 2023,
Vol. 32
2023,
Vol. 32



2. 自然资源部第二海洋研究所, 浙江 杭州 310012;
3. 上海海洋大学 环境DNA技术与水生态健康评估工程中心, 上海 201306
仔稚鱼时期是鱼类发育早期的浮游生活阶段。在此阶段海洋环境的任何细微变化,都可能对仔稚鱼的生长发育产生致命的影响[1]。许多研究[2-3]已探明仔稚鱼的群落组成、时空分布与海洋环境相关。
南海是西太平洋最大的半封闭边缘海,也是受东南亚季风影响的核心区域,其地理位置特殊,与吕宋海峡、民都洛海峡、台湾海峡、卡里马塔海峡等海域相连通[4-5]。夏季,在西南季风的驱使下,南海北部陆架海域形成季节性的沿岸上升流,主要包括粤东沿岸上升流和琼东沿岸上升流[6]。上升流能将含有丰富营养盐的低温深层水运送至表层,从而使沿岸上升流海区的表层水也具有深层水的低温、高盐等特征[7]。已有研究[8]表明,全球海洋近20%的渔业捕捞量,与仅占全球海洋表面积l‰的沿岸上升流密切相关。因此,开展沿岸上升流海区仔稚鱼资源的调查研究,有利于促进该海域鱼类资源的保护、合理开发与利用。除上升流之外,高营养盐的珠江口冲淡水也对南海北部水域的生态特征产生重要影响[9]。
目前对南海北部海域仔稚鱼的调查研究工作,已经取得一定进展。20世纪六十年代,南海水产研究所在南海北部调查中,对金线鱼(Nemipterus virgatus)等重要经济鱼种的仔稚鱼进行了相关研究[10];周美玉等[11]基于北部湾鱼类浮游生物的综合调查,共鉴定出153个类群;李开枝等[12]在南海北部有关鱼类浮游生物的调查中,报道了95种鱼卵、仔稚鱼,并揭示了该区域鱼类浮游生物多样性和丰度的季节变化;肖瑜璋等[13]研究了珠江口的鱼类浮游生物,共鉴定出109个类群;侯刚等[14]结合形态学鉴定技术和DNA条形码技术鉴定了南海北部的鱼类浮游生物,共鉴定出213个仔稚鱼类群;王九江等[15]在大亚湾鱼卵、仔稚鱼种群的调查中,研究了其特征与环境因子的相关性,共鉴定了鱼卵、仔稚鱼27个类群。但过往研究主要集中在南海近海的鱼类早期资源调查,结合珠江口冲淡水、上升流与仔稚鱼资源关系的大尺度调查研究相对较少,仅HUANG等[16]和CHEN等[17]结合相关水文特征对仔稚鱼资源进行了研究。本研究基于南海北部海域仔稚鱼调查,探讨种类组成与环境因子的关系,探究南海北部海域水文特征与仔稚鱼群聚的关系,旨在为南海北部鱼类早期资源的保护提供科学依据。
1 材料和方法 1.1 调查方法2012年夏季7月和9月在南海北部海域(110°E~120°E、16°N~22°N)设置23个采样站位(7月琼东海域10个站位、9月粤东海域13个站位),见图 1。按照中国海洋综合调查与评价专项的《海洋调查规范第6部分:海洋生物调查》(GB/T 12763.6—2007)规范,采用装有机械式流量计(HYDRO-BIOS)的WP2浮游生物网(网口内径57 cm,网长271 cm,网目200 μm)自水深200 m处到表层进行垂直拖网。样本采用体积分数为5%甲醛溶液保存。

|
图 1 南海北部调查站位图 Fig. 1 Sampling stations in the northern South China Sea |
基于仔稚鱼营浮游生活,主要分布于表层的特征,研究采用表层的相关环境数据。所使用的海表面温度(Sea surface temperature,SST)、海表面盐度(Sea surface salinities,SSS)和叶绿素a质量浓度(Chlorophyll a concentration,Chl.a)等环境参数均来自卫星遥感资料。海表面温度和叶绿素a质量浓度等数据来自美国国家航空航天局(National Aeronautics and Space Administration, NASA)官方网站(https://oceancolor.gsfc.nasa.gov/)。海表面盐度数据来源于法国瓦尔数据中心(Centre Aval de Traitement des Données SMOS, CATDS) 官方网站(http://www.salinityremotesensing.ifremer.fr/)。
1.3 样品分析在AXIO Zoom.V16体视显微镜下对仔稚鱼样品进行物种鉴定、个体计数及其体长测量工作,参考《中国近海鱼卵与仔鱼》和《日本产稚鱼图鉴》等进行物种鉴定。鱼类学名主要参照世界海洋生物物种登记库(Wo RMS, http://www.marinespecies.org),中文名主要参照《拉汉世界鱼类系统名典》, 鱼类分类系统参照《Fishes of the World》第五版和《中国海洋及河口鱼类系统检索》。对调查海域所采集的仔稚鱼按照生态类型进行划分,根据适温类型可划分为暖水性鱼类(Warm water species)、暖温性鱼类(Warm temperate species)[18];根据栖息水层可划分为中上层鱼类和底层、近底层鱼类[19]。
1.4 数据分析仔稚鱼的丰度(G)标准化为每100 m3的个体数量(ind./100 m3):
 (1)
(1)
式中:N为每次拖网所采集到的仔稚鱼个体数,ind.;S为网口面积,m2;L为流量计转数;C为流量计校正值,C= 0.3。
Shannon-Wiener多样性指数(H′)[20]、Pielou均匀度指数(J′)[21]和Margalef丰富度指数(D)[22]分别用于计算物种多样性和均匀度:
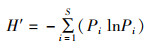 (2)
(2)
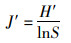 (3)
(3)
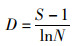 (4)
(4)
式中:S为一个样本中的物种总数;N为一个样本中的总尾数;Pi为i物种所占总尾数的比例。
应用典型相关分析(Canonical correspondence analysis, CCA)来分析仔稚鱼的丰度与海表面温度、海表面盐度和叶绿素a质量浓度的关系。以Ocean Data View4、Excel 2016、PRIMER 6.0以及Canoco 5.0完成数据分析及作图。
2 结果 2.1 种类组成本研究共采集到仔稚鱼230尾,隶属11目33科77种(表 1)。其中8种鉴定到科,23种鉴定到属,46种鉴定到种。鲈形目(Perciformes)种类最多,有16科24种,占调查海域仔稚鱼总种类数的31.17%。其次是灯笼鱼目(Myctophiformes)和巨口鱼目(Stomiiformes),分别有1科18种和4科12种。灯笼鱼目个体数最多,占总个体数的37.39%,其次是鲈形目(30.43%)和巨口鱼目(15.22%)。灯笼鱼科(Myctophidae)、巨口光灯鱼科(Phosichthyidae)、钻光鱼科(Gonostomatidae)的仔稚鱼数量约占仔稚鱼总捕获量的50%。琼东海域采集到仔稚鱼106尾,隶属10目24科43种。其中灯笼鱼目种类最多,有1科15种。其次是鲈形目和巨口鱼目,分别有10科13种和3科9种。粤东海域采集到仔稚鱼124尾,隶属6目19科47种。其中鲈形目种类最多,有11科15种。其次是灯笼鱼目和巨口鱼目,分别有1科8种和3科6种。

|
表 1 南海北部海域仔稚鱼的种类组成 Tab.1 Species composition of fish larvae and juveniles in the northern South China Sea |
南海北部仔稚鱼平均丰度为15.71 ind./100 m3。其中,粤东海域仔稚鱼隶属19科47种,平均丰度为13.13 ind./100 m3。琼东海域仔稚鱼隶属24科43种,平均丰度为19.08 ind./100 m3。仔稚鱼的水平分布显示出较大的站间差异(P < 0.05),其中B4站点丰度最高,达51.09 ind./100 m3,B7站丰度最低,为1.65 ind./100 m3(图 2)。

|
图 2 南海北部仔稚鱼丰度分布图 Fig. 2 Abundances of fish larvae and juveniles in the northern South China Sea |
聚类分析研究结果显示,采样站位在1.95%的水平上相似,根据地理位置和地形,可划分为2个群组: 大洋组和近海组(图 3)。大洋组包括站位A4、A5、A6、A7、A8、B5、B6、B7、B8、B9、B10、B11、B12、B14、B15和B16;近海组包括站位A1、A2、A3、A9、A10、B2和B4。大洋组多为栖息在水深较深或深海海域的鱼类,其中灯笼鱼目数量最多,占该组仔稚鱼总捕获量的50.00%,眶灯鱼属未定种(Diaphus sp.)是大洋组中数量最多的种类。近海组中鲈形目数量最多,占该组仔稚鱼总捕获量的56.06%,史氏谐鱼(Emmelichthys struhsakeri)是近海组中数量最多的种类。

|
图 3 南海北部海域仔稚鱼调查站位聚类分析图 Fig. 3 Group average clustering based on fish larvae and juveniles species among the sampled stations in the northern South China Sea |
各站间仔稚鱼种类数的变化,与Margalef丰富度指数的变化趋势较为一致。在粤东海域,B10站位仔稚鱼的Margalef丰富度指数和Shannon-Wiener多样性指数最高,B7站位Shannon-Wiener多样性指数和Pielou均匀度指数最低。在琼东海域,A8站位仔稚鱼的Margalef丰富度指数和Shannon-Wiener多样性指数最高,A10站位Shannon-Wiener多样性指数和Margalef丰富度指数最低(表 2)。大洋组Margalef丰富度指数、Shannon-Wiener多样性指数和Pielou均匀度指数的平均值均略高于近海组,而仔稚鱼平均丰度低于近海组(表 3)。各站位间仔稚鱼的Margalef丰富度指数和Shannon-Wiener多样性指数的变化较明显,而除B7站位外各站位间Pielou均匀度指数的变化相差并不大。总体而言,粤东海域和琼东海域的仔稚鱼群落结构相对稳定。

|
表 2 各站物种数量、仔稚鱼丰度和生物多样性指数 Tab.2 Number of species, abundance of fish larvae and juveniles, and biodiversity index at each station |

|
表 3 南海北部近海组与大洋组生物多样性指数及丰度平均值 Tab.3 Average of biodiversity index and abundance in the offshore group and oceanic group in the northern South China Sea |
由2012年夏季海表面温度(SST)的平面分布图可以看出,夏季南海北部陆架区海表面温度存在着明显的低温区,主要分布于海南岛东部沿岸一带海域(图 4a),粤东沿岸114°E~120°E区域存在有明显的低温带(图 4b),其海表面温度普遍低于外海2~3 ℃。夏季南海北部的大部分研究区域海水温度都较高,最高温度为30.1 ℃,同时最低温度为28.6 ℃。夏季沿岸上升流海区常显现出低温、高盐两大特征。而在本研究调查海域,琼东沿岸观察到异于周围海域的低温、高盐现象(图 4a、4c),表明调查期间琼东沿岸上升流的存在。此外,在西南季风的影响下,低盐度、高叶绿素a质量浓度的珠江冲淡水(图 4d、4f)沿大陆架向东扩散,加强了陆源水的跨海岸输送。

|
图 4 调查区域2012年7、9月海表面环境因子(温度、盐度和叶绿素a质量浓度)平面分布图 Fig. 4 The sea surface temperature(℃), sea surface salinities and chlorophyll a mass concentration (μg/L) at the depth of 5 m in July and September 2012 |
研究海域仔稚鱼的分布受海表面温度(SST)和叶绿素a质量浓度(Chl.a)影响较大(图 5)。CCA第一排序轴的特征值为0.500 7,CCA第二排序轴的特征值为0.297 8,各个物种与各个环境因子的相关性分析表明,CCA第一排序轴与环境因子的相关性数值为0.847 4,CCA第二排序轴与环境因子的相关性数值为0.866 5。蒙特卡洛检验的结果显示,第一排序轴和第二排序轴均未呈现显著性差异(F=1.9,P=0.182>0.05;F=1.2,P=0.476>0.05)。两个排序轴共解释了仔稚鱼种类组成23.22%的变异,其中,第一排序轴与叶绿素a质量浓度(Chl.a)及海表面盐度(SSS)呈较大正相关,而与海表面温度(SST)呈较大负相关。其中后灯笼鱼属未定种(Metelectrona sp.)、炬灯鱼属未定种(Lampadena sp.)、串光鱼属未定种(Vinciguerria sp.)主要受Chl.a、SST影响,环境因子与其分布呈正相关;圆罩鱼属未定种(Cyclothone sp.)主要受SST、Chl.a影响,与Chl.a呈正相关,与SST呈负相关;史氏谐鱼、真鲷(Pagrus major)、光滑平鲉(Sebastes levis)主要受SST影响,并与SST呈正相关。

|
图 5 南海北部仔稚鱼与环境因子的CCA排序 Fig. 5 CCA biplot of species composition of survey stations and environmental factors in the northern South China Sea |
南海是西太平洋最大的半封闭海,不仅地形复杂,而且与众多海域相连通,具有复杂的水文特征[23],南海北部水团的性质和分布在很大程度上受到吕宋海峡水体交换以及黑潮入侵南海的影响[24]。在夏季,南海西南季风的盛行,增强了黑潮主流的北上势力,削弱了黑潮分支对南海的入侵[25]。因此,南海北部近海海域主要受南海暖流影响[16];近岸海域受南海暖流和珠江冲淡水的影响[26-27];海南岛东部沿岸会受到季节性的沿岸上升流的影响[28]。
本研究中,灯笼鱼科、巨口光灯鱼科、钻光鱼科的仔稚鱼数量约占总捕获量的50%,该结果与以往的许多调查结果一致,这表明南海北部仔稚鱼的种类组成相对稳定[16-17, 29]。灯笼鱼科是一类在海洋中生物量很大的海洋鱼类,种类也十分的丰富(>230种),其仔稚鱼在其他海域的调查中也占有很高的比例[30]。
仔稚鱼的分布和物种组成还会受到水文条件、季节性季风驱动流和地理特征等因素的影响[31-32]。已有研究[33-34]表明鱼类早期资源的分布模式与水体的理化性质密切相关,在低温高盐的上升流海区,具有相对较高的仔稚鱼丰度。在本研究中,靠近琼东沿岸上升流海区的琼东海域A1、A2、A3站仔稚鱼丰度明显较高。靠近西沙群岛的A8、A9站出现较高的仔稚鱼丰度。可能是人类活动增加了水体营养丰富度所致[35]。高永利等[36]观察到在西沙群岛的各岛屿中,岛屿人口越多,水体中叶绿素a质量浓度和溶解无机氮含量越高,水体富营养化趋势越强。叶绿素a质量浓度通过影响浮游植物类群分布,从而也直接或间接地影响了仔稚鱼的分布[37]。肖瑜璋等[38]在珠江口水域鱼卵和仔稚鱼调查研究中发现,仔稚鱼个体数量和种类数与叶绿素a质量浓度含量均呈正相关。在具有高叶绿素a质量浓度的B4、B5站仔稚鱼丰度较高,则可能是受珠江口冲淡水影响。珠江冲淡水携带大量的营养盐,并且在夏季高温、强光照的作用下,浮游植物大量生长,浮游生物量也会增加,进一步丰富了该海域的鱼类饵料生物,为仔稚鱼的聚集栖息提供了良好的生存保障[39]。
3.2 粤东海域、琼东海域及相似海域仔稚鱼资源的比较粤东海域位于南海北部大陆架区,和琼东海域同处于热带季风影响区,从适温属性来看,两海域仔稚鱼具有明显的暖温带和亚热带特点,其中暖水性鱼类占绝对优势,其次是暖温性鱼类[40]。粤东海域和琼东海域地理位置相近,生境相似,并且都受到台湾暖流的影响[41]。本研究结果表明,两海域仔稚鱼共同种多达11种,其中暖水性鱼类10种,暖温性鱼类1种。从栖息水层来看,两海域均为中上层鱼类居多。从丰度分布水平来看,琼东海域的仔稚鱼丰度略高于粤东海域,这可能是由于采样时正处于琼东沿岸上升流作用的强盛期。两海域物种多样性指数无显著性差异。总体而言,两海域水体交换频繁,联系密切,仔稚鱼资源均较为丰富。受地理位置及洋流影响,两海域仔稚鱼资源也具有一定的相似性。
以往珠江口仔稚鱼的调查研究中,捕获的仔稚鱼多为近岸种类和近海种类,以康氏小公鱼(Stolephorus commersonii)、虾虎鱼科未定种(Gobiidae sp.)等为代表种类[13, 36]。本研究中粤东海域B断面跨越的空间尺度更大,因此在种类组成中大洋性鱼类更多,以眶灯鱼属未定种、大西洋明灯鱼(Diogenichthys atlanticus)等为代表种类。水深是影响鱼类群落结构变化的主要因素[42],粤东海域B断面由西向东,随水深梯度的变化,各站位仔稚鱼呈现出由近海种类向大洋性种类变化的趋势。SU等[34]的研究发现,台湾东部仔鱼群落中大洋性鱼类居多,其中鲭科(Scombridae)和灯笼鱼科数量较多。这与粤东海域靠近台湾南部的B14、B15、B16站捕获的仔稚鱼种类具有一定的相似性。吴钟解等[43]研究认为海南岛东南部珊瑚礁鱼类种类丰富,并且珊瑚礁鱼类的密度由南向北呈现逐渐递减的趋势。在本研究中琼东海域近海组站位A1、A2、A3、A9、A10站,聚集着丰富的珊瑚礁鱼类,包括鲬鱼(Platycephalus indicus)、斑点鸡笼鲳(Drepane punctata)、眼镜鱼(Mene maculata)等。
南海北部气候适宜,渔业资源丰富,然而随着经济的发展,由于过度捕捞及环境污染等人类活动,南海北部鱼类资源已逐渐衰退[44]。由各站Shannon-Wiener多样性指数可以看出,人类活动已在一定程度上影响了粤东海域和琼东海域的生态环境[45]。因此,须进一步提升渔政执法能力,加强对该海域的生态环境的监测和保护,保证南海渔业资源的可持续发展。
| [1] |
万瑞景, 孙珊. 黄、东海生态系统中鱼卵、仔稚幼鱼种类组成与数量分布[J]. 动物学报, 2006, 52(1): 28-45. WAN R J, SUN S. The category composition and abundance of ichthyoplankton in the ecosystem of the Yellow Sea and the East China Sea[J]. Current Zoology, 2006, 52(1): 28-45. |
| [2] |
邱永松. 南海北部大陆架鱼类群落的区域性变化[J]. 水产学报, 1988, 12(4): 303-313. QIU Y S. The regional changes of fish community on the northern continental shelf of South China[J]. Journal of Fisheries of China, 1988, 12(4): 303-313. |
| [3] |
LI Z G, YE Z J, WAN R. Spatial and seasonal patterns of ichthyoplankton assemblages in the Haizhou Bay and its adjacent waters of China[J]. Journal of Ocean University of China, 2015, 14(6): 1041-1052. DOI:10.1007/s11802-015-2603-3 |
| [4] |
许婷, 曹永港. 南海海洋环流季节变化的数值模拟研究[J]. 海岸工程, 2017, 36(1): 62-71. XU T, CAO Y G. Numerical simulation study on the seasonal variations of ocean circulations in the South China Sea[J]. Coastal Engineering, 2017, 36(1): 62-71. DOI:10.3969/j.issn.1002-3682.2017.01.007 |
| [5] |
苏纪兰. 南海环流动力机制研究综述[J]. 海洋学报(中文版), 2005, 27(6): 1-8. SU J L. Overview of the South China Sea circulation and its dynamic[J]. Acta Oceanologica Sinica, 2005, 27(6): 1-8. DOI:10.3321/j.issn:0253-4193.2005.06.001 |
| [6] |
汪彧, 经志友, 齐义泉. 2013年夏季琼东海域上升流观测研究[J]. 热带海洋学报, 2016, 35(2): 40-49. WANG Y, JING Z Y, QI Y Q. Coastal upwelling off eastern Hainan Island observed in the summer of 2013[J]. Journal of Tropical Oceanography, 2016, 35(2): 40-49. |
| [7] |
谢玲玲, 张书文, 赵辉. 琼东上升流研究概述[J]. 热带海洋学报, 2012, 31(4): 35-41. XIE L L, ZHANG S W, ZHAO H. Overview of studies on Qiongdong upwelling[J]. Journal of Tropical Oceanography, 2012, 31(4): 35-41. |
| [8] |
PAULY D, CHRISTENSEN V. Primary production required to sustain global fisheries[J]. Nature, 1995, 374: 255-257. DOI:10.1038/374255a0 |
| [9] |
杨阳, 李锐祥, 朱鹏利, 等. 珠江冲淡水季节变化及动力成因[J]. 海洋通报, 2014, 33(1): 36-44. YANG Y, LI R Y, ZHU P L, et al. Seasonal variation of the Pearl River diluted water and its dynamical cause[J]. Marine Science Bulletin, 2014, 33(1): 36-44. |
| [10] |
中华人民共和国水产部南海水产研究所. 南海北部底拖网鱼类资源调查报告(海南岛以东)第一册[M]. 中华人民共和国水产部南海水产研究所, 1966: 9-29. South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences. Survey report on bottom trawl fish resources in northern South China Sea (east of Hainan Island) Volume 1[M]. South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, 1966: 9-29. |
| [11] |
周美玉, 林元烧, 杨圣云, 等. 北部湾鱼类浮游生物种类组成和数量分布[J]. 海洋学报(英文版), 2011, 30(1): 94-105. ZHOU M Y, LIN Y S, YANG S Y, et al. Composition and ecological distribution of ichthyoplankton in eastern Beibu Gulf[J]. Acta Oceanologica Sinica, 2011, 30(1): 94-105. |
| [12] |
李开枝, 尹建强, 黄良民, 等. 南海北部上层鱼类浮游生物多样性和丰度的季节变化[J]. 海洋学报(英文版), 2014, 33(12): 145-154. LI K Z, YIN J Q, HUANG L M, et al. Seasonal variations in diversity and abundance of surface ichthyoplankton in the northern South China Sea[J]. Acta Oceanologica Sinica, 2014, 33(12): 145-154. |
| [13] |
肖瑜璋, 王蓉, 郑琰晶, 等. 珠江口鱼类浮游生物种类组成与数量分布[J]. 热带海洋学报, 2013, 32(6): 80-87. XIAO Y Z, WANG R, ZHENG Y J, et al. Species composition and abundance distribution of ichthyoplankton in the Pearl River Estuary[J]. Journal of Tropical Oceanography, 2013, 32(6): 80-87. DOI:10.3969/j.issn.1009-5470.2013.06.012 |
| [14] |
侯刚. 南海习见鱼类DNA条形码数据库与仔稚鱼种类鉴定研究[D]. 北京: 中国科学院大学, 2017. HOU G. Studies on DNA barcoding and larvae identification of fishes in South China Sea[D]. Beijing: University of Chinese Academy of Sciences, 2017. |
| [15] |
王九江, 刘永, 肖雅元, 等. 大亚湾鱼卵、仔稚鱼种群特征与环境因子的相关关系[J]. 中国水产科学, 2019, 26(1): 14-25. WANG J J, LIU Y, XIAO Y Y, et al. Population characteristics of fish eggs and larvae and their relation-ship with environmental factors in Daya Bay[J]. Journal of Fishery Sciences of China, 2019, 26(1): 14-25. |
| [16] |
HUANG D L, ZHANG X, JIANG Z J, et al. Seasonal fluctuations of ichthyoplankton assemblage in the northeastern South China Sea influenced by the Kuroshio intrusion[J]. Journal of Geophysical Research: Oceans, 2017, 122(9): 7253-7266. DOI:10.1002/2017JC012906 |
| [17] |
CHEN L C, LAN K W, CHANG Y, et al. Summer Assemblages and Biodiversity of Larval Fish Associated with Hydrography in the Northern South China Sea[J]. Marine and Coastal Fisheries, 2018, 10(5): 467-480. DOI:10.1002/mcf2.10037 |
| [18] |
田明诚, 孙宝龄, 杨纪明. 渤海鱼类区系分析[J]. 海洋科学集刊, 1993(34): 157-167. TIAN M C, SUN B L, YANG J M. Analysis of the fish fauna of the BoHai sea[J]. Studia Marina Sinica, 1993(34): 157-167. |
| [19] |
王小林. 海州湾及邻近海域鱼类群落结构的时空变化[D]. 青岛: 中国海洋大学, 2013: 9-15. WANG X L. Temporal and spatial variations of the fish community structure in Haizhou Bay and adjacent waters[D]. Qingdao: China Ocean University, 2013: 9-15. |
| [20] |
WILHM J L. Use of biomass units in Shannon's formula[J]. Ecology, 1968, 49(1): 153-156. DOI:10.2307/1933573 |
| [21] |
PIELOU E C. The Measurement of Diversity in Different Types of Biological Collections[J]. Journal of Theoretical Biology, 1966, 13(1): 131-144. |
| [22] |
MARGALEF R. Composición específica del fitoplancton de la costa catalano-levantina (Mediterráneo occidental) en 1962-1967[J]. Investigación Pesquera, 1969, 33(1): 345-380. |
| [23] |
程国胜, 孙佳东, 俎婷婷, 等. 2011年夏季南海北部海区水团分析[J]. 热带海洋学报, 2014, 33(3): 10-16. CHENG G S, SUN J D, ZU T T, et al. Analysis of water masses in the northern South China Sea in summer 2011[J]. Journal of Tropical Oceanography, 2014, 33(3): 10-16. |
| [24] |
NAN F, XUE H J, YU F. Kuroshio intrusion into the South China Sea: A review[J]. Progress in Oceanography, 2015, 137(2015): 314-333. |
| [25] |
CHEN Y X, LIN S Q, WANG C S, et al. Response of size and trophic structure of zooplankton community to marine environmental conditions in the northern South China Sea in winter[J]. Journal of Plankton Research, 2020, 42(3): 378-393. |
| [26] |
WANG X Z, WEI Y Q, WU C, et al. The profound influence of Kuroshio intrusion on microphytoplankton community in the northeastern South China Sea[J]. Acta Oceanologica Sinica, 2020, 39(8): 79-87. |
| [27] |
YIN K D, QIAN P Y, WU M C S, et al. Shift from P to N limitation of phytoplankton growth across the Pearl River estuarine plume during summer[J]. Marine Ecology Progress Series, 2001, 221(4): 17-28. |
| [28] |
许金电, 蔡尚湛, 宣莉莉, 等. 2006年夏季琼东、粤西沿岸上升流研究[J]. 海洋学报(中文版), 2013, 35(4): 11-18. XU J D, CAI S Z, XUAN L L, et al. Study on coastal upwelling in eastern Hainan Island and western Guangdong in summer 2006[J]. Acta Oceanologica Sinica, 2013, 35(4): 11-18. |
| [29] |
袁梦, 陈作志, 张俊, 等. 南海北部陆坡海域中层渔业生物群落结构特征[J]. 南方水产科学, 2018, 14(1): 85-91. YUAN M, CHEN Z Z, ZHANG J, et al. Community structure of mesopelagic fish species in northern slope of South China Sea[J]. South China Fisheries Science, 2018, 14(1): 85-91. |
| [30] |
HSIEH H Y, Lo W T, LIU D C, et al. Influence of hydrographic features on larval fish distribution during the south-westerly monsoon in the waters of Taiwan, western North Pacific Ocean[J]. Journal of Fish Biology, 2010, 76(10): 2521-2539. |
| [31] |
SABATES A, OLIVAR M P, SALAT, et al. Physical and biological processes controlling the distribution of fish larvae in the NW Mediterranean[J]. Progress in Oceanography, 2007, 74(2-3): 355-376. |
| [32] |
CHEN W Y, LEE M A, LAN K W, et al. Distributions and assemblages of larval fish in the East China Sea during the northeasterly and southwesterly monsoon seasons of 2008[J]. Biogeosciences, 2014, 11(3): 547-561. |
| [33] |
LO W T, HSIEH H Y, WU L J, et al. Comparison of larval fish assemblages between during and after northeasterly monsoon in the waters around Taiwan, western North Pacific[J]. Journal of Plankton Research, 2010, 32(7): 1079-1095. |
| [34] |
SU W C, LO W T, LIU D C, et al. Larval fish assemblages in the Kuroshio waters east of Taiwan during two distinct monsoon seasons[J]. Bulletin of Marine Science, 2011, 87(1): 13-29. |
| [35] |
袁梁英. 南海北部营养盐结构特征[D]. 厦门: 厦门大学, 2005. YUAN L Y. Distribution and Characteristics of Nutrients in the Northern South China Sea[D]. Xiamen: Xiamen University, 2005. |
| [36] |
高永利, 黄晖, 练健生, 等. 西沙群岛礁栖鱼类物种多样性及其食性特征[J]. 生物多样性, 2014, 22(5): 618-623. GAO Y L, HUANG H, LIAN J S, et al. The species diversity and trophic structure of reef fishes in the waters of the Xisha Archipelago[J]. Biodiversity Science, 2014, 22(5): 618-623. |
| [37] |
孙强, 洪娅岚, 鹿凤. 珠江口邻域海洋初级生产力与人类活动相关性研究[J]. 科技创新与生产力, 2019(3): 12-15. SUN Q, HONG Y L, LU F, et al. A Study on the Correlation Between the Marine Primary Productivity of Pearl River Estuary and Human Activities[J]. Technology innovation and productivity, 2019(3): 12-15. |
| [38] |
肖瑜璋, 王蓉, 欧强, 等. 春季珠江口水域鱼卵和仔稚鱼的分布及其与环境因子的关系[J]. 台湾海峡, 2010(4): 488-495. XIAO Y Z, WANG R, OU Q, et al. Relationship between abundance distribution of fish eggs, larvae and juveniles and environmental factors in the Pearl River Estuary waters in spring[J]. Journal of oceanography in Taiwan Strait, 2010(4): 488-495. |
| [39] |
施玉珍, 赵辉, 王喜达, 等. 珠江口海域营养盐和叶绿素a的时空分布特征[J]. 广东海洋大学学报, 2019, 39(1): 56-65. SHI Y Z, ZHAO H, WANG X D, et al. Distribution Characteristics of Nutritive Salts and Chlorophyll a in the Pearl River Estuary[J]. Journal of Guangdong Ocean University, 2019, 39(1): 56-65. |
| [40] |
孙典荣, 李渊, 林昭进, 等. 海南岛近岸海域鱼类群落结构研究[J]. 中国海洋大学学报(自然科学版), 2011, 41(4): 33-38. SUN D R, LI Y, LIN Z J, et al. Community structure of fish in the coastal waters of Hainan Island[J]. Periodical of Ocean University of China, 2011, 41(4): 33-38. |
| [41] |
孙典荣, 李渊, 王雪辉. 海南岛近岸海域鱼类物种组成和多样性的季节变动[J]. 南方水产科学, 2012, 8(1): 1-7. SUN D R, LI Y, WANG X H. Seasonal changes of species composition and diversity of fishes in coastal waters of Hainan Island, China[J]. South China Fisheries Science, 2012, 8(1): 1-7. |
| [42] |
王雪辉, 林昭进, 杜飞雁, 等. 南海西北部陆架区鱼类的种类组成与群落格局[J]. 生态学报, 2013, 33(7): 2225-2235. WANG X H, LIN Z J, DU F Y, et al. Fish species composition and community pattern in the continental shelf of northwestern South China Sea[J]. Acta Ecologica Sinica, 2013, 33(7): 2225. |
| [43] |
吴钟解, 蔡泽富, 陈石泉, 等. 海南岛东岸、南岸浅水礁区近10年珊瑚礁鱼类种类分布及多样性探讨[J]. 水产学报, 2015, 39(8): 1203-1217. WU Z J, CAI Z F, CHEN S Q, et al. Species distribution and diversity of the coral reef fishes in the shallow reefs along the east and south coasts of Hainan Island[J]. Journal of Fisheries of China, 2015, 39(8): 1203-1217. |
| [44] |
李纯厚, 贾晓平, 杜飞雁, 等. 南海北部生物多样性保护现状与研究进展[J]. 海洋水产研究, 2005, 26(3): 73-79. LI C H, JIA X P, DU F Y, et al. Current status and advances for the biodiversity protection in the Northern area of the South China Sea[J]. Marine Fisheries Research, 2005, 26(3): 73-79. |
| [45] |
樊紫薇. 浙江中部近岸海域鱼卵、仔稚鱼群落结构特征研究[D]. 舟山: 浙江海洋大学, 2020: 53-54. FAN Z W. A study on the structural characteristics of ichthyoplankton in the middle of Zhejiang Province coastal waters[D]. Zhoushan: Zhejiang Ocean University, 2020: 53-54. |
2. The Second Institute of Oceanography, Ministry of Natural Resources, Hangzhou 310012, Zhejiang China;
3. Engineering Research Center of Environmental DNA and Ecological Water Health Assessment, Shanghai Ocean University, Shanghai 201306, China