 2012, Vol. 28
2012, Vol. 28


2. Department of Geology, Miami University, Oxford, OH 45056
2. Department of Geology, Miami University, Oxford, OH 45056, USA
近年来,国际上关于海相泥质烃源岩有机质保存机理领域的研究取得了重大进展。传统观点认为,海相泥质烃源岩中有机质的保存主要受环境因素控制,即取决于表层海水中所产生的有机质数量与其在水体中沉积和埋藏过程中被氧化速度之间的平衡,因此,诸如缺氧条件、高有机质产率、快速埋藏等条件可有效地抑制有机质的氧化和微生物对其的降解,从而有利于有机质的保存(Zhang et al.,2007;Zhu et al.,2007)。正因为此,富有机质黑色页岩的形成长期以来被认为是缺氧沉积环境的证据(Demaison and Moore,1980)。然而,对控制有机质保存的机制并不清楚。近年来的研究发现,许多沉积在现代大陆边缘的海相富有机质沉积物中,有时可发现生物扰动构造(Kakuwa,1996)、相当轻的硫同位素比值(δ34S)以及较高的C/S比(Kajiwara et al.,1994),说明了一种次氧化(suboxic)的海底水环境(Suzuki et al.,1998),且沉积速率也不高,因此,必然存在着另外的机制控制了海相泥质岩中有机质的保存。
目前,越来越多的证据证明,海相泥质烃源岩中的有机质是通过溶解在水中的分子规模的碳有机质以化学吸附的形式被吸附到矿物表面上。Bader et al.(1970) 早在20世纪70年代初就提出了黏土矿物对溶解有机质的吸咐在有机质保存中的作用,但就其机理,以及沉积物中有机质保存与矿物比表面积之间的关系,只是到了最近才引起了海洋地球化学家的广泛注意(Keil and Hedges,1993; Mayer,1994a; Hedges and Keil,1999; Hedges and Oades,1997)。研究发现,现代海洋大陆边缘富有机质沉积物矿物比表面积(MSA)与总有机碳含量(TOC)之间存在着很强的相关关系(Mayer,1999,1994b; Ransom et al.,1998;Bergamashi et al.,1997;Bock and Mayer,2000); TEM图象也显示了有机碳在矿物表面的不规则吸附包壳(Ransom et al.,1998;Bishop et al.,1992;Ransom et al.,1997);矿物粒度分离表明,现代海相富有机质沉积物中只有不到10%的有机质呈颗粒状与矿物共生。现代海岸沉积物中被矿物吸附而稳定存在的沉积有机质,一旦被解吸,很快就被喜氧细菌所降解(Keil and Hedges,1993;Bergamashi et al.,1997)。这说明了矿物表面对有机分子的吸附是海相黑色页岩中有机质保存的主要因素。不稳定的有机组分通过单个矿物层间显微场所中的吸附而使其与外界环境隔离而得以保存(Bader et al.,1970; Gordon and Millero,1985; Heinrichs and Sugai,1993; Bishop and Phillip,1994; Collins et al.,1995; Salmon et al.,2000; Keil and Hedges,1993;Mayer,1994a;Hedges and Keil,1995)。最近,Kennedy et al.(2002) 注意到,2:1型层状蒙脱石的外表面积类似于其它黏土矿物和细粒矿物,但是结构层内的表面积可比外表面积高1到2个数量级。高岭石和石英粉砂通常有<10m2g-1的比表面积,而蒙脱石可达到900m2g-1的比表面积。因此,若沉积物中含有中等含量的蒙脱石,其总的矿物比表面积中蒙脱石的内表面积将是主要贡献者。其内表面提供了吸附有机碳组分的重要场所,其吸附的TOC含量可大于15%(Kennedy et al.,2002)。因此,有机质的蒙脱石层内吸附为有机质的有效保存提供了一个良好的解释机制。然而,目前关于微生物能否降解蒙脱石层间吸附有机质以及降解的程度等尚没有任何实验数据的支撑。本文试图通过人工合成含有层间吸附有机质的蒙脱石,利用海洋和湖泊沉积物中常见的降解有机质的微生物对其进行降解实验,据此探讨有机质的蒙脱石层间吸附在沉积物埋藏过程中对有机质保存的贡献。
1 实验思路通常蒙脱石矿物的d(001) 层间距为12Å,当蒙脱石结构层内吸附有机物后,该层间距将增大,可达到17~18Å。微生物主要通过2种途径破坏蒙脱石层间吸附有机质的保存:(1) 分泌有机酸,直接降解层间有机质;(2) 厌氧条件下通过铁的还原破坏蒙脱石结构,使有机质无法继续保存,释放后被其他微生物消耗。
按照第一种途径,如果微生物能将有机物很好的降解,则经微生物降解作用后的蒙脱石d(001) 层间距将较作用前合成样品减小,并随着降解程度的增加,其层间距可逐渐恢复到原始间距,或略大,同时,作用后矿物中有机质含量下降,溶液中有机质含量增加。按照第二种途径,若微生物将蒙脱石结构破坏,则有机质释放至溶液中,作用后溶液中有机质含量增加。
因此,实验过程采用人工合成层间吸附有机物的蒙脱石,在实验室模拟海洋和湖泊底部沉积物温度、pH值及氧气供应等条件下,用微生物对合成样品进行降解。通过对作用前后的样品及溶液进行蒙脱石矿物结构层间距、矿物中有机质含量、溶液中微生物数量、溶液中有机质含量等参数的测定和对比,探讨蒙脱石层间吸附作用在抵御微生物对其中有机质降解过程中的贡献。
2 实验材料的制备 2.1 黏土矿物和有机物的选择本研究中黏土矿物选择绿脱石(nontronite,富铁蒙脱石)。有机质选择半胱氨酸(cysteine)和甲苯(toluene)。半胱氨酸为生物生长所需的一种重要氨基酸,也存在于电子转移蛋白中(Zhang et al.,2007),微生物对其个体的降解作用较强,作为自然界有机成分的代表;甲苯是大量存在于土壤和沉积物中的一种致癌烃,多种细菌可以在有碳氢化合物的环境下降解甲苯(Zhang et al.,2007)。
2.2 层间吸附半胱氨酸和甲苯的绿脱石的合成 2.2.1 绿脱石样品制备本研究中选用0.5~0.2μm黏土颗粒。(1) 用重力沉降和离心进行分离,并自然风干;(2) 将样品置于微波灭菌设备中灭菌5min;(3) 取少量样品放入LB培养基培养,30℃,48h,无光有氧培养,无细菌生长,说明样品无菌。
2.2.2 半胱氨酸绿脱石与甲苯绿脱石的合成(1) 半胱氨酸绿脱石的合成
① 单离子黏土矿物(homoionic clay)的合成:将2g绿脱石NAu-2与200mL 1M CuCl2混合,室温下置于摇床内12h,后将绿脱石与CuCl2混合物离心,取上清夜,与新制备的1N CuCl2混合。如此重复3次,将该混合物用蒸馏水稀释洗涤,直至Cl-被洗净(AgNO3检测无沉淀)。
② 半胱氨酸加入绿脱石黏土矿物层间结构:将2g单离子绿脱石加入100mL 0.05M半胱氨酸溶液中,混合液室温下置于摇床24h,后将其离心,用蒸馏水洗涤10次。半胱氨酸的加入量由始末溶液中半胱氨酸浓度的差异测算。
合成后样品的各项参数见表 1。FTIR测试显示,在半胱氨酸绿脱石中,出现了有机物的红外吸收特征峰(图 1)。
|
|
表 1 有机质-蒙脱石合成样品参数表 Table 1 Parameters for composed samples of OM-smectite |

|
图 1 半胱氨酸绿脱石合成样品(C0-0) FTIR测试图 Fig. 1 FTIR analysis of composed sample of smectite with cysteine(C0-0) |
(2) 甲苯绿脱石的合成
① 十六烷基三甲基溴化铵(HDTMA)-黏土混合物的合成:将20mg/mL十六烷基三甲基溴化铵(HDTMA)加入10mg/mL黏土矿物溶液中,并用磁力搅拌器混合均匀,混合4h后,用去离子蒸馏水洗涤直至无其它盐离子。
② 甲苯加入绿脱石黏土矿物层间结构:将0.10gHDTMA-黏土混合物加入装有25mL蒸馏水的25mL离心管,之后在离心管中加入12μL甲苯/甲醇(toluene/methanol,4.6μL/7.4μL)溶液,室温下将离心管置于摇床摇18h,再离心,并用蒸馏水反复洗涤5次,制成样品。甲苯的加入量由始末溶液中甲苯浓度的差异测算。
合成后样品的各项参数见表 1。FTIR测试显示,在甲苯绿脱石中,出现了有机物的红外吸收特征峰(图 2)。

|
图 2 甲苯绿脱石合成样品(T0-0) FTIR测试图 Fig. 2 FTIR analysis of composed sample of smectite with toluol(T0-0) |
实验菌种选择恶臭假单胞杆菌(Pseudomonas putida)和腐败希瓦氏菌(Shewanella putrefaciens CN32)2种细菌。它们均为海洋和湖泊沉积物中的主导微生物,前者有较强的有机质降解能力,后者为铁的还原菌,厌氧代谢过程中能将蒙脱石结构中的Fe(Ⅲ)还原为Fe(Ⅱ)(Zhang et al.,2007)。上述菌种购回为冻干粉,按如下步骤培养:
(1) 假单胞菌培养
① 将菌种玻璃管灼烧,后用无菌水淋浇,至爆裂;② 注入600μL无菌水,震荡至悬浮;③ 将液体吸出,注入80mL LB中(盛放于250mL灭菌三角瓶中);④ 将三角瓶置于摇床,37℃,180r·m-1,有氧培养24~48h,至溶液变混浊;⑤ 培养后未被使用的希瓦氏菌与等体积甘油混合于1.5mL离心管,并冷冻于-80℃冰箱内保存;⑥ 以后使用时先将冷冻的菌种置于室温解冻,然后将3mL解冻的菌液加入50mL TSB培养基中培养,步骤同③~⑤。
(2) 希瓦氏菌培养
① 将菌种玻璃管灼烧,后用无菌水淋浇,至爆裂;② 注入600μL无菌水,震荡,至悬浮;③ 将液体吸出,注入80mL TSB中(盛放于250mL灭菌三角瓶中);④ 将三角瓶置于摇床,26℃,180r·m-1,有氧培养48~72h,至溶液变混浊;⑤ 培养后未被使用的希瓦氏菌与等体积甘油混合于1.5mL离心管,并冷冻于-80℃冰箱内保存;⑥ 以后使用时先将冷冻的菌种置于室温解冻,然后将3mL解冻的菌液加入50mLTSB培养基中培养,步骤同③~⑤。
3 实验方案和过程如前所述,沉积物中的微生物可以通过有机酸的生成从而降解有机质和/或将晶体结构中Fe(Ⅲ)还原成Fe(Ⅱ)而使蒙脱石溶解,从而释放有机质,2种方法均可实现对有机质保存的破坏。因此,在本实验中,预期假单胞菌主要对有机质进行直接降解,希瓦氏菌主要将蒙脱石结构中Fe(Ⅲ)还原为Fe(Ⅱ),从而通过对反应后最终溶液及矿物成分的分析确定有机质在黏土矿物层间的保存情况。
3.1 假单胞菌降解有机质实验(1) 活化后的假单胞菌培养至停滞期,将含菌培养基溶液注入1.5mL离心管中,4000r·m-1,离心10min,倒去上部LB,加入等量含碳源碳酸盐缓冲液 反复离心洗涤2次;
(2) 向离心管中加入1.5mL 含碳源碳酸盐缓冲液,与菌种混合均匀;
(3) 血球计数板对希瓦氏菌进行计数;
(4) 将半胱氨酸绿脱石和甲苯绿脱石紫外灭菌30min,之后如下表成分进行配置(表 2),其中PC0-1及PT0-1为空白对照;实验中缓冲液为寡营养,不含有机质,迫使生物只能从绿脱石层间获得电子,以完成新陈代谢作用。
|
|
表 2 恶臭假单胞杆菌降解实验样品成分表 Table 2 Composition of samples degraded by Pseudomonas putida |
(5) 以上各组样品均用250mL灭菌三角瓶盛装,纱布封口置于摇床,36℃,180r·m-1,有氧培养10d,其间保证氧气充足供应,以利于假单胞菌生长;
(6) 培养10d后将溶液注入1.5mL离心管中,14,000g,离心5min,分别回收上清液及黏土矿物,上清液编号依次为YPC2-2,YPT2-2,YPC0-1,YPT0-1;
(7) 将回收的糊状黏土矿物均匀盛放于培养皿中,置于无菌操作间自然风干。
3.2 希瓦氏菌还原Fe(Ⅲ)实验由于希瓦氏菌只有在无氧的条件下才能对Fe(Ⅲ)进行还原,因此本实验的主要作用在严格厌氧条件下进行。
(1) 活化后的假单胞菌培养至停滞期,将含菌培养基溶液注入1.5mL离心管中,4000r·m-1,离心10min,倒去上部TSB,加入等量含碳源碳酸盐缓冲液反复离心洗涤2次;
(2) 向离心管中加入1.5mL 含碳源碳酸盐缓冲液,与菌种混合均匀;
(3) 血球计数板对假单胞菌进行计数;
(4) 将配置好的含碳源的碳酸盐缓冲液分装入4个厌氧瓶中,用抽换气设备将内部气体成分控制为N2/CO2:80/20;
(5) 将换好气的厌氧瓶灭菌,121℃,30min;
(6) 将半胱氨酸绿脱石和甲苯绿脱石紫外灭菌30min,之后在厌氧箱内,如下表成分进配置(表 3),其中SC0-1及ST0-1为空白对照;
|
|
表 3 希瓦氏菌还原绿脱石层间Fe(Ⅲ)实验样品成分表 Table 3 Composition of samples degraded by Shewanella putrefaciens CN32 |
(7) 将培养瓶口封紧,置于摇床,26℃,180r·m-1,无氧培养10d;
(8) 培养10d后将溶液注入1.5mL离心管中,14,000g,离心5min,分别回收上清液及黏土矿物,上清液编号依次为YSC1-2,YSC0-1,YST0-1;
(9) 将回收的糊状黏土矿物均匀盛放于培养皿中,置于无菌操作间自然风干。
4 实验结果为了分析恶臭假单胞杆菌对有机物的降解和腐败希瓦氏菌对Fe(Ⅲ)的还原,我们通过细胞计数、粉晶X射线衍射、傅立叶转换红外线光谱和DTNB法检测实验前后流体中细菌数量、半光胺酸含量和矿物晶体结构的变化。
4.1 实验前后流体中细菌数量的变化2种细菌作用于半胱氨酸绿脱石反应后活细菌细胞含量(血球板计数)及以往相同条件下所做希瓦氏菌作用于甲苯绿脱石及绿脱石单矿物的实验数据如表 4。
|
|
表 4 细胞计数结果表 Table 4 Results from cell counting |
(1) 与半胱氨酸绿脱石作用后,假单胞菌数量没有明显变化(均为5×109左右),希瓦氏菌的数量有所增加(从1×108 cells·mL-1到8×108 cells·mL-1);
(2) 希瓦氏菌作用实验 半胱氨酸作用中,绿脱石单独体系中细胞数量明显减少(1×108 ~4.5×105 cells·mL-1);在甲苯作用中,希瓦氏菌数量没有增加,均有明显减少,且与半胱氨酸作用结果不同,细菌作用于甲苯绿脱石的减少量远大于绿脱石单独体系(分别为1×108~5×105 cells·mL-1和2×108~2.1×107 cells·mL-1)。
4.2 实验前后矿物晶体结构参数变化半胱氨酸绿脱石及甲苯绿脱石结构的变化由粉晶X射线衍射实验进行测试,衍射实验结果如表 5。
|
|
表 5 样品粉晶X射线衍射数据表 Table 5 Results from XRD analysis of samples |
(1) 假单胞菌与半胱氨酸绿脱石作用
① 空白对照(PC0-1) 与原始样品(C0-0) 峰形差异不大,峰形反应其结晶程度不好。空白对照d(001) 层间距略为增大(13.301~14.153Å),并出现3个新的峰值:4.149Å,2.820Å,1.933Å相应2θ分别为:21.400°,31.700°,45.480°。
② 假单胞菌作用后样品(PC1-1/PC1-2) 结晶程度明显高于原始样品和空白对照组,且出现较明显的新峰值如下:
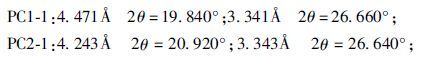
|
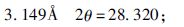
|
作用后d(001) 层间距略有减小,PC1-1从13.301Å减小为13.104Å,PC2-1减小为12.511Å。
③ 不同浓度假单胞菌作用后的样品(PC1-1/PC1-2) 波形差异不大,黏土矿物成分相同,浓度较小的PC2-1(1×108cells·mL-1)d(001) 层间距(12.511Å)小于浓度较大的PC1-1(1×1011cells·mL-1)(13.104Å)。但总体上看d(001) 间距变化不大。
(2) 假单胞菌与甲苯绿脱石作用
空白对照(PT0-1) 、原始样品(T0-0) 及作用后样品波形整体差异不大,矿物结晶程度较高,一强峰出现在2θ为6.000°左右,2θ为40.00°以后,不再出现明显波峰。
① 空白对照d(001) 层间距较原始样品略为减小(15.019~14.198Å),并出现1个新的峰值:2.800Å,2θ=31.940°。
② 假单胞菌作用后样品(PT1-1/PT2-1) 与空白对照相比,出现新峰值如下:
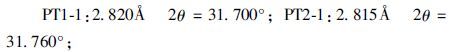
|
作用后d(001) 层间距减小,PT1-1从15.019Å减小为14.525Å,PC2-1减小为14.290Å。
③ 不同浓度假单胞菌作用后的样品(PT1-1/PT1-2) 波形差异不大,黏土矿物成分相同,浓度较小的PT2-1(1×108cells·mL-1)d(001) 层间距(14.290Å)小于浓度较大的PT1-1(1×109cells·mL-1)(14.525Å)。但总体上看d(001) 间距变化也不大。
(3) 希瓦氏菌与甲苯绿脱石作用
空白对照(ST0-1) 、原始样品(S0-0) 及作用后样品波形整体差异不大,矿物结晶程度较高,一强峰出现在2θ为6.000°左右,2θ为40.00°以后,不再出现明显波峰。
① 空白对照作用后d(001) 层间距略为减小,ST0-1从15.019Å减小为14.718Å;
② 作用后d(001) 层间距减小,SC1-1从15.019Å减小为14.383Å;
③ 有细菌作用的层间距减小量比空白对照多0.335Å。
4.3 实验前后样品中有机质的变化利用傅立叶转换红外线光谱测试分析样品中的有机质,结果如下:
(1) 假单胞菌作用
① 作用于半胱氨酸绿脱石实验中,原始样品(C0-0) 与细菌作用样品(PC2-1) 差异不大,无新峰值出现;空白对照(PC0-1) 的指纹区出现新峰值681.95cm-1(图 3)。

|
图 3 假单胞菌降解半胱氨酸绿脱石作用前后FTIR测试图 Fig. 3 FTIR analysis of composed sample of smectite with cysteine degraded by Pseudomonas putida |
② 作用于甲苯绿脱石实验中,原始样品(T0-0) ,细菌作用样品(PT2-1) 和空白对照(PT0-1) 三者差异不大,较强峰分别出现在3565cm-1,2926cm-1,2854cm-1,1640cm-1,1479cm-1,1022cm-1,679cm-1(图 4)。

|
图 4 假单胞菌降解甲苯绿脱石作用前后FTIR测试图 Fig. 4 FTIR analysis of composed sample of smectite with toluol degraded by Pseudomonas putida |
(2) 希瓦氏菌作用
① 作用于半胱氨酸绿脱石实验中,空白对照(SC0-1) 与原始样品差异不大,有菌作用后样品(SC1-1) 与原始样品(T0-0) 在1500~500cm-1 间出现一系列新峰值:1462cm-1,1418cm-1,1315cm-1,1118cm-1(图 5)。

|
图 5 希瓦氏菌对半胱氨酸绿脱石作用前后FTIR测试图 Fig. 5 FTIR analysis of composed sample of smectite with cysteine degraded by Shewanella putrefaciens CN32 |
② 作用于甲苯绿脱石实验中,空白对照(ST0-1) 与原始样品差异不大,有菌作用样品(ST1-1) 与原始样品相比,在1500~500cm-1 间出现一系列新峰值:1470cm-1,1418cm-1,1365cm-1,1315cm-1,1266cm-1,1122cm-1(图 6)。

|
图 6 希瓦氏菌对甲苯绿脱石作用前后FTIR测试图 Fig. 6 FTIR analysis of composed sample of smectite with toluol degraded by Shewanella putrefaciens CN32 |
分别取与假单胞菌和希瓦氏菌与半胱氨酸绿脱石作用的样品上清夜,用DTNB法测试,结果表明:
(1) 总体半胱氨酸浓度均较低,约为10-5mol·L-1;
(2) 希瓦氏菌作用后的半胱氨酸浓度大于假单胞菌作用(分别为1.95×10-5mol·L-1,1.50×10-5mol·L-1)。
5 讨论 5.1 微生物对蒙脱石层间吸附有机质保存和破坏的影响 5.1.1 恶臭假单胞杆菌的作用假单胞杆菌降解半胱氨酸绿脱石实验中,原始样品作用前后层间距总体差别不大,且细菌作用与否层间距变化不大,作用后层间距略有减小(图 7)。其中,半胱氨酸绿脱石实验中(C0-0,PC0-1,PC1-1) ,无菌对照组层间距比原样增大,可能是绿脱石在反应过程中从周围吸收了部分水分,可能导致层间距变大。从实验结果看,细菌作用后层间距的改变量(0.197Å)远小于注入有机物的改变量(3.301Å),说明假单胞菌无法降解层间有机质。

|
图 7 恶臭假单胞菌实验各样品d(001) 层间距比较柱状图 Fig. 7 Correlation of d(001) interlayer spaces of samples before and after experiment by Pseudomonas putida |
假单胞菌降解甲苯绿脱石实验中,无菌对照和有菌作用层间距均减小(图 7),但量都较小(0.821Å,0.494Å),远小于有机质注入后层间距的改变量(5.019Å)。且细菌作用后的层间距大于无菌对照组,说明假单胞菌对层间甲苯的降解几乎没有作用(图 3)。FTIR图象也显示,假单胞菌作用前后的样品无明显变化,依然出现半胱氨酸,甲苯的特征峰(图 3、图 4)。同时,假单胞菌作用于半胱氨酸绿脱石后的细胞数量没有变化(均为5×109cells·mL-1,表 4),说明细菌很难从绿脱石层间获取有机物以供自身生长。溶液中有机物浓度的DTNB测试表明,有细菌作用的溶液中有机质含量很少(1.50×10-5mol·L-1),而且少于无菌空白对照(10.02×10-5mol·L-1),这表明有机质-绿脱石集合体在无菌作用下也能释放一定量的有机质到溶液中。释放的一部分有机质可能会被假单胞菌消耗,但不足以维持细菌正常迅速生长,所以在假单胞菌作用的实验中,细菌量没有变化且有机质含量少于空白对照。
5.1.2 腐败希瓦氏菌的作用希瓦氏菌作用于半胱氨酸绿脱石实验中,原始样品作用前后层间距总体差别不大,且细菌作用与否层间距变化不大(图 8),作用后层间距略有减小(15.019~14.383Å),减小量略大于空白对照(15.019~14.718Å),但仍显示大于绿脱石层间距(12.000~13.000Å)。另外,希瓦氏菌作用后溶液颜色变为浅绿色,说明有新的Fe(Ⅱ)出现。以上特征表明希瓦氏菌可能对绿脱石结构有一定的破坏作用,导致溶液中出现新的F(Ⅱ),且作用后层间距比无菌对照小,又远大于绿脱石。

|
图 8 腐败希瓦氏菌菌实验各样品d(001) 层间距比较柱状图 Fig. 8 Correlation of d(001) interlayer spaces of samples before and after experiment by Shewanella putrefaciens CN32 |
同时,希瓦氏菌作用于半胱氨酸绿脱石后的细胞数量增多(1×108~8×108cells·mL-1),进一步说明希瓦氏菌已获得营养物质,从而使自身生长。
以上结果表明,希瓦氏菌不仅利用还原Fe(Ⅲ)获得电子,也消耗从半胱氨酸绿脱石释放到溶液中的有机质从而实现代谢。因此,希瓦氏菌在无氧条件下对绿脱石层间结构有破坏作用,且消耗半胱氨酸能力较强。无氧条件下,希瓦氏菌以还原Fe(Ⅲ)为主,对有机质的降解能力有限。
5.1.3 2种不同菌种作用结果的比较从甲苯-绿脱石层间距的减小的绝对量上看,假单胞菌为0.494Å,而希瓦氏菌为0.636Å;从与空白对照比较的相对值看,假单胞菌较空白对照减少0.327Å,而希瓦氏菌为0.335Å。由此表明,希瓦氏菌对层间距的减小能力更强。
FTIR也显示希瓦氏菌作用后的矿物质中出现一系列新的特征峰,表明矿物结构发生了变化(图 5、图 6)。
从菌种作用后溶液中半胱氨酸浓度看,希瓦氏菌作用后溶液中半胱氨酸绝对浓度(1.95×10-5mol·L-1)大于假单胞菌作用后溶液中半胱氨酸绝对浓度(1.5×10-5mol·L-1)。同时希瓦氏菌增长倍数是假单胞菌的近8倍,因此,希瓦氏菌从有机质-绿脱石中释放有机物和获取有机物的能力远大于假单胞菌本身。
综合以上因素考虑,假单胞菌从绿脱石层间获取有机物能力很弱,希瓦氏菌由于破坏了层间结构,从而使较多有机质释放到溶液中,也使自身顺利生长。
5.2 不同有机物对生物降解的影响半胱氨酸存在于溶液中时,可作为电解质,因此可以显著的促进Fe(Ⅲ)还原量。在我们的实验中,释放到溶液中的半胱氨酸可以作为电解质发挥作用,因此可以促进Fe(Ⅲ)的还原过程。半胱氨酸同时也是生命活动必须的氨基酸,因此,溶液中释放的半胱氨酸可能促进了生物的生长进而促进了Fe(Ⅲ)还原。实验中(表 4),假单胞菌作用后数量不变而希瓦氏菌还有很大增长,而希瓦氏菌作用于绿脱石单独体系时细胞数量有明显减少(1×108~4.5×105cells·mL-1)。
相反,甲苯对生物生长无益甚至有害,因此,甲苯能抑制细菌活动,同时也对抑制Fe(Ⅲ)的还原起着重要作用。实际上,在我们的数据中,该体系的希瓦氏菌活细胞减少量十分明显(2×108~5×105cells·mL-1)而无菌空白对照的活细胞减少量相对较小(2×108~2×107cells·mL-1)(表 4)。
5.3 蒙脱石层间吸附对有机质的保存作用从假单胞菌对有机质的降解情况看,有机物的蒙脱石层间吸附对其的保存有利,有效的阻碍了有机质被分解。无氧条件下,希瓦氏菌对蒙脱石的作用以还原Fe(Ⅲ)为主。希瓦氏菌对矿物结构的破坏是影响有机物保存的主要因素。然而,由于细菌对矿物晶体结构的破坏作用能力有限,故其对有机质的降解能力也是有限的。由此可见,有机质的蒙脱石层间吸附可以是有机质保存的重要方式之一。
5.4 实验结果的地质意义众所周知,黏土矿物在埋藏成岩过程中,随着埋深的增加,蒙脱石逐渐转变成伊/蒙混层矿物,进而在深埋条件下转变为伊利石。高岭石随着埋深的增加将逐渐消失。因此,大部分含油气盆地中浅埋藏的烃源岩中,其黏土矿物的组成中,蒙脱石和高蒙脱石结构层比例的伊/蒙混层矿物在含量上占有绝对优势。如渤海湾盆地,埋深在1500m以上的古近系烃源岩中,蒙脱石和高蒙脱石结构层比例的伊/蒙混层矿物的含量占80%左右;松辽盆地埋深在1200m以上的白垩系烃源岩中,蒙脱石和高蒙脱石结构层比例的伊/蒙混层矿物的含量占50%左右;北部湾盆地埋深在1500m以上的古近系烃源岩中,高蒙脱石结构层比例的伊/蒙混层矿物的含量可达60%(赵杏媛等,1995)。由此可以看出,蒙脱石和高蒙脱石结构层比例的伊/蒙混层矿物在原始烃源岩矿物组成中占有很大的比重。由于矿物表面吸附的有机质较易遭微生物降解,而黏土矿物中又只有蒙脱石的晶体结构才能提供层间吸附有机质的空间和条件,故蒙脱石的层间吸附有机质对于有机质的保存具有重要意义。实验结果较好地证明了蒙脱石层间吸附对有机质的保存作用,这些被吸附在蒙脱石结构层中的有机质,由于较好地抑制了埋藏早期微生物对其的降解作用,从而能较好地保存到中深埋藏环境中。随着埋深的增加,温度的升高,蒙脱石在逐渐向伊利石转化过程中,由于晶体结构的变化,会相应地排出吸附在其层间的有机质,同时,受温度的影响,将促使这些有机质转化为烃类。因此,有机质的蒙脱石层间吸附保存是烃源岩中有机质保存的有效机制之一。
6 结论(1) 微生物对蒙脱石层间吸附的有机质的降解方式主要有分泌有机酸直接降解和破坏层间结构释放有机物从而进行降解。代表菌种假单胞菌和希瓦氏菌对半胱氨酸绿脱石及甲苯绿脱石的作用表明,微生物通过分泌有机酸的形式对蒙脱石层间吸附的有机质降解作用很有限,该结构在恒定的有氧和无氧条件下对保存有机质有利;
(2) 希瓦氏菌在严格无氧条件下通过还原Fe(Ⅲ)进行代谢,实验表明,无氧条件下,希瓦氏菌可以一定程度破坏矿物结构,释放并消耗有机物,因此,铁还原微生物对蒙脱石层间吸附有机质的保存有一定的影响,但由于微生物对矿物晶体结构的破坏能力有限,故其对层间吸附有机质降解的能力也有限;
(3) 不同有机物对生物降解过程也有影响,这些影响取决于有机质的特性及有机质与细菌之间的相互作用。绿脱石层间吸附的半胱氨酸对生物生长有利,从而可能促进生物还原Fe(Ⅲ)作用。相反,甲苯却很明显的抑制了Fe(Ⅲ)的还原。
| [] | Bader RG, Hood DW and Smith JB. 1970. Recovery of dissolved organic matter in seawater and organic sorption by particulate material. Geochimica et Cosmochimica Acta , 19 :236–243. |
| [] | Bergamashi BA, Tsamakis E, Keil RA, Eglington TI, Montlucon DB and Hedges JI. 1997. The effect of grain size and surface area on organic matter, lignin and carbohydrate concentration, and molecular compositions in Peru Margin sediments. Geochimica et Cosmochimica Acta , 61 :1247–1260. DOI:10.1016/S0016-7037(96)00394-8 |
| [] | Bishop AN, Kearsley AT and Patience RL. 1992. Analysis of sedimentary organic materials by scanning electron microscopy: the application of back scattered electron imagery and light element X-ray microanalysis. Organic Geochemistry , 18 :431–446. DOI:10.1016/0146-6380(92)90106-8 |
| [] | Bishop AN and Phillip RP. 1994. Potential for amorphous kerogen formation via adsorption of organic material at mineral surfaces. Energy and Fuels , 8 :1494–1497. DOI:10.1021/ef00048a040 |
| [] | Bock MJ and Mayer LA. 2000. Mesodensity organo-clay associations in a near shore sediment. Marine Geology , 163 :65–75. DOI:10.1016/S0025-3227(99)00105-X |
| [] | Collins MJ, Bishop AN and Farrimond P. 1995. Sorption by mineral surfaces: Rebirth of the classical condensation pathway for kerogen formation. Geochimica et Cosmochimica Acta , 59 :2387–2391. DOI:10.1016/0016-7037(95)00114-F |
| [] | Demaison GT and Moor GT. 1980. Anoxic environments and oil source bed genesis. AAPG Bulletin , 64 :1179–1180. |
| [] | Gordon AS and Millero FJ. 1985. Adsorption mediated decrease in the biodegradation rate of organic compounds. Microbial Ecology , 11 :289–298. DOI:10.1007/BF02016813 |
| [] | Hedges JI and Keil RG. 1995. Sedimentary organic matter preservation: An assessment and speculative synthesis. Marine Geology , 49 :81–115. |
| [] | Hedges JI and Oades JM. 1997. Comparative organic geochemistries of soils and marine sediments. Organic Geochemistry , 27 :319–361. DOI:10.1016/S0146-6380(97)00056-9 |
| [] | Hedges JI and Keil RG. 1999. Organic geochemical perspective on esturine processes: sorption reactions and consequences. Marine Chemistry , 65 :55–65. DOI:10.1016/S0304-4203(99)00010-9 |
| [] | Heinrichs SM and Sugai SF. 1993. Adsorption of amino acids and glucose by sediments of Resurrection Bay(Alaska): Functional group effects. Geochimica et Cosmochimica Acta , 57 :823–835. DOI:10.1016/0016-7037(93)90171-R |
| [] | Kajiwara Y, Yamakita S, Ishida K and Ishiga H. 1994. Development of a largely anoxic stratified ocean and its temporary massive mixing at the Permian/Triassic boundary supported by sulfur isotopic record. Palaeogeography, Palaeoclimatology, Palaeoecology , 111 :367–379. DOI:10.1016/0031-0182(94)90072-8 |
| [] | Kakuwa Y. 1996. Permian-Triassic mass extinction event recorded in bedded chert sequence in Southwest Japan. Palaeogeography, Palaeoclimatology, Palaeoecology , 121 :35–51. DOI:10.1016/0031-0182(95)00070-4 |
| [] | Keil RG and Hedges JI. 1993. Sorption of organic matter to mineral surfaces and the preservation of organic material in coastal marine sediments. Chemical Geology , 107 :385–388. DOI:10.1016/0009-2541(93)90215-5 |
| [] | Kennedy MJ, Pevear DR and Hill RH. 2002. Mineral surface control of organic carbon in black shale. Science , 295 :657–660. DOI:10.1126/science.1066611 |
| [] | Mayer LM. 1994a. Surface area control of organic carbon accumulation in continental shelf sediments. Geochimica et Cosmochimica Acta , 58 :1271–1284. DOI:10.1016/0016-7037(94)90381-6 |
| [] | Mayer LM. 1994b. Relationship between mineral surfaces and organic cabon concentration in soils and sediments. Chemical Geology , 114 :347–363. DOI:10.1016/0009-2541(94)90063-9 |
| [] | Mayer LM. 1999. Extent of coverage of mineral surfaces by organic matter in marine sediments. Geochimica et Cosmochimica Acta , 63 :207–215. DOI:10.1016/S0016-7037(99)00028-9 |
| [] | Ransom B, Bennett RH, Baerwald R and Shea K. 1997. TEM study of in situ organic matter on continental margins: Occurrence and the monolayer hypothesis. Marine Geology , 138 :1–9. DOI:10.1016/S0025-3227(97)00012-1 |
| [] | Ransom B, Dongseon K, Kastner M and Wainwright S. 1998. Organic matter preservation on continental slopes: Importance of mineralogy and surface area. Geochimica et Cosmachimica Acta , 62 :1329–1345. DOI:10.1016/S0016-7037(98)00050-7 |
| [] | Salmon V, Derenne S, Lallier-Verges E, Largeau C and Beaudoin B. 2000. Protection of organic matter by mineral matrix in a Cenomanian black shale. Organic Geochemistry , 31 :463–474. DOI:10.1016/S0146-6380(00)00013-9 |
| [] | Suzuki N, Ishida K, Shinomiya Y and Ishiga H. 1998. High productivity in the earliest Triassic ocean: Black shales, Southwest Japan. Palaeogeography, Palaeoclimatology, Palaeoecology , 141 :53–65. DOI:10.1016/S0031-0182(98)00009-1 |
| [] | Zhang GX, Kim JW, Dong HL and Sommer AJ. 2007. Microbial effects in promoting the smectite to illite reaction: Role of organic matter intercalated in the interlayer. American Mineralogist , 92 :1401–1410. DOI:10.2138/am.2007.2331 |
| [] | Zhao XY, Wang XX, Zhang YY, Xin GQ and Wang JK.1995. Clay Minerals in Chinese Petroliferous Basin. Wuhan: China University of Geosciences Press : 50 -114. |
| [] | Zhu GY, Zhao WZ, Zhang SC, Liang YB and Wang ZJ. 2007. Discussion of gas enrichment mechanism and natural gas origin in marine sedimentary basin, China. Chinese Science Bulletin , 52 (Supp1.) :62–76. |
| [] | 赵杏媛, 王行信, 张有瑜, 辛国强, 王经科. 1995. 中国含油气盆地黏土矿物. : 50 -114. |