991)研究表明,鸟类的消化过程能够明显提高种子的萌发率,甚至使种子由不能萌发变为能够萌发,原因是鸟类消化了果肉,而果肉中可能存在萌发抑制物质(Cipollini & Levey, 1997)。鹊鸲在啄食果实的过程中会将种子附带吞入体内,未消化的种子以粪便的形式排出体外,从而成为了种子的传播者。在调查中也发现,在离母树很远的距离发现了苦楝Melia azedarach、桂花Osmanthus fragrans等幼苗,这些幼苗很可能是借助食果动物的传播才得以在适宜的生境萌发。动物对种子传播的生态学是值得深入研究的重要领域(申仕康等, 1998)。
研究过程中可见成对的鹊鸲在房顶上、空中打斗或嬉戏,停靠在电线杆上鸣叫,或者互相鸣叫。对于鹊鸲如何筑巢、繁殖,目前还没有相关的研究。研究期间曾发现鹊鸲在长穗鱼尾葵Caryota ochlandra和桂花上筑巢,深入研究鹊鸲的繁殖生物学才能更好地了解该物种的进化适应机制。
致谢: 感谢河池学院本科生雷志栋和张小娟同学在野外数据收集过程中给予的帮助。