 2022,
Vol. 31
2022,
Vol. 31



2. 大洋渔业资源可持续开发教育部重点实验室, 上海 201306;
3. 国家远洋渔业工程技术研究中心, 上海 201306;
4. 上海市水生野生动植物保护研究中心, 上海 200092;
5. 长江口水生生物资源监测与保护联合实验室, 上海 202162;
6. 中国水产科学研究院东海水产研究所, 上海 200090
我国对近海三疣梭子蟹(Portunus trituberculatus)资源的大规模开发始于20世纪80年代[1-2], 经过近40年的发展, 以梭子蟹为目标种类的渔业年产量近50万t[3]。蟹笼渔具以其结构简单、操作方便、渔获质量高等优点, 迅速成为东、黄海近海主要的捕捞作业渔具之一[4-5]。随着渔场范围不断扩大、捕捞努力量不断提高, 以及渔船甲板装备机械化和自动化程度逐渐提升, 单船携带蟹笼数量持续增加(目前已达8000笼), 蟹笼渔业给梭子蟹资源造成了巨大的压力, 单位捕捞努力量渔获量(CPUE)逐渐降低[4]、渔获个体逐渐减小[6], 三疣梭子蟹的最小可捕尺寸(MLS)从2004年《渤海生物资源养护规定》规定甲长不小于80mm锐减至2017年的49mm[7]。蟹笼数量的增加也加剧了渔民对渔场的争夺, 生产中因与其他渔具纠缠易导致蟹笼丢失, 其丢失后所产生的幽灵捕捞问题已受到了广泛的关注[8-12]。
甲壳类笼壶渔业实践已证实, 合理配置刚性释放装置不仅能减少非目标种类渔获[13-15]、改善渔具对目标种类的尺寸选择性[16-17], 还能减轻入笼个体损伤或抛弃死亡[14-15, 17-18], 减轻丢失笼壶的幽灵捕捞问题[11, 13, 15, 18]。目前, 已在全球许多重要甲壳类笼壶渔业中强制使用释放装置[19-20]。在我国东海蟹笼渔业中, 吴常文[21]、张洪亮等[22]、白伦等[23]、ZHANG等[24]先后开展了刚性释放装置对幼蟹释放效果的研究。然而我国近海蟹笼渔具尺寸较小, 单船作业笼壶数量巨大, 探索并寻求适合于我国近海蟹笼渔业的释放装置与方法是维持蟹笼渔业可持续发展的迫切需要。
BROADHURST等[15]发现, 笼壶渔具网衣破损后可以有效释放入笼个体, 表明入笼的生物可以穿越网衣破目逃逸, 而生物可降解网衣(或网线)被认为是解决笼壶渔业幽灵捕捞问题的主要技术手段[15, 25]。相比刚性逃逸装置, 在蟹笼网衣上形成特定的网目结构从技术上更容易实现。对于无论是特定结构的网目还是网衣(或网线)降解后形成的破目, 笼内捕捞对象对其反应行为是决定个体能否逃逸的关键, 然而国内外鲜有研究提及甲壳动物对网目破目的逃逸行为。
鉴于此, 本研究旨在通过水池观察实验, 对比三疣梭子蟹对刚性矩形逃逸口和网目逃逸口的行为反应, 探究适合我国蟹笼渔具逃逸的选择性装置结构, 以期为今后开展蟹笼渔获规格选择和蟹笼渔具的管理提供参考。
1 材料与方法 1.1 样本来源本实验于2019年11月和2020年9月在江苏省通州湾南通经济产业园三疣梭子蟹养殖基地内完成。使用实验水箱(长2.11m×宽1.21m×高0.8m)开展笼内个体逃逸行为的观察。
实验样本采自池塘养殖的成体三疣梭子蟹。所选取样本均为体型完整、活跃度较高、不处于临近蜕壳或蜕壳后生殖阶段的个体。个体被取样后放入暂养水池, 保持与池塘养殖一致的投饵频率和种类[寻氏肌蛤(Musculus senhousei)或杂鱼]暂养2~3d, 待个体摄食、移动等行为稳定后备用于水池行为观察。实验中所采用的三疣梭子蟹甲宽为(123.4±41.3) mm, 甲高为(30.2±12.2) mm。
1.2 蟹笼及释放口设置采用我国东海近海渔业中普遍使用的圆柱形蟹笼作为实验笼。实验笼框架底部直径580mm, 笼高240mm, 外罩网目尺寸40mm的聚乙烯(PE)网衣; 蟹笼侧面分隔为6个部位, 其中3个间隔部位装配水平缩结0.707的网片, 另3个间隔部位设置3个外大内小、末端呈长扁椭圆形的网片式入口(图 1a), 实际作业中在笼壶的中央位置设置诱饵框。

|
图 1 实验蟹笼、逃逸口及摄像装备设置 Fig. 1 Configuration of experimental crab pots, escape openings, and mounting of the underwater observation devices |
依据实验所使用的三疣梭子蟹个体尺寸范围, 结合各体型参数之间的关系[6], 在白色聚丙烯(PP)平板(外尺寸长300mm×高100mm×厚3mm)中间切割出长200mm×宽40mm的矩形开口(图 1b)作为刚性矩形逃逸口, 开口边缘距平板边缘25mm, 并将平板边缘固定于蟹笼侧面靠近底部框架的网衣外侧(图 1a)。网目逃逸口使用破目方式实现, 即在侧网靠近蟹笼底部的位置(图 1c)裁剪出相邻的2个2a×3a(40mm×60mm)和1个3a×3a(60mm×60mm)的网目, 模拟网片中经线连续断裂3和4个目脚时形成的破目形式, 3a×3a网目设置于2a×3a网目上方2个网目的位置, 如图 1d所示。
1.3 行为观察及数据采集每次行为观察实验均从暂养群体中随机选取4~8尾三疣梭子蟹放入实验笼内, 观察并记录个体从放入开始直至当日实验结束范围(当日15:00到次日12:00)内个体的所有行为; 为了强化笼内个体的逃逸行为, 观察期间未设置诱饵框; 为了模拟野外水深较大时的行为, 实验全程在黑暗的环境下开展。
使用云台式旋转防水红外水下摄像头(品牌: 晟优, 型号: FS-412)进行行为观察和记录, 在刚性释放口的观察实验中将摄像头设置于笼壶的中央位置, 在网目逃逸口的观察实验中将摄像头设置于笼外约250mm处, 分别如图 1a和图 1c所示; 对观察到的视频内容进行录像并存储, 再通过对视频剪辑分析, 实现个体行为的鉴定及量化。
1.4 数据分析 1.4.1 行为分析及指标量化甲壳动物动作灵活、行为多样, 对笼壶渔具的行为反应较鱼类等更加复杂, 将个体对笼壶的连续行为离散成不同阶段, 并将个体从入笼至逃逸连续行为过程划分如下几个阶段, 实现特征行为分析和量化。
探索: 个体被放入笼内开始, 有目的性地搜索食物或逃逸出口; 发现并接近: 在逃逸口附近探索行为停止且明显停留一段时间; 穿越企图: 个体有针对性地姿态调整, 将部分步足、游泳足、螯足或部分头胸甲伸入逃逸口尝试穿越; 逃逸: 头胸甲全部穿越逃逸口, 完全脱离逃逸口并不再返回。
鉴于实验条件限制和研究目的, 仅对个体不同行为阶段的起始时刻、持续时间、特定行为次数以及成功与否等指标进行量化, 并记录个体在笼内的运动方向、个体间的相互行为、接近和穿越逃逸口的方向和姿态等状态。
1.4.2 行为指标的比较与统计假设三疣梭子蟹在不同阶段的行为相互独立, 使用方差分析(ANOVA)分析比较个体在不同行为阶段中特征行为的持续时间(如发现和接近逃逸口距离入笼的时间、逃逸口前停留时间以及个体穿越逃逸口的时间等)在不同类型逃逸口之间的差异, 使用广义线性混合模型(GLMM, Logit分布)分析个体企图逃逸等行为与相关影响因子的关系, 即
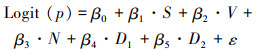 (1)
(1)
式中: p为个体是否企图逃逸(0或1);S为个体甲宽,mm; V为逃逸口类型; N为笼内个体数量, ind.; D1为距离入笼时间, h; D2为个体在逃逸口附近停留时间, s; β0为模型截距; β1~β5分别为各固定效应系数; ε为模型误差。除上述固定效应以外, 将月份和每次观察开始的时间作为随机效应。采用R语言lme4程序包中glmer函数实现GLMM模型分析。
2 结果与分析刚性逃逸口蟹笼实验共使用三疣梭子蟹43尾, 进行了8个观察日的水下观察; 网目逃逸口蟹笼实验共使用三疣梭子蟹24尾, 进行5个观察日的水下观察; 累计观察到个体接近逃逸口110次, 企图穿越逃逸口93次, 成功穿越逃逸口59次。
2.1 探索阶段三疣梭子蟹放入笼, 个体首先会保持静止一段时间(28±16) min以适应环境的变化, 而后表现出间歇性的探索行为; 探索行为主要发生在下午至凌晨, 但水下观察发现, 三疣梭子蟹在安装刚性逃逸口蟹笼中的探索时刻(主要集中在傍晚至凌晨3点, 图 2a)总体上早于网目逃逸口的实验蟹笼(傍晚18点至早晨6点); 从个体探索行为的发生距离入笼的时间(浸泡时间)来看, 刚性逃逸口蟹笼中主要集中在0~5h(图 2b), 而网目逃逸口蟹笼在各时间段内均有分布, 平均距离入笼时间明显大于刚性逃逸口蟹笼(图 2b, P=0.008)。

|
图 2 三疣梭子蟹在蟹笼中探索行为 Fig. 2 Search behavior for Portunus trituberculatus in crab pot |
当个体适应笼内环境后开始笼内探索行为, 即个体在蟹笼底部平面上背靠侧网做周向爬行或间歇性短暂游泳, 绕行一周的持续时间为2min左右, 持续时间不等; 在这一过程中, 当进行探索行为的个体遇见其他活跃个体时会表现出较为强烈的竞争行为, 即高举双螯、对峙驱赶, 当遇见静止个体时竞争行为相对缓和, 部分个体甚至尝试通过游泳或攀爬越过静止个体继续探索行为; 在没有安装逃逸口的对照蟹笼中, 在一段时间底部探索行为无果后, 大部分个体都会沿侧网网壁攀爬、部分个体甚至会停留在入口网片上方以探索蟹笼上部空间(图版-1)。

|
1.沿侧网攀爬至入口网片上方; 2.发现并接近矩形逃逸口; 3.发现并接近破目逃逸口; 4.逃逸口前的竞争行为; 5.尝试穿越刚性逃逸口; 6.尝试穿越破目逃逸口; 7.尝试穿越较高的破目逃逸口; 8.完全穿越刚性逃逸口; 9.个体穿越破目逃逸口时相邻网目变化。 1.Climbed along side panel to the upper panel of entrance; 2.Detected and approached the rigid rectangular escape vent; 3.Detected and approached the mesh escape opening; 4.Aggressive behavior in front of the rigid escape vent; 5.Attempted to pass through the rigid escape vent; 6.Attempt to pass through the lower broken mesh opening; 7.Attempted to pass through the higher broken mesh opening; 8.Completely passed through the rigid escape vent; 9.Distortion of the meshes near the broken meshes when crabs wedged by the opening. 图版 三疣梭子蟹在不同行为阶段中对不同类型逃逸口的行为 Plate Response behavior of P. trituberculatus to different escape openings in different stages of escaping behavior |
水下观察发现, 当笼内三疣梭子蟹通过探索行为定位并发现逃逸口后个体停止探索行为, 并在逃逸口附近停留(图版-2, 3)。在刚性逃逸口蟹笼中, 共观察到个体发现并接近逃逸口61次, 个体首次和所有发现并接近逃逸口距离入笼的平均时间分别为(0.91±0.79)h和(2.30±2.07)h, 个体平均发现并接近逃逸口次数为1.42次(=61/43, 图 3), 考虑到笼内数量随着个体逃逸逐渐减少, 并假设个体发现逃逸口行为相互独立, 则个体平均发现并接近逃逸口的次数(将一个观察日内个体每次发现逃逸口的概率累加获得)为(3.09±1.29)次; 而在网目逃逸口蟹笼中, 共观察到个体发现并接近逃逸口49次, 个体首次和所有发现并接近逃逸口时距离入笼的平均时间分别为(0.87±0.77)h(与刚性逃逸口无显著差异, P=0.940)和(6.13±4.91)h(显著大于刚性逃逸口, P < 0.001);个体平均发现逃逸口次数为2.04次(=49/24, 图版-2), 考虑到笼内数量随着个体逃逸逐渐减少后, 平均发现并接近逃逸口次数为(5.05±1.15)次, 与刚性逃逸口之间存在边际显著差异(P=0.061)。当多尾三疣梭子蟹同时出现发型并接近逃逸口时, 会表现出明显的竞争行为(图版-4)。

|
图 3 三疣梭子蟹对不同逃逸口的逃逸行为过程及量化指标 Fig. 3 Procedure and quantitative indices in different phases of escaping behavior for P. trituberculatus |
三疣梭子蟹发现并接近逃逸口后, 并不会立即穿越逃逸, 而是稍作停留后再尝试逃逸, 在刚性逃逸口附近的平均停留时间为(28.3±33.0)s, 而在柔性逃逸口附近平均停留时间为(116.0±207.3)s, 两者之间存在显著差异(P=0.006)。视频分析中共观察到93次逃逸企图, 其中刚性逃逸口51次, 即发现并接近刚性逃逸口的个体中83.7%选择企图逃逸, 16.3%选择远离逃逸口, 企图逃逸个体平均尝试逃逸次数为(1.18±0.53)次(图 3a); 而网目逃逸口蟹笼中观察到逃逸企图42次, 即发现并接近网目逃逸口的个体中85.6%选择企图逃逸, 14.4%的选择远离逃逸口, 企图逃逸的个体平均尝试逃逸次数为(1.63±0.76)次(图 3b), 显著大于刚性逃逸口个体平均尝试逃逸次数(P=0.001)。
使用GLMM模型分析逃逸口类型、笼内个体数量、个体大小、距离入笼的时间、个体在逃逸口附近停留的时间等因素对个体发现并接近逃逸口后是否企图逃逸这一行为的影响。结果(图 4)显示,个体在逃逸口附近的停留时间与个体是否企图逃逸之间存在显著的关系(P=0.049, 负相关), 即逃逸口附近停留时间越长, 企图逃逸的概率就越小, 而其他因素均对靠近逃逸口的个体是否选择逃逸企图没有显著影响。

|
图 4 三疣梭子蟹企图逃逸行为影响因素的GLMM拟合结果 Fig. 4 Output of GLMM fit for factors affecting attempt of P. trituberculatus to pass through escape opening |
所有个体均采用侧身的方式穿越逃逸口(图版-5, 6), 侧身的方向(左或右)取决于个体靠近逃逸口的方向(顺时针和逆时针), 且无明显差异。对于刚性逃逸口, 大多数个体会在逃逸口前适当调整姿态和方向, 使头胸甲一侧面向逃逸口, 随后将这侧的游泳足、步足以及头胸甲的一端先后伸入逃逸口, 两螯舒展并拢交替进行, 依靠步足爬动、游泳足划水推动, 使头胸甲全部穿越逃逸口。此次实验所选用的矩形逃逸口宽度较大(200mm), 因此逃逸口的高度是决定个体能否成功逃逸的关键因素, 当甲高较大个体会卡在逃逸口(图版-7)时, 大多会沿逃逸口横向移动, 同时借助两螯夹住逃逸口边缘继续调整姿态, 并伴随身体左右晃动或间歇性的剧烈抖动, 企图使头胸甲穿越逃逸口; 若尝试失败, 则会调转头胸甲方向, 用另一侧重复上述过程; 逃逸失败后, 部分个体退缩到逃逸口附近停留片刻后继续尝试穿越, 部分则停止逃逸行为并退缩至蟹笼其他区域。当逃逸口高度大于个体甲高时个体即可轻松穿越逃逸口, 但部分个体即便能顺利穿越, 仍会主动退缩至笼内停留片刻, 而后继续尝试穿越, 如此重复多次最终穿越逃逸, 单次成功穿越过程持续时间为(4.69±4.65)s。单次企图穿越刚性逃逸口的行为中68.7%(=35/51)逃逸成功, 总逃逸率为83.3%(=35/42)。
对于网目逃逸口, 企图逃逸的个体会不断调整姿态和位置以适应逃逸口的形状, 绝大多数个体首先选择了位置更低的逃逸口(2a×3a)尝试穿越, 此时个体将头胸甲一侧面向逃逸口, 游泳足、步足率先探入逃逸口(图版-6), 而后收拢双螯, 使头胸甲穿越逃逸口; 当个体尺寸较大无法顺利穿越较低的逃逸口时(图版-8), 部分个体会重新定位和搜索逃逸口, 当发现位置较高、尺寸更大的逃逸口后, 个体会背对或侧对网片, 步足依次轻触网目, 沿网爬动, 并逐步穿越逃逸口(图版-9);当个体一侧通过网目逃逸口后身体会快速抖动, 避免两螯或步足与网目缠住。由于逃逸口的方向与笼底相倾斜, 个体需要花费更多的时间和精力调整姿态和位置以适应逃逸口; 当较大个体被逃逸口卡住时一部分会通过调整侧身方向继续尝试逃逸, 也有部分会尝试用力通过, 逃逸口相邻网目变形(图版-9)表明个体被卡后逃逸口形状已经发生了变化; 未观察到个体有横向移动(受限于网目横向较小)和剧烈抖动等行为。成功穿越的过程持续时间为(6.05±4.34)s。单次企图穿越网目逃逸口的行为中仅有57.1%(=24/42)成功逃逸, 但逃逸率为100%(=24/24)。
3 讨论水池观察实验虽然存在无法模拟野外真实环境要素、个体生理状况及个体和群体间的行为影响等缺点[26], 但同时具备环境条件可控、可重复、群体丰度已知、所有个体都暴露于饵料气味范围且有足够的时间去接近蟹笼、观察操作方便、结果更加精准等优点[27-28], 是甲壳动物对渔具行为反应研究中较为常用的试验方法[19, 29-30]。此次试验采用了水池观察方法, 系统描述了入笼的三疣梭子蟹对笼壶渔具刚性矩形逃逸口和网衣破目逃逸口的反应行为及相互之间的差异, 实验结果对于了解笼内三疣梭子蟹个体的逃逸行为、进而优化蟹笼渔具的结构、评价生物降解材料在我国近海蟹笼渔业中的应用前景等具有一定的参考意义。当然, 受三疣梭子蟹野外群体取样、观察装备等实验条件的限制, 且不同逃逸口的行为对比试验并不完全同步, 因此群体观察结果并不能完全反映实际海洋环境下三疣梭子蟹对不同类型逃逸口的反应行为, 仍需开展进一步海上观察和渔获试验加以验证。
3.1 逃逸行为阶段的行为差异及原因蟹类、龙虾等海洋甲壳动物具有动作灵活、行为多样的特点, 对笼壶渔具不同部分以及在不同阶段行为各异、反应复杂。为了能准确或定量描述甲壳动物对笼壶的行为, 便于行为比较, 很多学者将甲壳动物在笼壶内外的连续行为离散并做相应的定义, 进而探索各阶段行为特征及其主要影响因素及机制, 这已成为甲壳动物行为观察的常用研究方法[26, 31-33]。此研究中, 根据三疣梭子蟹入笼至逃逸的时间顺序, 结合笼内个体对不同类型逃逸口的反应行为特征, 将个体整体行为划分为“探索”、“发现并接近逃逸口”、“企图逃逸”和“穿越逃逸口”等4个阶段并作相应定义, 进而通过量化不同阶段的行为指标比较笼内个体对刚性矩形逃逸口和网衣破目2种不同逃逸装置在不同行为阶段下的行为特点和差异。
在探索行为阶段, 逃逸口类型会影响三疣梭子蟹对蟹笼探索行为发生的时间和频率。水下观察发现, 入笼的三疣梭子蟹在刚性逃逸口实验蟹笼中的探索行为更早发生且更集中(入笼后的0~5h), 而在网目逃逸口蟹笼中个体的探索时间跨度更大。引起这一行为差异的主要原因是不同类型的逃逸装置对笼内个体的视觉刺激差异。三疣梭子蟹具有一定的视觉能力, 喜弱光、畏强光[34], 对光强的变化较为敏感[35]; 颜色视觉方面, 虽然郑微云等[35]认为三疣梭子蟹眼睛只有单一感受系统, 缺乏颜色分辨能力, 但CRONIN等[36]认为多数十足目甲壳动物是二色性视觉, 能分辨不同波长的光, 且波长较长的红光对甲壳动物的行为影响相对较小[37]。此外, 此次水下观察实验条件相对简陋, 无法完全营造暗室环境, 而构建刚性逃逸口时采用的基板颜色为白色(图版-2,4,5,8), 与背景具有较大的反差, 加之使用了红外摄像装备, 造成刚性矩形逃逸口可能对笼内个体产生了较强的视觉刺激, 进而可能影响并制约了笼内个体的探索行为; 而网目逃逸口与背景差异不大(图版-3,6,7,9), 个体受到的视觉刺激相对较小, 需要更多的探索行为和次数才能发现网衣上的破目。
在发现并接近逃逸口阶段, 水下观察结果表明, 笼内的三疣梭子蟹能准确定位、发现并接近2种不同的逃逸口, 但不同实验蟹笼中个体探索行为的差异造成了个体发现刚性矩形和网目逃逸口时距离入笼的平均时间(刚性矩形逃逸口2.30h和网目逃逸口6.13h)和平均次数均存在一定差异。网目逃逸口蟹笼中的个体在持续不断的探索行为中, 定位和发现逃逸口的平均次数为(5.05±1.15)次, 大于刚性逃逸口蟹笼中的(3.09±1.29)次, 表明了三疣梭子蟹具有积极的逃逸行为, 而明显的矩形逃逸口视觉刺激反而可能会限制个体的逃逸或觅食行为, 从这一点出发, 刚性逃逸口的设计和制作材选择时, 应尽量降低逃逸装置与背景的反差。
在企图逃逸阶段, 多数成功定位、发现和接近逃逸口的三疣梭子蟹会尝试穿越逃逸口, 但仍有约15%的个体会选择离开逃逸口, 且逃逸口的类型对此行为没有显著影响。笼内个体发现逃逸口后不愿穿越的真实原因难以掌握, 一方面可能是因为龙虾和蟹类等甲壳动物本身具有穴居习性, 会将笼壶作为避难所而不愿逃逸[10, 27-28, 38]; 另一方面此次实验样本均来自于养殖群体, 在生长发育过程中长期处于饵料充沛、缺乏捕食者威胁且群体密度较高的环境, 三疣梭子蟹本身的逃逸意愿已被弱化, 且手工放入的个体较自行入笼的个体在笼内的行为更加平静[32]。相比刚性矩形逃逸口, 三疣梭子蟹在面对网目逃逸口时停留了更多的时间后才企图穿越逃逸口, 一方面可能是因为刚性逃逸口面积更大、位置更低、视觉刺激更强, 个体更容易作出行为反应, 另一方面可能是因为刚性逃逸口整体平行于笼底, 而网目逃逸口则与笼底相倾斜导致个体行为差异。甲壳类坚硬的头胸甲和行为的敏捷性使其能够定向到最有利的逃生位置, 因此笼壶渔具上使用刚性BRD较柔性BRD具有更加精确的渔获选择性能[29, 39-40]。通常认为, 个体能否通过逃逸口取决于其身体最大截面尺寸与逃逸口对应尺寸之间的匹配关系[29, 41]。因此在穿越逃逸口阶段, 对于刚性矩形逃逸口, 由于三疣梭子蟹采用侧身的姿态穿越逃逸口, 当逃逸口宽度足够时, 决定个体能否逃逸的对应尺寸为甲高与逃逸口高度。此次水下观察实验中, 几乎所有被刚性矩形逃逸口卡住的个体均未能穿越而返回笼内, 这也说明刚性逃逸口对个体尺寸具有较为精确的选择性能。而对于网目逃逸口, 由于其柔性结构和倾斜于笼底的方向, 水下观察难以确定到底是网目开口高度还是宽度限制了个体的逃逸, 但可以发现当三疣梭子蟹穿越逃逸口被卡时, 相邻网目形状发生变形(图版-9), 表明个体的穿越行为导致了逃逸口的形状发生了变化, 这也说明柔性逃逸口难以精确选择个体尺寸。当然, 由于刚性逃逸口和网目逃逸口在材料、尺寸大小以及逃逸口的方向等方面都存在差异, 无法定量比较逃逸口类型对个体逃逸成功率的影响, 有待进一步深入观察研究。
3.2 网目逃逸口的应用前景ELDRIDGE等[42]提出, 笼壶渔具选用兼捕减少装置(BRD)应满足能保留目标渔获、减少兼捕渔获、结构简单和成本低廉等技术要素, 这一适用标准在后续各类渔具BRD的应用中得到了广泛认可。甲壳动物笼壶渔业中, 使用最小网目尺寸是渔业管理有效技术手段之一[43-44], 这表明甲壳动物入笼后具有穿越网目逃逸的行为习性; 相比刚性逃逸装置, 在笼壶渔具的网衣上使用破目实现网目逃逸口在技术上更容易实现, 然而, 即便研究人员已开展了大量有关甲壳动物对笼壶的行为反应的观察[31, 38, 45], 但对于个体能否以及如何穿越网目进入或者逃离的行为观察仍相对较少, ZHOU等[40]发现笼外的拟石蟹(Paralithodes camtschaticus)会将螯伸入网目企图入笼; BARBER等[46]发现笼内黄道蟹(Cancer magister)除穿越逃逸口以外, 也会尝试将螯伸入网目、调整头胸甲方向穿越网目实现逃逸; WATSON等[47]和GROENEVELD等[28]发现小个体龙虾(Homarus americanus)和岩龙虾(Jasus lalandii)能轻易穿越网目逃逸, 但未具体描述其逃逸行为。对于三疣梭子蟹, 吴常文[21]通过水池观察也发现了蟹笼网目较大时, 个体会将鳌足直接伸入网目以钳取食物, 且放大网目可以释放幼蟹。此次通过水下观察结果表明, 虽然笼内的三疣梭子蟹个体对不同类型的逃逸口的反应行为存在一定的差异, 但个体能在低照度下发现并精确定位到不同位置和不同尺寸的网目逃逸口并最终逃逸, 表明网衣破目形成的网目逃逸口具有成为东海近海蟹笼渔具BRD的潜力。
此外, 笼壶渔具丢失后形成的幽灵捕捞问题一直是困扰笼壶渔业的生态难题[8-12], 在许多笼壶渔业中管理者推荐使用生物可降解材料制作网片、或使用可降解材料或有机材料网线绑扎各类BRD[12, 16, 25, 29, 48]。此次水下观察结果从一个方面反映出降解材料缓解笼壶渔具幽灵捕捞问题方面具有可操作性。当然, 相比刚性释放口, 网衣逃逸口作为柔性体容易变形、材料本身具有一定的伸缩性, 且同一笼壶各处的网衣可能缩结不同, 其对捕捞对象的精准选择性难以实现[28, 32-33], 需要进一步开展深入研究。
| [1] |
俞存根, 宋海棠, 姚光展, 等. 东海大陆架海域经济蟹类种类组成和数量分布[J]. 海洋与湖沼, 2006, 37(1): 53-60. YU C G, SONG H T, YAO G Z, et al. Composition and distribution of economic crab species in the East China sea[J]. Oceanologia et Limnologia Sinica, 2006, 37(1): 53-60. DOI:10.3321/j.issn:0029-814X.2006.01.009 |
| [2] |
俞存根, 宋海棠, 姚光展. 东海大陆架海域蟹类资源量的评估[J]. 水产学报, 2004, 28(1): 41-46. YU C G, SONG H T, YAO G Z. Assessment of the crab stock biomass in the continental shelf waters of the East China Sea[J]. Journal of Fisheries of China, 2004, 28(1): 41-46. |
| [3] |
农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会. 2020中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2020. Rural Fisheries Administration, National Fisheries Technology Extension Center, China Society of Fisheries. 2020 China fishery statistical yearbook[M]. Beijing: China Agriculture Press, 2020. |
| [4] |
贺舟挺, 刘子藩, 周永东. 浙江蟹笼渔业现状的探讨[J]. 浙江海洋学院学报(自然科学版), 2007, 26(1): 54-58. HE Z T, LIU Z F, ZHOU Y D. Study on status of the crab pot fishery in Zhejiang province[J]. Journal of Zhejiang Ocean University (Natural Science), 2007, 26(1): 54-58. DOI:10.3969/j.issn.1008-830X.2007.01.012 |
| [5] |
俞存根, 宋海棠, 姚光展, 等. 浙江近海蟹类资源合理利用研究[J]. 海洋渔业, 2003, 25(3): 136-141. YU C G, SONG H T, YAO G Z, et al. Study on rational utilization of crab resources in the inshore waters of Zhejiang[J]. Marine Fisheries, 2003, 25(3): 136-141. DOI:10.3969/j.issn.1004-2490.2003.03.008 |
| [6] |
张健, 王忠秋, 管卫兵. 近海蟹笼两种主要渔获蟹类的渔业生物学特性[J]. 大连海洋大学学报, 2015, 30(5): 540-545. ZHANG J, WANG Z Q, GUAN W B. Fishery biology of two crab species caught by offshore pots[J]. Journal of Dalian Ocean University, 2015, 30(5): 540-545. |
| [7] |
李明爽. 浙江发布海洋渔业资源重点保护品种最小可捕过渡性规格标准[J]. 中国水产, 2017(3): 23. LI M S. Zhejiang provice has published the transitional standard for minimum catchable specification of key conservational species of marine fishery stocks[J]. China Fisheries, 2017(3): 23. |
| [8] |
SMOLOWITZ R J. Trap design and ghost fishing: an overview[J]. Marine Fisheries Review, 1978, 40(5/6): 2-8. |
| [9] |
BREEN P A. A review of ghost fishing by traps and gillnets[C]//Proceedings of the Second International Conference on Marine Debris. Hawaii Honolulu, 1990: 571-599.
|
| [10] |
GUILLORY V. Ghost fishing by blue crab traps[J]. North American Journal of Fisheries Management, 1993, 13(3): 459-466. DOI:10.1577/1548-8675(1993)013<0459:GFBBCT>2.3.CO;2 |
| [11] |
MATSUOKA T, NAKASHIMA T, NAGASAWA N. A review of ghost fishing: scientific approaches to evaluation and solutions[J]. Fisheries Science, 2005, 71(4): 691-702. DOI:10.1111/j.1444-2906.2005.01019.x |
| [12] |
MACFADYEN G, HUNTINGTON T, CAPPELL R. Abandoned, lost or otherwise discarded fishing gear[R]. Rome, Italy: FAO, 2009.
|
| [13] |
ROTHERHAM D, JOHNSON D D, MACBETH W G, et al. Escape gaps as a management strategy for reducing bycatch in net-covered traps for the giant mud crab Scylla serrata[J]. North American Journal of Fisheries Management, 2013, 33(2): 307-317. DOI:10.1080/02755947.2012.760502 |
| [14] |
UHLMANN S S, BROADHURST M K. Mitigating unaccounted fishing mortality from gillnets and traps[J]. Fish and Fisheries, 2015, 16(2): 183-229. DOI:10.1111/faf.12049 |
| [15] |
BROADHURST M K, MILLAR R B. Relative ghost fishing of portunid traps with and without escape gaps[J]. Fisheries Research, 2018, 208: 202-209. DOI:10.1016/j.fishres.2018.07.018 |
| [16] |
GANDY R L, CROWLEY C E, LEONE E H, et al. Increasing the selectivity of the stone crab Menippe mercenaria trap by the addition of a cull ring[J]. North American Journal of Fisheries Management, 2018, 38(6): 1275-1283. DOI:10.1002/nafm.10232 |
| [17] |
BROADHURST M K, MILLAR R B, HUGHES B. Performance of industry-developed escape gaps in Australian Portunus pelagicus traps[J]. Fisheries Research, 2017, 187: 120-126. DOI:10.1016/j.fishres.2016.11.013 |
| [18] |
BUTCHER P A, LELAND J C, BROADHURST M K, et al. Giant mud crab (Scylla serrata): relative efficiencies of common baited traps and impacts on discards[J]. ICES Journal of Marine Science, 2012, 69(8): 1511-1522. DOI:10.1093/icesjms/fss109 |
| [19] |
HAVENS K J, BILKOVIC D M, STANHOPE D, et al. Location, location, location: the importance of cull ring placement in blue crab traps[J]. Transactions of the American Fisheries Society, 2009, 138(4): 720-724. DOI:10.1577/T08-168.1 |
| [20] |
BROADHURST M K, MILLAR R B. Configuring escape gaps in recreational rectangular traps to improve size selection for eastern rock lobster, Sagmariasus verreauxi[J]. Fisheries Research, 2018, 207: 182-186. DOI:10.1016/j.fishres.2018.05.027 |
| [21] |
吴常文. 一种资源保护型蟹笼的初步试验[J]. 海洋渔业, 1996, 18(3): 114-116. WU C W. Preliminary tests on a kind of crab pot for protecting resources[J]. Marine Fisheries, 1996, 18(3): 114-116. |
| [22] |
张洪亮, 徐汉祥, 黄洪亮, 等. 东海区蟹笼渔具的选择性研究[J]. 水产学报, 2010, 34(8): 1277-1284. ZHANG H L, XU H X, HUANG H L, et al. Selectivity of crab pot for Portunus trituberculatu in the East China Sea region[J]. Journal of Fisheries of China, 2010, 34(8): 1277-1284. |
| [23] |
白伦, 张新峰, HE P G, 等. 基于张网渔获的蟹笼渔具对三疣梭子蟹的相对选择性[J]. 中国水产科学, 2015, 22(2): 340-346. BAI L, ZHANG X F, HE P G, et al. Relative selectivity of crab pots for Portunus trituberculatus based on stownet catch in the East China Sea[J]. Journal of Fishery Sciences of China, 2015, 22(2): 340-346. |
| [24] |
ZHANG J, PEI Z W, HE P G, et al. Effect of escape vents on retention and size selectivity of crab pots for swimming crab Portunus trituberculatus in the East China Sea[J]. Aquaculture and Fisheries, 2021, 6(4): 340-347. DOI:10.1016/j.aaf.2020.04.008 |
| [25] |
WINGER P D, LEGGE G, BATTEN C, et al. Evaluating potential biodegradable twines for use in the snow crab fishery off Newfoundland and Labrador[J]. Fisheries Research, 2015, 161: 21-23. DOI:10.1016/j.fishres.2014.06.007 |
| [26] |
JURY S H, HOWELL H, O'GRADY D F, et al. Lobster trap video: in situ video surveillance of the behaviour of Homarus americanus in and around traps[J]. Marine and Freshwater Research, 2001, 52(8): 1125-1132. DOI:10.1071/MF01096 |
| [27] |
MILLER R J, ADDISON J T. Trapping interactions of crabs and American lobster in laboratory tanks[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1995, 52(2): 315-324. DOI:10.1139/f95-032 |
| [28] |
GROENEVELD J C, KHANYILE J P, SCHOEMAN D S. Escapement of the cape rock lobster (Jasus lalandii) through the mesh and entrance of commercial traps[J]. Fishery Bulletin, 2005, 103(1): 52-62. |
| [29] |
WINGER P D, WALSH P J. The feasibility of escape mechanisms in conical snow crab traps[J]. ICES Journal of Marine Science, 2007, 64(8): 1587-1591. DOI:10.1093/icesjms/fsm125 |
| [30] |
ARCHDALE M V, ANASCO C P, KAWAMURA Y, et al. Effect of two collapsible pot designs on escape rate and behavior of the invasive swimming crabs Charybdis japonica and Portunus pelagicus[J]. Fisheries Research, 2007, 85(1/2): 202-209. |
| [31] |
SMITH G S, SUMPTON W D. Behavior of the commercial sand crab Portunus pelagicus (L.) at trap entrances[J]. Asian Fisheries Science, 1989, 3: 101-113. |
| [32] |
ZHOU S J, SHIRLEY T C. Behavioural responses of red king crab to crab pots[J]. Fisheries Research, 1997, 30(3): 177-189. DOI:10.1016/S0165-7836(97)00005-2 |
| [33] |
BERGSHOEFF J A, MCKENZIE C H, BEST K, et al. Using underwater video to evaluate the performance of the fukui trap as a mitigation tool for the invasive european green crab (Carcinus maenas) in Newfoundland, Canada[J]. PeerJ, 2018, 6: e4223. DOI:10.7717/peerj.4223 |
| [34] |
罗会明, 郑微云. 水平光梯度下三疣梭子蟹趋光反应的实验研究[J]. 海洋学报, 1981, 3(2): 300-306. LUO H M, ZHENG W Y. Experimental studies on the phototactic reaction of portunid (Portunus trituberculatus, (Miers)) in horizontal photogradients[J]. Acta Oceanologica Sinica, 1981, 3(2): 300-306. |
| [35] |
郑微云, 柴敏娟. 三疣梭子蟹视觉特性的电生理研究[J]. 海洋与湖沼, 1990, 21(5): 490-494. ZHENG W Y, CHAI M J. Electrophysiological studies on the visual characteristics of Portunus trituberculatus (Miers)[J]. Oceanologia et Limnologia Sinica, 1990, 21(5): 490-494. |
| [36] |
CRONIN T W, HARIYAMA T. Spectral sensitivity in crustacean eyes[M]//WIESE K. The Crustacean Nervous System. Berlin, Heidelberg: Springer, 2002: 499-511.
|
| [37] |
CRONIN T W. Photoreception in marine invertebrates[J]. American Zoologist, 1986, 26(2): 403-415. DOI:10.1093/icb/26.2.403 |
| [38] |
STURDIVANT S K, CLARK K L. An evaluation of the effects of blue crab (Callinectes sapidus) behavior on the efficacy of crab pots as a tool for estimating population abundance[J]. Fishery Bulletin, 2011, 109(1): 48-55. |
| [39] |
MILLER R J. Effectiveness of crab and lobster traps[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1990, 47(6): 1228-1251. DOI:10.1139/f90-143 |
| [40] |
ZHOU S J, SHIRLEY T C. Performance of two red king crab pot designs[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1997, 54(8): 1858-1864. DOI:10.1139/f97-094 |
| [41] |
TREBLE R J, MILLAR R B, WALKER T I. Size-selectivity of lobster pots with escape-gaps: application of the select method to the southern rock lobster (Jasus edwardsii) fishery in Victoria, Australia[J]. Fisheries Research, 1998, 34(3): 289-305. DOI:10.1016/S0165-7836(97)00072-6 |
| [42] |
ELDRIDGE P J, BURRELL JR V, STEELE G. Development of a self-culling blue crab pot[J]. Marine Fisheries Review, 1979, 41(11/12): 21-27. |
| [43] |
BRČIĆ J, HERRMANN B, MAŠANOVIĆ M, et al. Size selection of Nephrops norvegicus (L.) in commercial creel fishery in the Mediterranean Sea[J]. Fisheries Research, 2018, 200: 25-32. DOI:10.1016/j.fishres.2017.12.006 |
| [44] |
BROADHURST M K, SMITH T M, MILLAR R B, et al. Cumulative selectivity benefits of increasing mesh size and using escape gaps in australian Portunus armatus traps[J]. Fisheries Management and Ecology, 2019, 26(4): 319-326. DOI:10.1111/fme.12351 |
| [45] |
ADDISON J, BANNISTER R. Quantifying potential impacts of behavioral factors on crustacean stock monitoring and assessment: modeling and experimental approaches[J]. Canadian Special Publication of Fisheries and Aquatic Sciences, 1998, 125: 167-177. |
| [46] |
BARBER J S, COBB J S. Qualitative observations of dungeness crabs, Cancer magister, in and around traps: evidence of resource guarding and clustering[J]. Marine and Freshwater Behaviour and Physiology, 2009, 42(2): 135-146. DOI:10.1080/10236240902860011 |
| [47] |
WATSON W, JURY S H. The relationship between american lobster catch, entry rate into traps and density[J]. Marine Biology Research, 2013, 9(1): 59-68. DOI:10.1080/17451000.2012.727430 |
| [48] |
HÉBERT M, MIRON G, MORIYASU M, et al. Efficiency and ghost fishing of snow crab (Chionoecetes opilio) traps in the gulf of St. Lawrence[J]. Fisheries Research, 2001, 52(3): 143-153. DOI:10.1016/S0165-7836(00)00259-9 |
2. Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai 201306, China;
3. National Engineering Research Centre for Oceanic Fisheries, Shanghai 201306, China;
4. Shanghai Aquatic Wildlife Conservation and Research Center, Shanghai 200092, China;
5. Joint Laboratory for Monitoring and Conservation of Aquatic Living Resources in the Yangtze Estuary, Shanghai 202162, China;
6. East China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Shanghai 200090, China