 2018, Vol. 29
2018, Vol. 29 



b University of Chinese Academy of Sciences, Beijing 100049, China;
c CAS Key Laboratory of Separation Science for Analytical Chemistry, Dalian Institute of Chemical Physics, Chinese Academy of Sciences, Dalian 116023, China
Biomolecular interactions between metabolites and proteins are important to cellular physiological processes. In addition, covalent and enzymatic modifications of proteins (also known as post translational modification, PTM) play a crucial role in disease progression [1]. Identification of interacting small metabolite and protein is a key step ahead to therapeutic drug discovery. Generally, enzymes catalyze different types of metabolites under different certain conditions. Inversely, these metabolites serve as substrates to control the enzymatic activity of proteins. On the other hand, metabolites may act as transcription co-factors, signaling ligands and signaling hormones without affecting the enzymatic activity of interacting proteins. Current studies have reported that about 20% proteins bind with metabolites [2]. Recently, several metabolite-protein interactions have been reported by different groups (Table 1). Wan and her colleagues have reported that cholesterol functionally binds to the nuclear receptor ERRα and regulates downstream signaling pathway [3]. The results demonstrate that cholesterol and its biosynthetic pathway regulate the ERRα activity in cells [3]. However, it is still not clear whether cholesterol universally regulates ERRα transcriptional activity in disease progression. The molecular mechanism of how metabolite regulates the protein enzymatic activity needs to be investigated in detail in large scale biological studies.
|
|
Table 1 List of identified metabolite-protein interactions. |

To open a new frontier in small metabolite and protein interactions, a variety of applicable approaches should be employed. Previously, various groups have reported different approaches to analyze small metabolite and protein interactions in in vitro, in silico, and in vivo models [9]. For example, Li et al. identified some hydrophobic metabolites interacting with ergosterol biosynthetic proteins and protein kinases by using affinity protein purification and mass spectrometry analysis in yeast [7]. They successfully demonstrated both known and novel proteinhydrophobic metabolite interactions in vivo through co-immunoprecipitation. However, this approach is very limited only to stable bindings. Most of the non-covalently interacted metabolites might lose binding during the process of affinity purification, due to weak binding force.
With the introduction of ABPP, the lacunae in the small metabolite and protein interaction study has been addressed. This can explore the covalently and non-covalently interacting proteins by specifically designed chemical probes of metabolites [10-12]. This chemical probe is specifically designed to retain the original bioactivity of metabolite, and coupled with a reporter tag that can be used to enrich the target proteins. Then, these enriched target proteins can be detected by gel-based or MS-based proteomics. This method can directly identify the interacting protein-metabolite in vivo under various physiological conditions in conjunction with advances in the sensitivity, resolution, and throughput of analytical platforms. To globally map the direct interactions between cholesterol and proteins, Cravatt and colleagues designed clickable and photoreactive sterol probes and quantified about 250 interacting proteins by MS-based proteomics [11]. For proteomic mapping of non-covalently interacting proteins, a photoreactive group is tagged or ligated to the metabolite probes. However, this approach may yield few non-specific interactions and fail to detect some sterol-protein interactions if the cholesterol undergoes modifications either by diazirine or ester [11]. In parallel, the same group has globally mapped both known and previously unannotated lipid-binding proteins by using a set of lipid-based chemical proteomic probes in mammalian cells [12]. Although these lipid-protein interactions constitute a vital landscape for ligand development, this sometimes gives false-negative outcomes due to structural modifications imposed by the photo-crosslinking and alkyne groups [12]. In another study, Wang et al. have profiled a list of bile acid (BA) interacting proteins through multiple BA-based chemical probes in combination with stable-isotope labeling by amino acids in cell culture (SILAC) based high throughput quantitative proteomics approach [13]. Here, some sets of novel interacting BA-proteins have been traced. These findings will pave a new direction for functional studies of BA signaling and BA-mediated pathways in human disease metabolism [13].
To identify the covalent interactions between proteins and metabolites, alkyne and azide group linked metabolites could be included and enriched with the target proteins. Recently, Song and colleagues have reported that cholesterol modification of Smoothened (Smo) protein is required for Hedgehog signaling pathway [8]. Several covalently modified cholesterol-proteins have also been identified by MS-based ABPP [8]. The post-translational modification of ADP-ribosylation is known to be involved in cellular homeostasis and stress. To prove this mechanism biochemically has still remained a challenge to us. To detect the ADP-ribosylated proteins, Hang and colleagues designed an alkyne-adenosine analog, N6-propargyl adenosine which is metabolically incorporated to mammalian cells to analyze the proteomic of ADP-ribosylated proteins [14].
Metabolite-protein interaction plays an important role in cellular homeostasis. But, so far only a small fraction of interaction networks could be identified. Based on its importance and wide applications in various samples, ABPP may be recommended as a powerful approach to dissect out the interactions or modifications (both covalent and non-covalent) between metabolites and proteins. The more challenging now is if the proteins which interact with the metabolites share common specific fractions of proteomic space or not. Metabolites, in nature, possess ring or ring like structures, such as bile acids, which reversibly interact with or covalently modify the proteins.
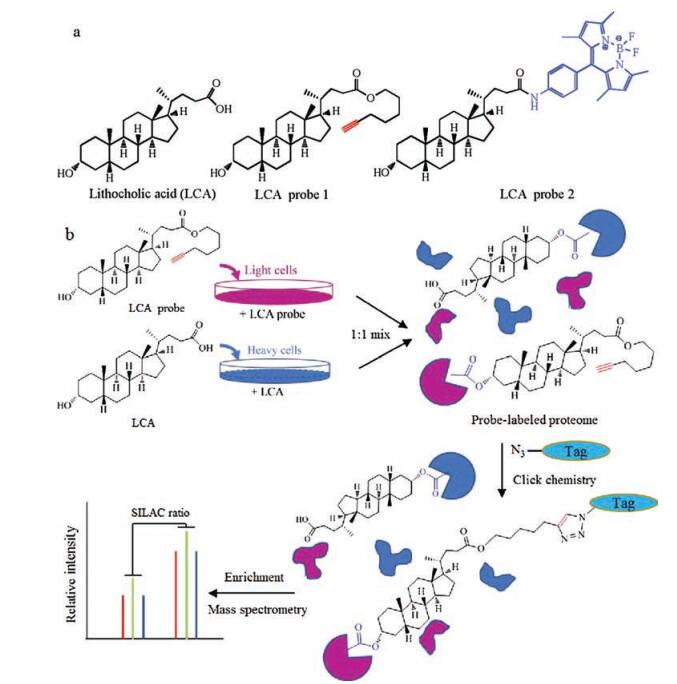
Here, Lithocholic acid (LCA) is an example of MS-based ABPP (Fig. 1). The hydroxyl moiety of the LCA probe forms an ester bond with carboxyl groups of proteins, and the azide group on the side chain would allow the click chemistry reaction. The covalent interaction of LCA probe and protein will provide a more comprehensive portrait of modified proteins. Consequently, LCA probe-modified proteins would be processed for quantitative identification by SILAC-based quantitative proteomic approach. However, it is noticeable that there are still some challenges to apply MS-based ABPP to complex biological systems. The structural modification should minimally perturb the biophysical properties of metabolites when selecting chemical probes. We should also mention that endogenous esterases might cleave the ester linkage of esterified alkyne modifications, which could reduce the sensitivity of proteomic profiling. Of course it is entirely inevitable that there are lots of false-negative outcomes and that chemical probes may not detect certain metabolite-protein interactions that are impaired by the modifications to the structure of metabolites.

|
Download:
|
| Fig. 1. Identification of covalently modified proteins by LCA probe. (a) Structures of LCA active chemical probes. (b) Schematic representation of the procedure to detect and identify LCA probe-modified protein(s). | |
{kind=link}
Here, we have updated on the interactions of active endogenous metabolites and proteins. The current literature suggests that combination of quantitative proteomics with activity-based protein profiling is a powerful and robust tool to systematically map the network of metabolite-protein interaction. This approach may pave a new direction to develop new chemical compound(s) or molecule(s) in metabolic disease and malignant cancer progression, and suggest metabolites to be the novel potential biomarkers in the personalized medicine.
AcknowledgmentsThe metabolite-protein interaction research in the Piao's Lab is supported by the National Natural Science Foundation of China (No. 81672440), Innovation Program of Science and Research from the DICP, CAS (No. DICP TMSR201601) and the 100 Talents Program of Chinese Academy of Sciences.
| [1] |
D.J. Leeming, A.C. Bay-Jensen, E. Vassiliadis, et al., Biomarkers 16 (2011) 193-205. DOI:10.3109/1354750X.2011.557440 |
| [2] |
X. Li, M. Snyder, Bioessays 33 (2011) 485-489. DOI:10.1002/bies.v33.7 |
| [3] |
W. Wei, A.G. Schwaid, X. Wang, et al., Cell Metab. 23 (2016) 479-491. DOI:10.1016/j.cmet.2015.12.010 |
| [4] |
W. Gilbert, B. Mullerhi, Proc. Natl. Acad. Sci. U. S. A. 56 (1966) 1891-1898. DOI:10.1073/pnas.56.6.1891 |
| [5] |
P. Magotti, I. Bauer, M. Igarashi, et al., Structure 23 (2015) 598-604. DOI:10.1016/j.str.2014.12.018 |
| [6] |
R.A. Heyman, D.J. Mangelsdorf, J.A. Dyck, et al., Cell 68 (1992) 397-406. DOI:10.1016/0092-8674(92)90479-V |
| [7] |
X. Li, T.A. Gianoulis, K.Y. Yip, M. Gerstein, M. Snyder, Cell 143 (2010) 639-650. DOI:10.1016/j.cell.2010.09.048 |
| [8] |
X. Xiao, J.J. Tang, C. Peng, et al., Mol. Cell 66 (2017) 154-162. DOI:10.1016/j.molcel.2017.02.015 |
| [9] |
G.X. Yang, X. Li, M. Snyder, Methods 57 (2012) 459-466. DOI:10.1016/j.ymeth.2012.06.013 |
| [10] |
B.F. Cravatt, A.T. Wright, J.W. Kozarich, Annu. Rev. Biochem. 77 (2008) 383-414. DOI:10.1146/annurev.biochem.75.101304.124125 |
| [11] |
J.J. Hulce, A.B. Cognetta, M.J. Niphakis, S.E. Tully, B.F. Cravatt, Nat. Methods 10 (2013) 259-264. DOI:10.1038/nmeth.2368 |
| [12] |
M.J. Niphakis, K.M. Lum, A.B. Cognetta, et al., Cell 161 (2015) 1668-1680. DOI:10.1016/j.cell.2015.05.045 |
| [13] |
S. Zhuang, Q. Li, L. Cai, C. Wang, X. Lei, ACS Cent Sci. 3 (2017) 501-509. DOI:10.1021/acscentsci.7b00134 |
| [14] |
N.P. Westcott, J.P. Fernandez, H. Molina, H.C. Hang, Nat. Chem. Biol. 13 (2017) 302-312. DOI:10.1038/nchembio.2280 |