 2016, Vol. 37
2016, Vol. 37文章信息
- 高强, 熊成龙, 周毅彬, 曹晖, 姜庆五.
- Gao Qiang, Xiong Chenglong, Zhou Yibin, Cao Hui, Jiang Qingwu.
- 上海中心城区白纹伊蚊侵害状况及蚊媒传染病危险性分析
- Infestation status Aedes albopictus and related mosquito-borne infectious disease risk in central urban area in Shanghai
- 中华流行病学杂志, 2016, 37(5): 600-605
- Chinese Journal of Epidemiology, 2016, 37(5): 600-605
- http://dx.doi.org/10.3760/cma.j.issn.0254-6450.2016.05.003
-
文章历史
- 投稿日期: 2016-02-25




2. 200032 上海, 复旦大学公共卫生学院卫生微生物学教研室;
3. 200023 上海市黄浦区疾病预防控制中心病媒生物防治科;
4. 200336 上海市疾病预防控制中心病媒生物防治科
2. Department of Public Health Microbiology, School of Public Health, Fudan University, Shanghai 200032, China;
3. Department of Vector Control, Huangpu District Center for Disease Control and Prevention, Shanghai 200023, China;
4. Department of Vector Control, Shanghai Municipal Center for Disease Control and Prevention, Shanghai 200336, China
白纹伊蚊(Aedes albopictus Skuse,1894)又称为“亚洲虎蚊”(Asian tiger mosquito),最早发现于东南亚热带雨林[1],近30年由于人类活动而造成全球地区的迅速扩展[2]。作为最重要的传染病媒介,白纹伊蚊可以传播包括黄热病毒属(Flavivirus)、阿尔法病毒属(Alphavirus)以及布尼亚病毒属(Bunyavirus and Phlebovirus)等在内的至少26种病毒[1],最具代表性的是登革热病毒(Dengue virus,DENV)[3]、基孔肯雅病毒(Chikungunya virus)[3]及寨卡病毒(Zika virus)[4]等。白纹伊蚊在国内分布范围较埃及伊蚊更加广泛,因此是国内大部分地区登革热的最主要传播媒介[5]。
白纹伊蚊嗜吸人血[6, 7],是上海地区造成叮咬骚扰的重要蚊种,上海城区人群密集,城市环境衍生出较多特异的蚊虫孳生地,白纹伊蚊作为潜在的病毒传播媒介逐渐引起政府及卫生部门的重视,但白纹伊蚊在上海城区的侵害特点及传染疾病的风险性研究依然较少,本研究基于近年来专项开展的成蚊连续性观察,对登革热、寨卡病毒病媒介白纹伊蚊在上海市中心城区的侵害特征、种群变化趋势及危险性进行分析。
资料与方法1. 实验器材:CO2捕蚊机,蝙蝠王捕蚊机(基础型)为上海申雷节能设备技术有限公司产品;MOTIC SMZ-168体式显微镜为麦克奥迪(厦门)医疗诊断系统有限公司产品;昆虫DNA提取试剂盒(Omega公司,货号D0926)。
2. 成蚊密度观察方法:①观察区域及观察点设置:选择上海城区中心地带黄浦区人民广场和人民公园区域以及周边的3个住区类型。人民广场和人民公园区域为上海市的绝对中心,是城区最典型的公园绿地区域,人民公园按照西、中、东区域设置3个观察点;人民广场按照东、西区域设置2个观察点。为增加可比性,居民区观察点选择人民广场和人民公园周边的区域,选取的3个相邻居民区分别代表上海城区不同的居民区类型:第一种住区为石库门类型老式居民区,该类型居民区是上海老城区独特的居民区类型,观察点设于小区建筑物的背阴面;第二种为新式的高层住宅楼小区,该类型居民区是上海现代化都市建设中快速扩增的一类居民区,观察点设于小区中央的绿化地带;第三种为老式的别墅小区,观察点位于某栋别墅庭院绿化区内。②观察方法:观察时间为4月下旬至11月下旬,2012与2013年进行连续性、无间断的(每年合计224 d)成蚊密度观察,2014年每周观察1 d,采用CO2捕蚊机观察法:24 h连续开机观察,为保证效果,机器配置的CO2气体钢瓶(8 L)每5天更换,诱饵每10天更换。
3. 白纹伊蚊基因扩增:参照昆虫DNA提取试剂盒使用说明书提取白纹伊蚊DNA,参考廉国胜等[8]报道,采用通用引物扩增真核生物细胞色素C氧化酶亚基Ⅰ(COⅠ)DNA条形码序列基因序列,经凝胶电泳检测目的条带后,将扩增反应液送华大基因南方中心(上海分部)进行测序。
4. 基因序列分析:基因测序结果以Lasergene 7.0软件进行序列对位和快速进化树构建,选取代表性序列进行局部序列比对基本检索( http://blast.ncbi.nlm.nih.gov/Blast.cgi),最高匹配数(Max target sequences)设为1 000,全部下载各自返回的1 000个参考序列,合并、去重,采用Mega 6.0软件进行序列对位,以Kimura 2-parameter model构建N-J进化树,采用1 000个多序列组(replicates)进行Bootstrap验证。
5. 统计学分析:采用SPSS 13.0软件进行统计学分析,组间密度采用单因素方差分析,LSD两两比较;构成比采用Pearson χ2检验,P<0.05表示差异有统计学意义。
结 果1. 白纹伊蚊种群密度:2012-2014年CO2捕蚊机法成蚊观察结果显示,上海中心城区未发现埃及伊蚊(Aedes aegypti Linneaus,1762),绿地与居民区捕捉白纹伊蚊分别为7 015与12 002只,两种环境白纹伊蚊构成比差异明显,住区白纹伊蚊构成比显著高于绿地环境(78.53% vs. 19.99%,χ2=15 525.168,P=0.000)。根据优势度指数计算公式D=Nmax/N(其中Nmax为优势种的个体数;N为功能团全部物种的个体数),D>10%为优势种[9],白纹伊蚊在上海市中心住区与绿地环境中均为优势蚊种(表 1)。
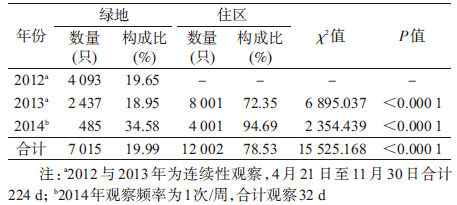
2012-2014年白纹伊蚊种群密度季节消长分析显示,上海城区白纹伊蚊高峰期主要在7-9月。白纹伊蚊密度采用单因素方差分析(one-way ANOVA)比较,结果显示不同年份白纹伊蚊密度差异有统计学意义(F=7.704,P<0.001),因数据方差不齐(Levene统计量=15.818,P<0.001),采用Dunnett’s t3进行两两比较,结果显示,绿地环境中,2012-2014年白纹伊蚊密度差异不显著[3.65、2.18和2.73只/(天·观察点),均P>0.05];住区环境白纹伊蚊密度[分别为11.91和42.02只/(天·观察点)均高于绿地环境[分别为3.65、2.18和2.73只/(天·观察点)],差异有统计学意义(F=7.704,P值均<0.001)。住区环境中,2014年白纹伊蚊密度升高[42.02 vs. 11.91只/(天·观察点)],差异有统计学意义(F=7.704,P<0.001)。见表 2。
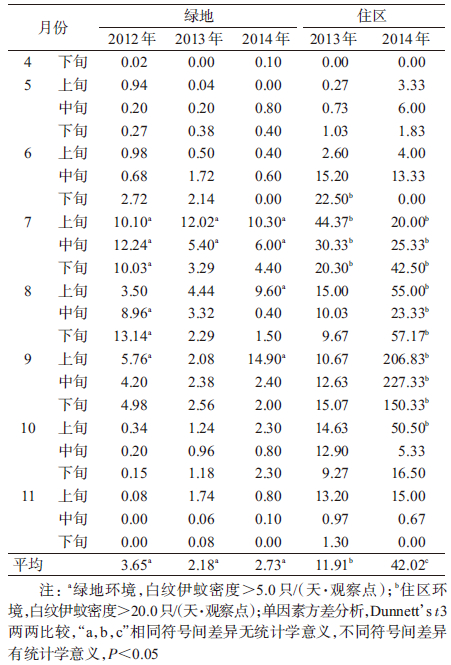
2014年白纹伊蚊侵害整体上呈现出上升的趋势,尤其在住区环境,白纹伊蚊构成比达94.69%,密度达42.02只/(天·观察点)。住区环境3个观察点比较显示,上海城区旧式里弄环境白纹伊蚊密度最高,2014年9月10日白纹伊蚊密度达到924只/(天·观察点);高层住宅与老式别墅白纹伊蚊密度相对较低(图 1)。

|
| 图 1 2014年上海中心城区3个住区环境白纹伊蚊密度变化趋势 |
2. DNA条形码序列分析:共获得90个白纹伊蚊DNA 条形码序列,各序列之间遗传距离<0.019,表明采集的上海市白纹伊蚊高度近缘,为同一种或亚种。进化树分析表明,伊蚊属各蚊种间有较高的亲缘关系(遗传距离<0.25),即使如此,种间仍然存在差别,且表现出根据不同蚊种聚类的特性,以及同一蚊种还存在按照地理位置聚类的特性。我国上海市的白纹伊蚊与2005-2011年在非洲地区开展白纹伊蚊基因分型研究时所取得的DNA 条形码序列虽然在进化树上仍属同一支系(clade),彼此之间的遗传距离甚大,其种内遗传距离甚至大于某些埃及伊蚊、刺扰伊蚊(Aedes vexans Meigen,1830)、曼氏伊蚊(Aedes Mcintoshi)、朝鲜伊蚊(Aedes koreicus Edwards,1971)、日本伊蚊(Aedes japonicus Theobald,1901)、东乡伊蚊(Aedes togoi Theobald,1907)等的种间差异。此外,本研究所采集的白纹伊蚊与2011年采自日本的东乡伊蚊、2014年采自肯尼亚的曼氏伊蚊,其亲缘关系也较2005-2011年在非洲所采集的白纹伊蚊近(图 2)。

|
| 注:●本研究所取得的序列; ■为2011年采自日本的东乡伊蚊序列;□为2014年采自肯尼亚的曼氏伊蚊序列;▲为2005-2011年非洲白纹伊蚊序列 图 2 不同区域白纹伊蚊线粒体细胞色素氧化酶(COⅠ)DNA序列进化树 |
住区与绿地是城市居民生活休闲的主要场所,因此,本研究采用的观察数据为上海市2012及2013年每年224 d连续性成蚊观察及2014年每周1次观察频率的成蚊数据,观察频率及观察点设置方面均优于常规的观察方法,数据可真实详细地反映近年蚊虫在观察点的侵害状况。本研究显示,白纹伊蚊为上海中心城区优势蚊种,尤其在住区环境中白纹伊蚊已成为最主要的侵害蚊种,且2014年的数据显示白纹伊蚊的侵害程度有上升的趋势。
市中心住区环境的白纹伊蚊整体构成比为78.53%,是优势种群。徐仁权等[10, 11]2002及2007年在上海地区开展的研究亦显示,居民区是白纹伊蚊分布的主要环境之一。针对住区环境白纹伊蚊侵害明显的现象,原因分析主要与居民区的环境特点及白纹伊蚊的生态习性有关,同时亦较强的受到住区人类活动的影响。白纹伊蚊属于“容器积水”孳生型蚊虫[6],居民区往往富含适宜白纹伊蚊孳生的小型积水,尤其是夏季雨水过后各类容器积水均可以产生大量的白纹伊蚊孳生地;此外白纹伊蚊喜刺吸人血,住区人口密集,为白纹伊蚊繁殖提供大量的潜在血源,陆宝麟[6]亦指出白纹伊蚊是半野生的蚊种,在居民点及居民点附近的竹林树林等生境大量发生,但远离居民点同样孳生条件下为数很少。
2014年上海市住区环境白纹伊蚊构成比明显高于2013年,主要与旧式里弄观察点白纹伊蚊密度大幅上升有关,该观察点在高峰期(2014年9月10日)白纹伊蚊密度达到924只/(天·观察点),且连续6个观察日(45 d观察区间)白纹伊蚊密度均高于200只/(天·观察点),这在城区环境中较为罕见。对于该异常现象进行现场调查,该观察点周边有一旧房拆迁工地,拆迁后未及时清除造成的洼地积水可能是造成白纹伊蚊异常升高的重要原因。
白纹伊蚊最敏感的观察方法是人诱停落法[12],本研究中采用的CO2捕蚊机法对白纹伊蚊的敏感性相对略低,敏感性较低的方法依然显示白纹伊蚊较高的构成比,说明在自然情况下,白纹伊蚊密度和构成比高于本研究中获取的数据,其叮咬骚扰及潜在的蚊媒传染病传播风险可能更大。
2014年以来,美洲多个国家相继发生寨卡病毒感染病例,其中以巴西疫情尤为严重,欧洲、亚洲、大洋洲等地区也有输入病例报告,截止2016年2月5日,至少在非洲、亚洲、美洲地区的45个国家有寨卡病毒的传播证据[13, 14]。寨卡病毒1947年首次在乌干达恒河猴中发现,1952年一项回顾性血清学调查确定人类可以感染该病毒,随后1954 年在尼日利亚检出人类感染该病的病原体[15, 16]。根据基因型别分为非洲型和亚洲型,本次美洲流行的为亚洲型[17, 18]。
寨卡病毒是一种蚊媒病毒,伊蚊叮咬是寨卡病毒病的主要传播途径,媒介蚊种在传播寨卡病毒中的重要性依次为埃及伊蚊、白纹伊蚊,非洲伊蚊和黄头伊蚊也可能传播该病毒[15, 16]。作为长期自然选择的必然结果,媒介昆虫的种类与其所能携带或传播的病原体有着一定的协同进化关系,并表现出病原体相对于昆虫媒介有着较高的专一性。本研究显示此次采集于上海市的白纹伊蚊与非洲白纹伊蚊的相似性高于同处亚洲地区的白纹伊蚊[19, 20, 21],而本次流行于南美各国的寨卡病毒为亚洲型,尚不知这种差别能否导致寨卡病毒传入我国时被有效中止;亦不知这种与非洲伊蚊所表现出来的更为近缘的关系会不会促进一些非洲优势流行的病原体在传入我国时得以有效传播,我们仅能从这样一些线索上谨慎推知本次出现在南美诸国及其他疫区的寨卡病毒可能不会在我国上海等地造成较大的流行。例如,作为寨卡病毒的最有效昆虫媒介,埃及伊蚊仅在我国海南、广东以及云南等省的局部地区分布[22],当前在全国并未发现及上海地区亦无分布[23];白纹伊蚊在寨卡病毒传播和流行过程中的宿主地位次于埃及伊蚊[15, 16],且本次流行的寨卡病毒为亚洲型,而采自上海地区的白纹伊蚊与非洲白纹伊蚊的基因型更为接近,按照前述病原体与其传播媒介的专一性原则,这种差别可能会影响宿主对病毒的传播能力。当前我国已有输入性寨卡病毒病病例报道[24, 25],同时大部分地区天气已回暖,虑及寨卡病毒在全球的流行形势,不仅需要加强对寨卡病毒的可能传播媒介的种群动态进行密切监控,同时尚需要防控其他蚊媒疾病,上海中心城区需要大幅降低蚊虫侵害,尤其是白纹伊蚊的侵害程度。
利益冲突 无| [1] Paupy C,Delatte H,Bagny L,et al. Aedes albopictus,an arbovirus vector:From the darkness to the light[J]. Microbes Infect,2009,11(14/15):1177-1185. DOI:10.1016/j.micinf.2009.05.005. |
| [2] Benedict MQ,Levine RS,Hawley WA,et al. Spread of the Tiger:Global risk of invasion by the mosquito Aedes albopictus[J]. Vector Borne Zoonotic Dis,2007,7(1):76-85. DOI:10.1089/vbz.2006.0562. |
| [3] Paupy C,Ollomo B,Kamgang B,et al. Comparative Role of Aedes albopictus and Aedes aegypti in the Emergence of Dengue and Chikungunya in Central Africa[J]. Vector Borne Zoonotic Dis,2010,10(3):259-266. DOI:10.1089/vbz.2009.0005. |
| [4] Musso D,Nilles EJ,Cao-Lormeau VM. Rapid spread of emerging Zika virus in the Pacific area[J]. Clin Microbiol Infect,2014,20(10):O595-596. DOI:10.1111/1469-0691.12707. |
| [5] 龚道方,周红宁. 中国登革热重要媒介白纹伊蚊的研究进展[J]. 中国媒介生物学及控制杂志,2009,20(6):607-610. Gong DF,Zhou HN. Progress in Dengue fever important vector Aedes albopictus in China[J]. Chin J Vector Biol Control,2009,20(6):607-610. |
| [6] 陆宝麟. 中国动物志昆虫纲第八卷双翅目蚊科(上)[M]. 北京:科学出版社,1997:38-55,433. Lu BL. China fauna:Insect (Ⅷ):Diptera:Culicidae (1st part)[M]. Beijing:Science Press,1997:38-55,433. |
| [7] 柳支英,陆宝麟. 医学昆虫学[M]. 北京:科学出版社,1990:127-129. Liu ZY,Lu BL. Medical entomology[M]. Beijing:Science Press,1990:127-129. |
| [8] 廉国胜,柯明剑,伍碧梅,等. 国境口岸常见11种蚊虫DNA条形码分子鉴定研究[J]. 中国国境卫生检疫杂志,2015,38(3):171-175. DOI:10.16408/j.1004-9770.2015.03.006. Lian GS,Ke MJ,Wu BM,et al. DNA-barcode based molecular identification for eleven common mosquito species at frontier ports[J]. Chin J Front Health Quarant,2015,38(3):171-175. DOI:10.16408/j.1004-9770.2015.03.006. |
| [9] 庞雄飞,尤民生. 昆虫群落生态学[M]. 北京:中国农业出版社,1996:12-35. Pang XF,You MS. Insect community ecology[M]. Beijing:China Agriculture Press,1996:12-35. |
| [10] 徐仁权,蔡恩茂,徐友祥,等. 上海地区白纹伊蚊监测与药物防制研究[J]. 中国媒介生物学及控制杂志,2003,14(4):256-260. DOI:10.3969/j.issn.1003-4692.2003.04.005. Xu RQ,Cai EM,Xu YX,et al. Surveillance on larvae and adult of Aedes albopictus and the study of insecticide in Shanghai[J]. Chin J Vector Biol Control,2003,14(4):256-260. DOI:10. 3969/j.issn.1003-4692.2003.04.005. |
| [11] 徐仁权,徐宏,李洪宝,等. 居民区控制登革热媒介蚊虫的方法与效果探索[J]. 中国媒介生物学及控制杂志,2008,19(4):287-290. DOI:10.3969/j.issn.1003-4692.2008.04.004. Xu RQ,Xu H,Li HB,et al. Study on the control methods and control effect of the vector of dengue fever in residential area in Shanghai[J]. Chin J Vector Biol Control,2008,19(4):287-290. DOI:10.3969/j.issn.1003-4692.2008.04.004. |
| [12] Marquetti MC,Valdes V,Aguilera L. Entomological surveillance over Aedes aegypti and other culicids in Ciudad de La Habana,Cuba 1991-1996[J]. Rev Cubana Med Trop,2000,52(2):133-137. |
| [13] Pan American Health Organization/World Health Organization. Epidemiological Update:Neurological syndrome,congenital anomalies and Zika virus infection[EB/OL]. Washington,D. C.:PAHO/WHO,2016.[2016-02-05]. http://www.paho.org/hq/index.php?option=com_docman&task=doc_view&Itemid=270&gid=32879&lang=en. |
| [14] World Health Organization. Zika strategic response framework & joint operations plan[EB/OL].[2016-02-19]. http://apps.who.int/iris/bitstream/10665/204420/1/ZikaResponseFramework_JanJun 16_eng.pdf?ua=1. |
| [15] Faye O,Freire CCM,Iamarino A,et al. Molecular evolution of Zika virus during its emergence in the 20th century[J]. PLoS Negl Trop Dis,2014,8(1):e2636. DOI:10.1371/journal.pntd. 0002636. |
| [16] Lanciotti RS,Kosoy OL,Laven JJ,et al. Genetic and serologic properties of Zika virus associated with an epidemic,Yap State,Micronesia,2007[J]. Emerg Infect Dis,2008,14(8):1232-1239. DOI:10.3201/eid1408.080287. |
| [17] Mlakar J,Korva M,Tul N,et al. Zika virus associated with microcephaly[J]. N Engl J Med,2016,374:951-958. DOI:10.1056/NEJMoa1600651. |
| [18] Ioos S,Mallet HP,Goffart IL,et al. Current Zika virus epidemiology and recent epidemics[J]. Med Mal Infect,2014,44(7):302-307. DOI:10.1016/j.medmal.2014.04.008. |
| [19] Kamgang B,Ngoagouni C,Manirakiza A,et al. Temporal patterns of abundance of Aedes aegypti and Aedes albopictus (Diptera:Culicidae) and mitochondrial DNA analysis of Ae. albopictus in the Central African Republic[J]. PLoS Negl Trop Dis,2013,7(12):e2590. DOI:10.1371/journal.pntd.0002590. |
| [20] Raharimalala FN,Ravaomanarivo LH,Ravelonandro P,et al. Biogeography of the two major arbovirus mosquito vectors,Aedes aegypti and Aedes albopictus (Diptera:Culicidae),in Madagascar[J]. Parasit Vectors,2012,5:56. DOI:10.1186/1756-3305-5-56. |
| [21] Mousson L,Dauga C,Garrigues T,et al. Phylogeography of Aedes (Stegomyia) aegypti (L.) and Aedes (Stegomyia) albopictus (Skuse) (Diptera:Culicidae) based on mitochondrial DNA variations[J]. Genet Res,2005,86(1):1-11. DOI:10.1017/S0016672305007627. |
| [22] 国家卫生计生委. 国家卫生计生委办公厅关于印发寨卡病毒病诊疗方案的通知[EB/OL].[2016-02-03]. http://www.nhfpc.gov.cn/yzygj/s3593g/201602/e7c1402a03024501ad8f036de346c145.shtml. |
| [23] 周正斌,吕山,张仪,等. 上海市蚊媒种类、分布及其病原[J]. 中国媒介生物学及控制杂志,2015,26(1):28-32. DOI:10.11853/j.issn.1003.4692.2015.01.007. Zhou ZB,Lyu S,Zhang Y,et al. Mosquito species,distribution and their pathogens in Shanghai,China[J]. Chin J Vector Biol Control,2015,26(1):28-32. DOI:10.11853/j.issn.1003.4692. 2015.01.007. |
| [24] 廖勇,曾志笠,胡国良,等. 中国大陆首例输入性寨卡病毒病病例调查分析[J]. 中华流行病学杂志,2016,37(5):593-596. DOI:10.3760/cma.j.issn.0254-6450.2016.05.001. Liao Y,Zeng ZL,Hu GL,et al. Survey of the first case of Zika virus disease in the mainland of China[J]. Chin J Epidemiol,2016,37(5):593-596. DOI:10.3760/cma.j.issn.0254-6450. 2016. 05.001. |
| [25] 董选军,孙继民,楼莲青,等. 中国大陆第三例输入性寨卡病毒病病例调查分析[J].中华流行病学杂志,2016,37(5):597-599. DOI:10.3760/cma.j.issn.0254-6450.2016.05.002. Dong XJ,Sun JM,Lou LQ,et al. Survey of the third Zika virus disease case in the mainland of China[J]. Chin J Epidemiol,2016,37(5):597-599. DOI:10.3760/cma.j.issn.0254-6450. 2016. 05.002. |