 2023, Vol. 44
2023, Vol. 44文章信息
- 赵锦, 杨学文, 刘如春, 周银柱, 陈水连, 邱劲松, 黄政.
- Zhao Jin, Yang Xuewen, Liu Ruchun, Zhou Yinzhu, Chen Shuilian, Qiu Jinsong, Huang Zheng
- 长沙市1例人感染H3N8禽流感病例的流行病学调查和病原学分析
- Epidemiological investigation and etiological analysis on a case of human infection with avian influenza A (H3N8) virus in Changsha
- 中华流行病学杂志, 2023, 44(11): 1776-1780
- Chinese Journal of Epidemiology, 2023, 44(11): 1776-1780
- http://dx.doi.org/10.3760/cma.j.cn112338-20230524-00329
-
文章历史
收稿日期: 2023-05-24




H3N8禽流感病毒宿主范围广,除感染禽类外,还可感染马、犬、骆驼等哺乳动物,且跨越物种屏障传播广泛存在[1]。2022年4月26日河南省驻马店市通报1例人感染H3N8禽流感病例,5月19日湖南省长沙市通报1例人感染H3N8禽流感病例。以上案例表明,H3N8禽流感病毒存在感染人类的风险。本研究对长沙市报告病例的流行病学特征、分子生物学特征和临床特征进行分析,探讨人感染H3N8禽流感发生和传播途径,为人感染H3N8禽流感防控提供科学依据。
对象与方法1. 研究对象:2022年5月长沙市CDC在1份流感哨点监测标本中检测到H3N8禽流感病毒核酸阳性。本研究对该病例临床诊治、流行病学史、密切接触者、可疑暴露者、可疑禽类及暴露场所开展调查。
2. 流行病学调查:依据《人感染动物源性流感预防控制技术指南(试行)》对病例进行流行病学调查[2],内容主要包含病例基本情况、家居环境、发病及就诊经过、既往就诊信息、暴露史、密切接触者医学观察情况、相关涉疫场所等,依此进行病例搜索、应急监测和涉疫重点人员管理。
3. 标本采集:采集病例、密切接触者、可疑暴露者咽拭子标本,病例家外环境涂抹标本、可疑暴露场所涂抹标本、可疑禽类粪便及禽饮水标本,病例、密切接触者、发热病例(病例搜索)及其家属血清标本。所有标本按照B类感染性物质在4 ℃条件下进行低温包装运输。
4. 实验室检测:由长沙市CDC完成核酸检测及基因测序。所有标本在生物安全柜中预处理后,使用江苏硕世核酸提取试剂盒(磁珠法)对标本进行核酸提取,使用江苏硕世流感病毒核酸荧光PCR试剂进行甲型流感病毒(IAV)核酸检测以及亚型鉴定。对于病例H3N8禽流感阳性标本采用二代测序技术进行基因测序,使用Illumina公司的二代测序miseq平台,以nextera XT文库构建试剂盒,下机数据先用Fastqc和Cutadapt软件进行数据质量分析和去接头,筛选后的数据用中国CDC的Virus Identification Pipeline软件进行比对分析,数据过滤后再用SPAdes 3.13.0软件进行序列拼接与组装。采用BioEdit软件进行氨基酸位点分析。
结果1. 病例基本情况:男性,5岁,汉族,长沙籍,居住于长沙市A小区Y栋,就读于B幼儿园大班。与父母、祖父母共同居住,日常起居主要由祖父母负责。既往身体健康,无慢性病。发病前10 d未接触过发热、咳嗽等呼吸道症状病例。
2. 发病及临床诊治过程:病例于2022年5月9日晚出现发热(39 ℃)、畏寒症状,当晚先后服用头孢克洛干混悬剂、布洛芬混悬液,并持续用毛巾擦拭进行物理降温。
5月10日上午先后2次服用布洛芬混悬液,发热未见缓解(40 ℃),遂由家属送至C医院发热门诊就医,并采集咽拭子作为流感哨点监测标本送至长沙市CDC检测。体格检查:咽部充血,双侧扁桃体Ⅱ度肿大,双肺呼吸音粗,未闻及明显啰音。血常规示淋巴细胞比率为14.1%(降低)、嗜酸性粒细胞比率为0.3%(降低),中性粒细胞比率为77.5%(升高)。诊断为急性扁桃体炎,予以阿莫西林克拉维酸、开喉剑喷雾剂、小儿豉翘清热颗粒治疗,未住院。当日下午仍发热(37.6 ℃),由家属前往D社区卫生服务中心购药,社区医生根据家属的病情描述及C医院诊治情况,诊断为急性扁桃体炎、急乳蛾病、风热犯肺证,开具中药配方颗粒治疗(配方为金银花、连翘、浙贝母、桔梗、石膏、防风、蝉蜕、柴胡、甘草、芦根、地骨皮、薄荷、葛根)。病例当晚体温恢复正常,其后未再出现发热、畏寒等不适症状。
5月11-12日继续服用阿莫西林克拉维酸和中药配方颗粒。5月13-15日未服用任何药物。因作为流感哨点监测采集的咽拭子标本检出H3N8禽流感病毒阳性,5月16日转入C医院单间隔离,体格检查正常,无不适症状。血常规结果:单核细胞数为0.28×109/L(降低)、淋巴细胞比率为49.2%(升高)、中性粒细胞比率为39.7%(降低);胸部电子计算机断层扫描平扫未见明显异常。予以奥司他韦治疗(45 mg,一天2次)。5月19、20日H3N8禽流感病毒核酸均为阴性,5月20日出院。
3. 流行病学调查:病例发病前1个月未离开长沙市,日常活动范围局限于A小区及B幼儿园周边,5月3日由祖母带其前往E农贸市场购买蔬菜,停留约20 min,往返途中均经过E农贸市场内的5个活禽交易摊位,但未直接接触。离开E农贸市场后2人前往F生鲜超市购买豆制品,豆制品与冷鲜禽肉摆放在同一层冷柜中销售,停留约10 min。2人在E农贸市场和F生鲜超市活动期间均佩戴口罩(成年人型)。发病前10 d家中未购买或食用禽类及禽肉制品,未直接接触活禽、死禽、禽肉制品。
A小区Y栋共7层,只有1个出入口,无电梯。病例家住7层,通风良好,家中未养鸡鸭等禽类。病例平时与祖父母同睡一屋,日常由祖母负责购买食材,购买途径以网购及F生鲜超市为主,较少去E农贸市场。
病例所在楼栋与邻近楼栋之间有一绿化草坪,5月6-16日有业主曾在此圈养2只鸡。圈养地距离Y栋出入口5 m,病例外出活动时距圈养地距离均超过3 m。鸡为5月6日从外市农村带回,其中1只5月14日已宰杀食用,另一只5月16日在动物卫生监督部门指导下进行无害化处理。
E农贸市场为小型农贸市场,面积约1 000 m2,有摊位40个,工作人员80人,市场内通风条件较差。现场调查发现工作人员均未佩戴口罩,顾客多数未佩戴口罩。E农贸市场有5个活禽摊位(Ⅰ~Ⅴ),各活禽摊位日销售禽类20~30只均来源于H农产品批发市场,其中Ⅰ、Ⅱ、Ⅳ、Ⅴ摊位的活禽供货商不固定,Ⅲ摊位的活禽主要来源于J活禽批发商铺。经调查,近1个月各活禽摊位均未出现病/死禽。F生鲜超市面积约38 m2,通风较好,工作人员6人。工作人员均佩戴口罩,顾客多数未佩戴口罩。销售种类为果蔬、鸡蛋、鲜肉、冷鲜禽肉等,每日销售禽类4~5只。B幼儿园为1栋3层楼房,通风良好,园内整体环境良好。病例所在班级有教师及保育员共3人,学生36人,师生均未佩戴口罩。
4. 病例搜索:搜索自4月29日以来,A小区居民、B幼儿园教职工及幼儿、E农贸市场及F生鲜超市工作人员中出现发热、咳嗽、咽痛、鼻塞、流涕等呼吸道症状者,共发现1例发热患儿。该患儿与病例同班级,于5月13日出现发热、头痛、咽痛、鼻塞症状。
5. 禽流感病原学监测:
(1) 应急监测:病例所在行政区的二级及以上医疗机构开展为期2周(5月16-31日)的流感样病例和发热肺炎病例应急监测,发现1例H3N2亚型流感病例。
(2) 年度监测:长沙市2017-2021年在全市范围内对活禽交易市场开展禽流感病毒外环境监测,检测项目为IAV、H5、H7、H9亚型核酸。监测结果显示外环境IAV、H5、H7、H9亚型阳性率分别为59.03%(817/1 384)、14.87%(128/861)、5.60%(83/1 384)、44.13%(380/861)。活禽交易市场普遍存在禽流感病毒污染。见图 1。

|
| 图 1 长沙市2017-2021年活禽交易市场环境标本禽流感病原学监测结果 |
6. 涉疫重点人员管理:经流行病学调查,结合实验室检测结果,共判定密切接触者4人,均为病例家属。按要求开展居家健康监测,判定可疑暴露者18人,其中F生鲜超市工作人员6人、散养鸡业主及其家属3人、E农贸市场内及周边活禽摊位工作人员9人。对以上人员开展自我医学观察。一般接触者37人,为病例同班级师生,要求开展自我医学观察。
7. 采样及实验室检测结果:
(1) 病例标本:5月10、15日咽拭子均为禽流感H3亚型阳性、N8亚型阳性;5月10日咽拭子经基因测序为H3N8禽流感。5月19、20日咽拭子均为IAV核酸阴性。
经基因深度测序,序列分析显示病毒的6个内部基因片段均来自H9N2禽流感病毒;HA蛋白裂解位点为PEKQTR↓G,提示对禽类低致病性;HA蛋白226和228位分别为Q、G,提示病毒主要结合禽流感病毒受体;PB2蛋白627位由E突变为V,提示发生了哺乳动物适应性突变;M2蛋白31位突变为N提示对烷胺类药物敏感性降低;未检测到神经氨酸酶抑制剂类药物和聚合酶抑制剂类药物相关的耐药性突变。
(2) 其他人员咽拭子标本:5月15日采集10份咽拭子,其中病例家属4人,F生鲜超市工作人员6人;5月16日采集64份咽拭子,其中5个活禽交易摊位工作人员9人,同班级发热患儿及其家属4人,A小区散养鸡业主及其家属3人,B幼儿园教职工14人,病例同班级其他幼儿34人。均为IAV核酸阴性。
(3) 禽类及相关环境标本:5月15日采集病例家环境涂抹样本5份、A小区散养鸡所在场所环境涂抹样本及粪便、禽类饮水等标本4份、F生鲜超市环境涂抹样本16份,均为IAV核酸阴性;5月16日采集E农贸市场活禽交易摊位环境涂抹样本10份(每个摊位2份),均为IAV核酸阳性,其中4个摊位为禽流感H3、N8亚型阳性。5月16-25日,先后多次前往H批发市场采集活禽批发商铺环境涂抹样本、禽类粪便、空气、禽类饮水等标本共55份,其中40份为IAV阳性,17份为禽流感H3亚型阳性,9份为N8亚型阳性。活禽交易市场(E农贸市场、H批发市场)禽流感H3、N8亚型阳性率分别为38.46%(25/65)、23.08%(15/65)。见表 1。
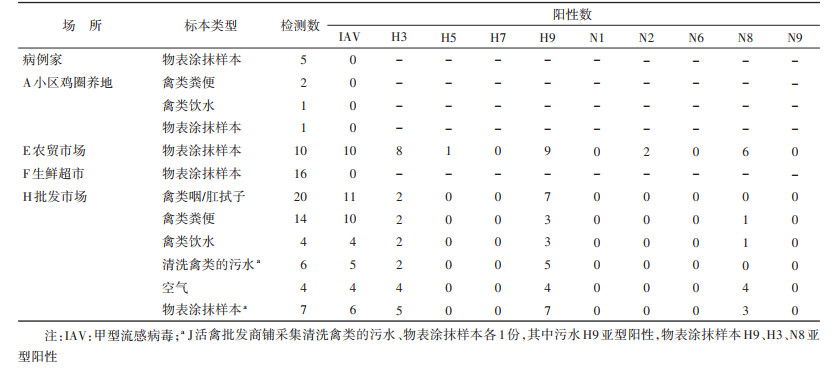
本案例为人感染H3N8禽流感病例,对病例感染来源、感染途径、传播能力进行调查分析,可为今后人感染H3N8禽流感疫情防控工作带来新的启发和思考。
接触携带病毒的动物或暴露于被病毒污染的环境是感染动物源性流感的主要途径。病例发病前10 d未接触过发热病例,发病前6 d有活禽交易市场暴露史。后续在E农贸市场、H农产品批发市场采集的多个标本中检测到禽流感H3、N8亚型阳性,且在固定供货的J活禽批发商铺检测到禽流感H3、N8亚型阳性。结合流行病学调查,判定病例是在E农贸市场暴露于被病毒污染的环境而感染。该活禽交易市场工作人员、病例祖母和其他人均未感染H3N8禽流感病毒,提示H3N8禽流感的感染存在个体差异性。
病例家属、同幼儿园师生均未发现人感染H3N8禽流感病例。病例从5月9日晚出现发热症状,5月10日晚症状消退,期间并未服用神经氨酸酶抑制剂类药物和聚合酶抑制剂类药物。但由于全球报道的病例数有限,尚无法判断H3N8禽流感病毒在人际间的传播能力以及对人群的致病能力。基因深度测序显示,H3N8禽流感病毒对神经氨酸酶抑制剂类药物和聚合酶抑制剂类药物敏感,提示在治疗人感染H3N8禽流感病例时,以上药物依然有效。
活禽交易市场应急监测发现禽流感H3、N8亚型阳性率分别为38.46%、23.08%,结合年度监测结果,活禽交易市场可能普遍存在H3N8禽流感病毒污染,提示可能存在一定数量病例未被发现。建议各级疾病预防控制机构将H3N8纳入禽流感常规监测,定期对活禽交易市场禽类及环境、职业人群开展H3N8禽流感病毒核酸检测。医疗机构对于有活禽交易市场及禽类暴露史的流感样病例,若IAV核酸阳性,常规流感(H1N1、H3N2)亚型阴性,除进一步检测禽流感H5、H7、H9亚型外,还要考虑检测禽流感H3亚型,以免发生漏诊。
本案例基因测序显示,HA蛋白裂解位点为PEKQTR↓G,提示对禽低致病性。HA蛋白226和228位分别为Q、G,提示病毒主要结合禽流感病毒受体。PB2蛋白627位由E突变为V,提示发生了哺乳动物适应性突变。显示H3N8禽流感病毒对禽类易感且为低致病性,因此禽类感染变得更隐匿,难以发现。与此同时,病毒又发生了对于哺乳动物的适应性突变,可能更容易突破物种屏障,亦可能对人类更易感。应强化对活禽养殖场、交易市场、相关从业人员的常态化监测,做到早发现、早处置,尽早落实各项防控措施。
中国人的饮食文化注重食物的“鲜活”,人们通常会购买活禽现场宰杀,活禽在市场的销售、存栏过夜及宰杀处理都会造成禽流感病毒污染环境。人在售卖活禽的场所活动时会暴露于被污染的环境,且在购买活禽的过程中会直接接触禽类,均有感染禽流感病毒的风险。研究显示,不同场所的禽类及环境禽流感病毒阳性率有显著差异,由高至低分别为活禽交易市场、散养户、野禽栖息地、养殖场,且活禽交易市场阳性率显著高于其他场所[3-5],提示存在广泛交叉污染。活禽交易市场是人感染禽流感病毒的主要场所[6],禽类“集中屠宰,生鲜上市”是防控人感染禽流感的主要措施。现有的疫情处置经验表明,关闭活禽交易市场后,人感染禽流感病毒发病率明显降低[7-8]。休市也是防控禽流感的重要措施。休市虽然可以迅速减少市场污染,但复市后病毒阳性率会迅速升高,且环境阳性率上升速度明显高于活禽阳性率[5]。提示应大力发展规模化禽类养殖,规范接种禽流感疫苗,注重禽类上市前检疫,从源头上消除传染源。要大力宣传禽流感防控知识,改变人们食用“鲜活”禽类习惯,推广冷鲜禽类,逐步取消活禽销售,减少人暴露于禽流感病毒的机会。
H3N8禽流感病毒在野鸟和家禽中普遍存在,近年来的禽类监测结果显明,H3N8禽流感病毒的检出率呈逐年上升趋势,由禽类跨越物种感染哺乳动物的情况时有发生[1]。目前该亚型病毒对禽类表现为低致病性,但马、犬、海豹感染后可引起明显呼吸道症状甚至死亡[9-11]。动物实验显示,H3N8禽流感病毒已经获得了在雪貂的呼吸道飞沫传播能力,而雪貂的呼吸道唾液酸受体分布与人类相似,上呼吸道含有大量人型受体,故雪貂通常被用来做动物模型评价IAV对人类的感染及免疫过程[12-13]。H3N8禽流感病毒对哺乳动物和人类的威胁持续增加,且存在人传人的风险。本研究病例感染存在一定的偶然性和个体差异性,但从咽拭子分离的标本基因测序发现,病毒发生了哺乳动物适应性突变,这表明H3N8禽流感病毒存在感染人的分子学基础。综上,不排除H3N8禽流感病毒进一步突变,从而形成人际间传播。应加强禽类的H3N8流感病毒监测,密切关注其突变情况。
利益冲突 所有作者声明无利益冲突
作者贡献声明 赵锦、杨学文、刘如春:数据整理、论文撰写/修改;周银柱、陈水连、邱劲松、黄政:论文修改、文献查阅
| [1] |
崔鹏飞. 2009-2013年中国H3N8亚型禽流感病毒生物学特性研究[D]. 兰州: 甘肃农业大学, 2016. Cui PF. Biological characterization of H3N8 avian influenza viruses isolated from 2009 to 2013 in China[D]. Lanzhou: Gansu Agricultural University, 2016. |
| [2] |
国家卫生健康委员会. 人感染动物源性流感预防控制技术指南(试行)[J]. 传染病信息, 2021, 34(5): 385-392. DOI:10.3969/j.issn.1007-8134.2021.05.001 National Health Commission. Guidelines for the Prevention and Control of Animal-derived Influenza in Human Infection (Trial)[J]. Infect Dis Inf, 2021, 34(5): 385-392. DOI:10.3969/j.issn.1007-8134.2021.05.001 |
| [3] |
赵腾. 禽流感病毒及其感染风险的流行病学监测研究[D]. 北京: 军事科学院, 2018. Zhao T. Epidemiological surveilence study on avian influenza virus and its infection risk[D]. Beijing: Academy of Military Science, 2018. |
| [4] |
焦铭. 2016-2017年度鄱阳湖地区禽流感病毒监测与进化分析[D]. 北京: 中国疾病预防控制中心, 2018. Jiao M. Surveillance and evolution analysis of avian influenza virus in Poyang Lake Region, 2016-2017[D]. Beijing: Chinese Center for Disease Control and Prevention, 2018. |
| [5] |
刘涛, 祝光湖, 张兵, 等. 我国活禽交易市场休市对人感染H7N9禽流感流行的影响[J]. 中华流行病学杂志, 2017, 38(12): 1716-1718. DOI:10.3760/cma.j.issn.0254-6450.2017.12.027 Liu T, Zhu GH, Zhang B, et al. The effects of closure to live poultry markets on avian influenza A (H7N9) epidemics in China[J]. Chin J Epidemiol, 2017, 38(12): 1716-1718. DOI:10.3760/cma.j.issn.0254-6450.2017.12.027 |
| [6] |
Wang W, Artois J, Wang XL, et al. Effectiveness of live poultry market interventions on human infection with avian influenza A(H7N9) virus, China[J]. Emerg Infect Dis, 26(5): 891-901. DOI: 10.3201/eid2605.190390.
|
| [7] |
He F, Chen EF, Li FD, et al. Human infection and environmental contamination with Avian Influenza A(H7N9) Virus in Zhejiang Province, China: risk trend across the three waves of infection[J]. BMC Public Health, 2015, 15: 931. DOI:10.1186/s12889-015-2278-0 |
| [8] |
Yu HJ, Wu JT, Cowling BJ, et al. Effect of closure of live poultry markets on poultry-to-person transmission of avian influenza A H7N9 virus: an ecological study[J]. Lancet, 2014, 383(9916): 541-548. DOI:10.1016/S0140-6736(13)61904-2 |
| [9] |
Crawford PC, Dubovi EJ, Castleman WL, et al. Transmission of equine influenza virus to dogs[J]. Science, 2005, 310(5747): 482-485. DOI:10.1126/science.1117950 |
| [10] |
Yondon M, Heil GL, Burks JP, et al. Isolation and characterization of H3N8 equine influenza A virus associated with the 2011 epizootic in Mongolia[J]. Influenza Other Respir Viruses, 2013, 7(5): 659-665. DOI:10.1111/irv.12069 |
| [11] |
Hussein ITM, Krammer F, Ma E, et al. New England harbor seal H3N8 influenza virus retains avian-like receptor specificity[J]. Sci Rep, 2016, 6: 21428. DOI:10.1038/srep21428 |
| [12] |
Karlsson EA, Ip HS, Hall JS, et al. Respiratory transmission of an avian H3N8 influenza virus isolated from a harbour seal[J]. Nat Commun, 2014, 5: 4791. DOI:10.1038/ncomms5791 |
| [13] |
Belser JA, Lu XH, Maines TR, et al. Pathogenesis of avian influenza (H7) virus infection in mice and ferrets: enhanced virulence of Eurasian H7N7 viruses isolated from humans[J]. J Virol, 2007, 81(20): 11139-11147. DOI:10.1128/JVI.01235-07 |