 2003, 14 (3): 257-265
2003, 14 (3): 257-265


2. 中国科学院地理科学与资源研究所,北京100101
2. nstitute of Geographic Sciences and Natural Resources, CAS, Beijing 100101
1965年,de Wit首次根据气象因子对作物影响的机理,利用数值方法求解作物高产上限,从而提出了作物生长的数值模拟理论[1]。此后,随着计算手段的不断提高,作物生长数值模拟得以迅速发展,20世纪80年代以来,国际上英、美及荷兰等国家陆续开发了大量作物数值模式,其中有代表性的如荷兰的SUCROS[2]、ARID CROP[3],英国的AFRCWHEAT[4],以及美国的CERES[5]模式。进入90年代后,作物生长数值模式作为下垫面子模型开始与区域气候模式相耦合,并结合卫星遥感形成综合的地气耦合系统,其中有代表性的为1995年欧盟主持开发的WOFOS T模型[6]。
在中国,竺可桢在20世纪60年代首次从农业气候学角度论证了气候对农业生产力的影响[7],黄秉维提出了作物高产的光温生产潜力定义[8]。高亮之、金之庆、黄耀等则在中国首次进行了农业气象数值模拟的一些初步工作,1989年完成了“苜蓿农业气象模拟模型 (ACFAMOD)”[9],并在国内首次建立起具有一定推广面积的RCSODS作物生长优化决策系统。此后王石立与王馥棠[10]、黄策与王天铎[11]、冯定原[12]等相继进行了水稻和小麦的数值模式研究。进入20世纪90年代后,吴连海、韩湘玲等[13]首次将作物生长数值模式引入气候资源研究中,自此作物模型在生产力分析中得以广泛应用[14-18]。最近,作物生长模型作为重要的地表生态系统过程,开始考虑大气环境因素对作物的影响[19,20],并且与生物地球化学模型相耦合[21-27]。
应当指出,国际上的作物生长数值模型,在建立模式前均进行了大量的生理生态实验,并且即使在模型完成后,也还在不断收集各地不同国家的相关光合生理生态资料,进行模型验证以使模型不断完善。然而迄今为止,中国作物生长数值模拟中的一些作物参数,甚至叶片光合速率等最基本参数也经常直接引用国外相关文献。众所周知,中国目前主要冬小麦品种的光合速率与国外相比存在一定差异,并且国内不同年代的主要品种间也存在一定差异,模型中最基本参数的直接引用会引起对作物生长数值模拟可靠性的怀疑。因此,作者利用Licor-6200光合测定仪对不同年代的主要冬小麦品种叶片光合作用速率进行了大量测定,并确定了各自的光合作用曲线,从而为未来数值模型在实际生产中的应用以及与区域气候模式耦合提供了最基本的模型参数。
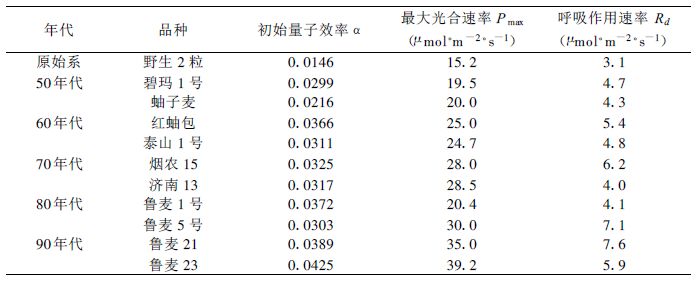
1 实验场地及方法实验在中国科学院禹城综合试验站 (36°50′N,116°40′E) 及山东农业大学实验基地 (36°16′N,117°16′E) 冬小麦标本园进行,山东农业大学实验基地是黄淮海地区冬小麦品种繁育及栽培主要基地,标本园内栽种了1949年以来黄淮海地区不同年代的200多个冬小麦品种。选取黄淮海地区各年代推广面积最大和最具代表性的两个主要品种作为研究对象,20世纪50年代:碧玛1号和蚰子麦;20世纪60年代:红蚰包和泰山1号;20世纪70年代:烟农15和济南13;20世纪80年代:鲁麦1号和鲁麦5号;20世纪90年代:鲁麦21和鲁麦23。此外,标本园中种植了冬小麦原始系野生2粒,在20世纪初期中国尚未进行育种研究,因此研究中将此原始系品种作为1900年左右的一种对比品种进行测定。
1998年10月中旬播种,水肥管理均适宜,各冬小麦品种长势良好,于1999年4月27日小麦抽穗期利用美国LICOR公司生产的Licor-6200便携式光合作用测定仪对冬小麦的叶片光合作用速率进行了系统测定。Licor-6200便携式光合测定仪每次可以同时采集叶片光合作用速率、光合有效辐射光量子通量密度、CO2浓度、温度等多项指标存贮在机内芯片中形成一个数据文件,大量测定后将结果一次打印全部输出。测定中通过遮光方法测定了光强渐变时叶片光合速率变化情况。1999~2000年及2000~2001年分别在该标本园进行了两年重复实验,共得到数万条原始数据。
2 实验结果及数值敏感分析 2.1 不同年代冬小麦叶片的光合参数利用Licor-6200光合作用测定仪可以非常迅速地测定光合速率随光强变化情况,每测定一个品种大约15 min左右,因此在测定过程中温度基本保持不变。测定过程中水肥及温度处于适宜状态,因此光量子通量密度成为叶片光合速率的主要影响因子,此时叶片净光合作用速率 (Pn) 可表示为
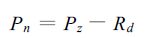
|
(1) |
式中Pz为总光合速率;Rd为暗呼吸作用速率。Pz与光量子通量密度 (I) 之间关系通常表示为[22]
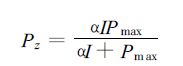
|
(2) |
式中α,Pmax分别是初始量子效率和最大光合速率;利用最小二乘法原理拟合光合作用的光响应曲线,即通过迭代使拟合残差平方和δ达到最小。
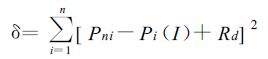
|
(3) |
式中Pni为第i个样本的实际净光合速率,n为样本数。由式 (1) 和式 (2) 可以看出,由α、Pmax和Rd三个参数即可决定光合作用的光响应曲线,因此将其作为光合作用特征参数。
将实测数据代入式 (1)、(2) 进行迭代运算,得到冬小麦光合作用曲线 (图 1)。对全部品种分别确定各自的光响应曲线,即可得到不同年代冬小麦叶片光合作用特征参数 (表 1)。

|
|
| 图 1. 不同品种冬小麦叶片光合作用光响应曲线模拟 (a) 野生2粒;(b) 鲁麦1号;(c) 鲁麦23 | |
|
|
表 1 不同品种冬小麦叶片光合作用特征参数 |
2.2 冬小麦叶片光合特征参数的演化过程
为了进一步揭示不同年代品种冬小麦光合特征参数随时间的演变过程,将20世纪50年代品种时间设为1955年,60年代品种设为1965年,并依次类推,从而得到光合特征参数随时间的变化规律 (图 2)。应当指出:不同年代间的品种种植为一个时段,而非一个时刻,因而用直方图表达各时段的参数更接近实际,但由于各年代选取了两个最主要的品种,因而难以直接用直方图表示品种光合特性演变过程。实际上,即使可以采用直方图表示,其演变规律分析结果与以上处理方法得到的结果显然也是完全等同的。

|
|
| 图 2. 冬小麦叶片光合作用参数随时间变化规律 (a)初始量子效率α;(b)最大光合速率]Pmax;(c)呼吸作用速率Rd | |
利用线性回归、指数回归以及双曲线回归等多种拟合方法,探讨α及Pmax随时间变化情况。从总体来看,初始量子效率α随时间演变以线性增长形式拟合效果最好 (图 2a),而最大光合速率Pmax则呈一种指数增长形式 (图 2b),呼吸速率Rd变化规律则不太明显 (图 2c)。初始量子效率α和最大光合速率Pmax随时间变化函数可以表示为
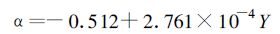
|
(4) |
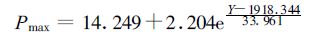
|
(5) |
式中Y为年份。由式 (1) 和式 (2) 可以看出,单纯从数学公式上也可以发现α和Pmax是决定光合作用曲线的最主要的两个参数,Rd并不能改变光响应曲线的形式而仅能改变曲线在坐标系中的上下位移,并且R d是一个较小量 (表 1),对整个曲线的影响相对α和Pmax而言要小得多。从整体趋势来看,作为影响光响应曲线的最主要的两个特征参数,α和Pmax在1949年以来有了较大幅度的提高,这也是黄淮海地区作物生产力逐步提高的最直接的根本原因。
2.3 叶片光合参数对光合日总量计算结果影响的数值分析近年来,中国作物生长数值模拟研究中α和Pmax两个最重要的光合特征参数大量直接引用H.v an Keulen在20世纪80年代的研究结论[3],即α=15 ×10-6g ·J-1,Pmax=3.5 g·m-2·h-1,按目前国际单位制α=0.0741,Pmax=22.1 μmol·m-2·s-1。为了将此特征参数与国内冬小麦特征参数进行对比,在图 2a和图 2b中分别读取模拟线上对应于1 95 5年的α和Pmax值作为5 0年代的光合特征参数理论值,R d取平均值,依此类推,即可得到不同年代的光响应曲线 (图 3)。图中清楚表明:荷兰小麦品种与中国不同年代冬小麦品种相比,叶片光合作用均存在较大差异。在辐射强度较低的情况下,荷兰小麦光合能力较强;然而在辐射强度较大的环境下,荷兰20世纪80年代小麦光合作用能力低于我国20世纪80年代和90年代小麦光合能力。由此可见,时至今日中国模型大量直接引用国外参数是不恰当的。

|
|
| 图 3. 不同年代冬小麦光合作用光响应曲线 | |
冬小麦逐日光合作用累积对最终产量形成至关重要。因此,为了进一步说明参数引用不当可能带来的误差,有必要进一步对光合日总量进行数值分析。如果计算净光合作用日总量,在积分运算中暗呼吸要分为生长呼吸和维持呼吸,因子增多而无法比较单纯由特征参数带来的光合日总量差异;由于Rd相对而言较小,特别是光合作用较强的典型晴天,相对值更小,因此此处计算总光合作用的日总量。设水、肥、温度等条件均保持适宜状态,在黄淮海地区典型晴天,一天中任一时刻直接辐射S(ω) 和散射辐射D(ω) 可表示为[19]
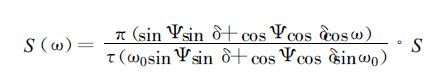
|
(6) |
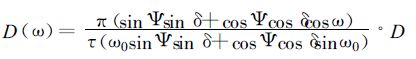
|
(7) |
式中Ψ为地理纬度;ω为时角;ω0=arccos (-tg Ψtgδ) 为日没时角;τ为日长;δ为赤纬,由左大康Fourier分析结果进行计算[14,16-20]; S为直接辐射日总量;D为散射辐射日总量。任一时刻的直接光合有效辐射S *(ω)=0.42S (ω) 及散射光合有效辐射D *(ω)=0.53D (ω),对于波段为400~700 nm的光合有效辐射,1 J光合有效辐射所含光量子数为4.6 μmol[29]。
在太阳光线ns方向上G函数可以表示为[19]
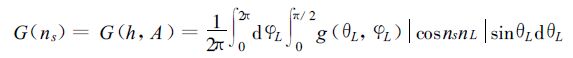
|
(8) |
式中h为太阳高度角,A为太阳方位角,g (θL,φL) 为叶方位角为φL、叶倾角为θL的叶倾角分布函数,nL为叶片法线方向的单位矢量,cosnsnL为太阳光线方向和叶片法线方向夹角的余弦,直射光穿透函数τs(L,ns) 和散射光穿透函数τd(L) 为
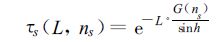
|
(9) |
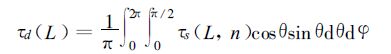
|
(10) |
其中τs(L,n) 为倾角θ和方位角φ所决定矢量方向的透过函数。累积叶面积指数 (L-1)~L层次内、倾角为θL、方位角为φL叶片上的直接光合有效辐射为
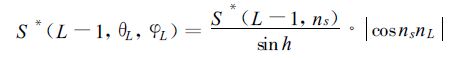
|
(11) |
在太阳高度角为h,方位角为A时考虑了一次散射过程后的累积叶面积指数L处水平面散射辐射强度为
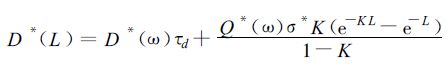
|
(12) |
式中σ*=(ρ*+τ*)/2,其中ρ*为叶片光合有效辐射反射系数,τ*为叶片光合有效辐射透射系数,根据各向同性的辐射理论,在累积叶面积指数 (L -1)~L层次内、倾角为θL、方位角为φL的叶片收到的散射辐射量与叶方位角无关,可以表达为
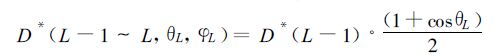
|
(13) |
将太阳时角ω时刻,累积叶面积系数L、叶倾角θL、方位角为φL叶片上的光合有效辐射Q*(ω,L,θL,φL) 代入单叶总光合作用式 (2),得到此叶片净光合速率Pz (ω,L,θL,φL),那么时角ω时刻群体总光合速率为
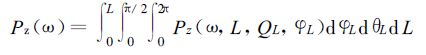
|
(14) |
对时角ω积分即可得到总光合作用日总量。
首先设定数值分析的基值,模拟地点为黄淮海地区中部济南,日期为2001年5月15日,典型晴天,直接辐射日总量为18 MJ·m-2,散射辐射日总量为6 MJ·m-2,冬小麦叶面积指数为5。实际上由于品种不同,同一日期叶面积显然不可能完全相同,但数值分析的目的是为了揭示仅由参数引用不当造成的模型计算误差,因此进行这种理论假设是必要的。
数值分析结果 (图 4a) 清楚表明:在黄淮海地区典型晴天,荷兰品种与中国冬小麦品种之间群体光合作用日变化形式存在一定差异,中午时段20世纪90年代品种鲁麦23的群体光合能力大于荷兰品种,而其它时刻则刚好相反。从光合日总量而言,荷兰品种的光合作用积分值与鲁麦23大体相当,即荷兰品种在低光强下的较大的模拟值与高光强下的较小的模拟值造成的计算误差正好大体抵消,因此得到的结论是如果用荷兰品种参数,那么最终光合日总量不会存在较大差异。但是从科学角度来讲,同时存在两种误差,仅由于误差相互抵消而认为模型正确显然是不允许的。并且,如果模型的研制目的是为了揭示瞬时光合作用并进行调控,必然导致一天中绝大多数时间均存在较大误差。

|
|
| 图 4. 品种特征参数对冬小麦群体光合的影响 (a) 不同品种群体光合日变化过程;(b) 特征参数对光合日总量影响的数值分析 (图中等值线单位:g·m -2·d -1) | |
光合特征参数α和Pmax对光合日总量影响的数值分析 (图 4b) 表明:当α较小时,Pmax增加对光合日总量的影响较小;但是当α较大时,光合日总量随Pmax增加迅速增大。
3 结论与讨论(1) 利用Licor-6200便携式光合作用测定仪,对1949年以来不同历史年代的冬小麦叶片光合作用进行了大量测定,拟合了不同年代12个典型品种的叶片光合作用光响应曲线,首次揭示了黄淮海地区冬小麦叶片光合特征参数随时间演变规律。数值分析表明:20世纪80年代的荷兰品种与中国20世纪80年代及90年代的冬小麦品种相比,虽然从光合日总量而言,荷兰品种光合作用日积分值与中国黄淮海地区代表品种鲁麦23大体相当,但两者之间群体光合作用日变化形式存在一定差异。从科学角度来讲,同时存在两种误差,仅由于误差相互抵消而认为模型正确显然是不允许的,并且在揭示瞬时光合作用时,引用荷兰参数必然导致一天中绝大多数时间均存在较大误差。
(2) 作物模型已经成为地学领域生产力分析中的主要研究手段,模型的基本参数对模型计算结果至关重要,直接引用国外模型参数需要十分慎重。地学界广泛引用的世界粮农组织气象组广为推荐的FAO生产力模型中的相关参数比本文提及的荷兰参数取值更小,无论在低光强还是在高强下光合作用值均小于中国20世纪90年代水平,以致中国的现实生产力甚至超过FAO计算的生产潜力,存在很大的不合理性,对此作者曾对FAO原有基本参数进行了必要的订正[17]。而本文研究则表明:直接引用国外参数即便在光合日总量值计算正确的情况下,也可能在中间过程中存在较大的计算错误,因此,模型参数应当以实测为准。
(3) 目前各学科广泛交叉,植被作为重要的地球表层生态因子,已经开始与地球生物化学模型相耦合[21-26],并且正在试图与大气模式相耦合[30]。本文所得到的大量参数,为未来建立综合模式提供了最基本的模型参数。此外,本文首次揭示了叶片光合作用的演变过程,由此可以断定作物生产潜力也必然存在一种演变过程,因此,在本文研究基础上,有可能进一步揭示中国近代作物生产力的演变过程及未来演变趋势。作为人口大国,这一研究将对中国的粮食安全提供重要理论依据。此外,气候变化对农业影响成为目前国内外研究热门课题,在研究中对未来生产力进行预测时,一般假定目前作物特性保持不变,模拟未来几十年后的气候情景下生产力变化状况。然而,本文清楚地揭示出,自1949年到目前20世纪90年代,随着品种的进化,叶片光合能力已经得到大幅度提高,而且品种进化已使叶片光合能力提高了不止一倍。因此,在以后有关气候变化对作物生产力影响的研究中应当进一步考虑品种进化的影响。本文提供的大量基础数据,为设计品种进化情景提供了重要的科学依据。
致谢 毕建杰及樊广华二位同志协助进行了大量实验测定工作,中国科学院禹城综合试验站科研人员对该工作给予一定帮助,谨此致谢。| [1] | de Wit C T. Photosynthesis of Leaf Canopies. PUDOC Wangeningen, 1965, 1: 1–12. |
| [2] | Penning de Vries F W T, van Laar H H. Eds Simulation of plant growth and crop production Simulation Monographs. PUDOC Wangeningen, 1982: 23–126. |
| [3] | van Keulen H. Simulation of water use and herbage growth in arid regions-a reevaluation and further development of the model "ARID CROP". Agric Syst, 1981, 6: 159–193. DOI:10.1016/0308-521X (81)90001-9 |
| [4] | Porter J R. Comparison of the wheat simulation models AFRCWHEAT2, CERES-Wheat and SWHEAT for nonlimiting conditions of crop growth. Field Crop Research, 1993, 33: 131–157. DOI:10.1016/0378-4290(93)90098-8 |
| [5] | Grant R F. Simulation of carbon assimilation and partitioning in maize. Agron. J, 1989, 6: 193–220. |
| [6] | Supit I, Hooijer A A. System description of the WOFOST 6.0 crop simulation model implemented in CGMS. JRC, 1994: 3–144. |
| [7] | 竺可桢. 论我国气候的几个特点及其与粮食作物的关系. 地理学报, 1964, 30, (1): 1–13. |
| [8] | 黄秉维. 中国农业生产潜力--光合潜力.中国科学院北京农业生态系统试验站编.农业生态环境研究. 北京: 气象出版社, 1989: 17-24. |
| [9] | 高亮之, 金之庆, 著. 水稻栽培计算机模拟优化决策系统. 北京: 中国农业科学技术出版社, 1992: 1-80. |
| [10] | 王石立, 王馥棠. 春小麦生长简化模拟模式研究. 应用气象学报, 1991, 2, (3): 294–300. |
| [11] | 黄策, 王天铎. 水稻群体物质生长过程的计算机模拟. 作物学报, 1986, 12, (1): 1–8. |
| [12] | 冯定原. 水稻生长和产量形成的数值模式. 南京气象学院学报, 1987, 10, (2): 201–211. |
| [13] | 吴连海, 韩湘玲. 冬小麦生产力估算方法研究. 自然资源学报, 1991, 6, (1): 80–87. |
| [14] | 刘建栋, 傅抱璞, 林振山. 冬小麦生产潜力数值模拟. 地理研究, 1998, 17, (1): 56–65. |
| [15] | 刘建栋, 傅抱璞, 金之庆. 应用ARID CROP模型对中国黄淮海地区冬小麦气候生产力的数值模拟研究. 自然资源学报, 1997, 12, (3): 282–287. |
| [16] | 刘建栋, 于强, 傅抱璞. 黄淮海地区冬小麦光温生产潜力数值模拟研究. 自然资源学报, 1999, 14, (2): 169–174. |
| [17] | 刘建栋, 周秀骥, 于强. FAO生产力模型中基本参数的修正. 自然资源学报, 2001, 16, (3): 240–247. |
| [18] | 刘建栋, 周秀骥, 于强. 温度对夏玉米光合生产力影响的数值模拟研究. 应用气象学报, 2002, 13, (4): 398–405. |
| [19] | 刘建栋, 周秀骥, 于强. O3, CO2浓度及太阳光谱变化对作物光合作用影响的数值模拟研究. 气象学报, 2002, 60, (6): 715–721. |
| [20] | 刘建栋, 周秀骥, 于强. 近地层在气臭氧对水稻光合作用影响的数值模拟. 环境科学学报, 2003, 23, (3): 1–7. |
| [21] | Zhang Yu, Li Changsheng, Zhou Xiuji. A simulation model linking crop growth and soil biogeochemistry for sustainable agriculture. Ecological Modelling, 2002, 151: 75–108. DOI:10.1016/S0304-3800(01)00527-0 |
| [22] | Liu Jiandong, Zhou Xiuji, Yu Qiang. Numerical analysis of the source-sink alternation of composite global warming potential of the rice paddy ecosystem in the Yangtze River delta. Science in China, Series D, 2003, 46, (4): 385–396. |
| [23] | 刘建栋, 周秀骥, 于强. 长江三角洲稻田生态系统综合增温潜势源汇交替的数值分析. 中国科学, D辑, 2003, 33, (2): 105–113. |
| [24] | 刘建栋, 周秀骥, 于强. 长江三角洲稻田CH4及N2O排放规律的数值模拟. 自然科学进展, 2001, 11, (9): 960–965. |
| [25] | 刘建栋, 周秀骥, 丁国安. 农田生态系统N2O排放的数值模拟研究. 环境科学, 2002, 23, (6): 36–39. |
| [26] | Liu Jiandong, Zhou Xiuji, Zhang Yu. Numerical simulation of methane emissions from rice paddy fields in the Yangtze River Delta of China. Acta Meteorologica Sinica, 2001, 15, (1): 1–11. |
| [27] | 刘建栋, 周秀骥, 王建林, 等. 稻田CH4排放的农业气象数值模拟研究. 应用气象学报, 2001, 12, (4): 409–418. |
| [28] | 索恩利J H M. 植物生理的数学模型. 王天铎译, 北京: 科学出版社, 1983. 107-143. |
| [29] | 董振国. 光量子光照度与总辐射的关系.见:中国科学院北京农业生态系统实验站编.农业生态系统能量物质交换. 北京: 气象出版社, 1987: 115-122. |
| [30] | 薛永康, 曾凡荣. 一个植被模式的检验和它在陆气相互作用的应用. 大气科学, 1998, 22, (4): 575–586. |