 2022,
Vol. 31
2022,
Vol. 31



2. 大洋渔业资源可持续开发教育部重点实验室, 上海 201306;
3. 国家远洋渔业工程技术研究中心, 上海 201306;
4. 农业农村部大洋渔业开发重点实验室, 上海 201306
海洋渔业资源量在时间上存在波动, 与物种生活史特征、生境及捕捞等因素关系密切[1-2]。对资源量波动的描述主要依据研究的时间尺度包括季节性(seasonal)、年间(annual, 一年与一年的比较)、年际(interannual, 多年间的比较)和年代际(interdecadal, 或称为十年代际, 大于10年的多年间比较)变动等[3-6]。在不同时间尺度上, 资源量波动程度不一致, 且影响因素不同[6]。例如, 季节性的资源量变动被认为与物种洄游[7-8]、生境(如水温、食物条件等)[9]及捕捞(如禁渔期等)[10]等因素有关; 而长期的资源量波动则与气候变化等因素关联密切[6]。研究发现, 一些种类资源量的长期变动常与某些气候指数呈现明显的“共振关系”: 例如MANTUA等[5]报道太平洋年代际涛动(pacific decadal oscillation, PDO)指数为正值期时(即PDO暖期), 北太平洋三文鱼种类资源量为高位, 而PDO指数为负值期时(即PDO冷期), 情况则相反; HERRIZ等[11]发现, 东北大西洋挪威龙虾(Nephrops norvegicus)的资源丰度指数与滞前6.5年的北大西洋涛动(north atlantic oscillation)指数呈显著的负交相关关系。相关研究表明, 资源量的波动存在不同时间尺度的特征, 但是目前鲜有研究结合相关影响因素对不同时间尺度的波动特征进行全面的分析, 研究结果可为渔业科学管理及探索渔业资源变动的主要成因提供一个全新的角度。
秘鲁鳀(Engraulis ringens)是栖息于东南太平洋南美洲沿岸的一种小型中上层鱼类, 是世界上产量最大的单鱼种渔业[12], 为我国进口鱼粉的主要来源[13]。研究表明, 其资源量波动受到厄尔尼诺拉尼娜事件[14-15]及东南太平洋洪堡洋流生态系统周期性变动[16-17]的影响。但是相关研究结果仅基于整体资源量时间序列的观测和分析, 秘鲁鳀不同时间尺度的波动特征及规律对其生境和气候因素是否存在差异?因此, 研究以1985—2021年间秘鲁鳀资源量时间序列为研究对象, 利用自适应噪声的完全集合经验模态分解(complete empirical mode decomposition with adaptive noise, CEEMDAN)的方法[18], 提取秘鲁鳀资源量波动的不同时间尺度特征, 并评价各时间尺度波动对总体资源波动的贡献程度; 此外使用交相关分析的方法探究厄尔尼诺和拉尼娜事件对不同时间尺度下秘鲁鳀资源量波动的影响。研究结果将为研究秘鲁鳀及其他种类渔业资源波动及其成因提供理论和方法支撑。
1 材料与方法 1.1 数据来源秘鲁鳀资源量数据来源于秘鲁国家海洋局(Instituto del Mar del Perú, IMARPE, http://www.imarpe.pe/imarpe)和秘鲁生产部(Ministerio de la Producción Perú,https://www.gob.pe/produce)。秘鲁鳀每年的捕捞渔汛分为2次, 分别为当年的4至8月和11月至翌年的1月。在每次捕捞渔汛开始前的3月和10月左右, IMARPE指派声学调查船对秘鲁沿岸秘鲁鳀资源进行调查并估算资源量, 根据估算结果决定当季渔汛的可捕量及捕捞天数[19]。研究[16]表明, 秘鲁鳀在东南太平洋洪堡洋流生态系统中长期处于优势种的地位且大量集群; 秘鲁鳀资源声学调查始于20世纪70年代末, 调查技术成熟, 且资源量估计结果已得到拖网调查结果的印证[20]; 同时基于声学估算的资源量进行秘鲁鳀资源管理被认为是渔业管理中较为成功的案列[21]。因此认为相关声学评估结果可较好地反映秘鲁鳀真实的资源量水平。研究使用的秘鲁鳀资源量数据时间范围为1985—2021年, 由于每年3月和10月左右各调查1次, 因此认为其结果分别为当年上、下半年的资源量(即资源量数据的时间分辨率为0.5年)。
环境数据为表征厄尔尼诺拉尼娜事件的海洋尼诺指数(oceanic niño index, ONI)和Niño1+2区海表面温度距平(sea surface temperature anomaly, SSTA, 下文以Niño1+2指数表示), 数据来源于美国国家大气局(NOAA)的物理科学实验室网站(https://psl.noaa.gov/data/climateindices/list/), 数据的时间分辨率为月, 时间范围为1984—2021年。
1.2 分析方法 1.2.1 资源波动时间尺度特征分析研究使用自适应噪声的完全集合经验模态分解(CEEMDAN)的方法对秘鲁鳀资源量波动的不同时间尺度特征进行分析。CEEMDAN是经验模态分解(empirical mode decomposition, EMD)的一种方法, 主要目的为对时间序列信号(T)进行去噪并解析自身的不同时间尺度波动特征; 其基本原理与EMD相同, 通过希尔伯特-黄转换(hilbert-huang transform, HHT)方法将时间序列逐级筛选和分解为若干个本征模态函数(intrinsic mode function, IMFi, i=1, 2, 3…)分量和一个残差项(residual, RES), 最终使得各IMFi分量能够反映时间序列波动的主要时间尺度及变动规律[18, 22]。但CEEMDAN为EMD的改进方法, 通过在分解过程中对时间序列添加白噪声并进行自适应分解的办法, 改进了原始EMD方法模态重叠、分解效率慢等问题[18], 并有:
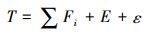 (1)
(1)
式中: Fi为分解得到的第i个本征模态函数IMFi, 反映了特定时间尺度的波动规律; E为残差项RES, 其随时间变动不呈现波动性, 反映了时间序列的趋势特征; ε为残余噪声, 其时间上的变动通常小于IMFi和RES几个量级, 通常可以忽略不计。分解的具体计算算法可见文献[18, 22], 算法利用R软件(版本4.1.0)中的Rlibeemd包。分解过程中的参数设置为集合总数(ensemble size): 50, 噪声强度(noise strength): 原始总资源量时间序列标准差的0.2倍。
将资源量时间序列进行分解后, 利用准周期(quasi-period)对各IMFi分量的时间特征进行定量化描述。准周期定义: 各IMFi分量上相邻极大(小)值间(即两个波峰或波谷间)的时间跨度。根据不同时间尺度的定义可认为: 当准周期为1年时, 则IMFi分量包含了季节性波动的特征; 当准周期为1.5至2年时, 则IMFi分量包含了年间波动的特征; 当准周期为2.5至9.5年时, 则IMFi分量包含了年际波动的特征; 准周期大于10年时, 则IMFi分量包含了年代际波动的特征。
1.2.2 不同时间尺度波动重要性评价使用3种方法对各IMFi分量及RES对总资源量时间序列的贡献程度进行评价。第一, 通过计算各IMFi分量中各准周期的极值差, 极值差定义为一个准周期内最大值和最小值之差, 计算各IMFi分量的极值差平均值, 并与整体资源量时间序列的极值差平均值进行对比, 其值越接近则表示该IMFi越能反映原资源量时间序列的波动程度; 第二, Pearson相关系数及其检验, Pearson相关系数衡量了各IMFi分量及RES与总资源量时间序列同时超过或低于序列均值的概率, 概率越大, 相关系数显著且其值越接近于1;第三, Kendall系数, Kendall衡量了各IMFi分量及RES与总资源量时间序列在同一时点与上一时点相同时变化方向相同的概率, 衡量了各IMFi分量及RES与总资源量时间序列; Pearson相关系数和Kendall系数分别从趋势的程度和方向两个方面衡量了各IMFi分量及RES与总资源量时间序列的相关性[23], 相关系数越高, 则表明该IMFi(或RES)更能够驱动整体资源量时间序列的走势。
1.2.3 厄尔尼诺拉尼娜事件对各IMFi分量及RES的影响分析ONI是目前国际上用来表征厄尔尼诺拉尼娜事件通用的一个指标, 而近期发表的秘鲁鳀资源量的预测模型通常以Niño1+2指数表征厄尔尼诺拉尼娜事件并作为一个因子加入预测中[24-25], 但并未解释原因。因此, 研究使用交相关分析的办法将这2个因子与各IMFi分量的响应关系分别进行分析, 以探究这2个因子的影响差异。研究首先将ONI和Niño1+2指数的时间尺度转换为0.5年, 以保证其与资源量数据的时间分辨率一致, 依据声学调查的时间, 分别平均去年10月至当年3月和当年3月至9月的月度ONI和Niño1+2指数, 作为对应上、下半年资源量的厄尔尼诺和拉尼娜事件情况。由于秘鲁鳀为3年生个体[12], 因此假设当次调查3年内的厄尔尼诺和拉尼娜事件都对秘鲁鳀资源量产生影响, 将交相关分析中的滞后影响上限设置为3年, 即只检验滞前0~2.5年内ONI和Niño1+2指数与各IMFi分量的交相关关系及其显著性。
2 结果 2.1 秘鲁鳀资源量的时间变动特征1985—2005年, 秘鲁鳀资源量呈现波动上升的趋势, 2005年后, 资源量在700万~800万t的水平上下波动。资源量最低和最高的时间分别为1987年上半年(117万t)和2005年上半年(1 270万t); 可以看到, 秘鲁鳀资源量波动剧烈, 尤其是在2014年, 上半年的资源量为970万t, 下半年的资源量仅为145万t, 随后在2015年的上半年, 资源量迅速回升至940万t(图 1)。

|
图 1 1985—2021年秘鲁鳀资源量时间序列 Fig. 1 Time series of anchoveta (Engraulis ringens) biomass during 1985-2021 |
秘鲁鳀资源量时间序列经过CEEMDAN分解后, 得到5个IMFi分量(IMF1至IMF5)及一个RES(图 2)。随着IMFi分量级数i的增加, 平均准周期增加且准周期出现次数减少(表 1), 可反映秘鲁鳀资源量由高频至低频的周期性波动。其中, IMF1随时间波动最为剧烈, 准周期为1~3年, 出现准周期小于等于2年的次数最多, 为38次, 其中有12次准周期为1年, 可知, IMF1主要表现出秘鲁鳀资源波动的季节和年间特征; IMF2和IMF3的平均准周期相似, 分别为2.6年和3.2年, 但IMF3与IMF2相比, 包含几个时间跨度较大的准周期, 如1993年下半年至1999年上半年(先增加后减少)和2012年上半年至2018年下半年(先减少后增加), 而IMF2的准周期最大值仅为3.5年, 表明IMF2和IMF3包含了秘鲁鳀资源波动的季节(IMF3)、年间(IMF2和IMF3)和年际(IMF2和IMF3)特征, 但是IMF3其波动存在由高频向低频的过渡的特征; IMF4和IMF5与前3个IMFi相比更能体现出秘鲁鳀资源量波动的低频特征, 平均准周期分别为5.5年和6.6年, 出现准周期的次数减少, 分别为12次和7次, IMF4的准周期为2~8年, 表明IMF4主要表现出秘鲁鳀资源波动的年间和年际特征; IMF5的准周期为4~14年, 短时间跨度的准周期发生在2000年以前, 如1990年上半年至1994年上半年, 但增加减少趋势较为平缓, 这表明IMF5表现出秘鲁鳀资源波动的年际特征的同时已显示出更为低频的年代际特征(图 2)。RES则反映了秘鲁鳀资源量在1985—2021年间的趋势性特征, 先上升后保持平缓, 这与对整体时间序列的观测结果一致(图 1)。

|
图 2 自适应噪声的完全集合经验模态分解对秘鲁鳀资源量时间序列分解结果 Fig. 2 Results of Complete Empirical Mode Decomposition with Adaptive Noise (CEEMDAN) to the anchoveta(Engraulis ringens) biomass time series |

|
表 1 各IMFi分量波动的时间尺度特征(准周期和出现次数) Tab.1 Time-scale features of each IMFi fluctuation (quasi-period and ouurrence number) |
1985—2015年间, 秘鲁鳀资源量序列的极值差平均值为371.49万t, IMFi分量中(表 2), 极值差平均值最大的是IMF1(232.67万t), 其次为IMF2(205.21万t), IMF1和IMF2的极值差平均值也与总资源量时间序列也最为接近, 其他3个IMFi分量的极值差平均值与总资源量时间序列相差较大, 均小于100万t, 此外趋势项RES的变动也较为明显(图 2), 最大值和最小值之差为563.09万t; Pearson相关系数和Kendall相关系数及检验均表明, 各IMFi和RES与总资源量时间序列均呈显著的相关关系, 其中RES的相关系数值最大, 分别为0.70(Pearson相关系数)和0.43(Kendall相关系数); 各IMFi分量中, IMF1与总资源量时间序列的Pearson相关系数最大, 为0.54, 其次为IMF4, 为0.41;Kendall相关系数的结果与Pearson相关系数的相似, 各IMFi分量中, IMF1与总资源量时间序列的Kendall相关系数最大, 为0.34, 其次为IMF4, 为0.28。

|
表 2 各IMFi分量及RES对秘鲁鳀资源波动贡献程度检验 Tab.2 Test for the contribution of each IMFi and RES to the anchoveta(Engraulis ringens) biomass fluctuation |
因此, 综合以上结果可得, IMF1和RES对秘鲁鳀资源波动的贡献程度较大, 表明其资源波动主要受到其短期高频波动及长期趋势的控制。
2.4 厄尔尼诺拉尼娜事件对各IMFi分量的影响交相关分析表明, ONI指数仅对IMF3存在滞后0.5年的影响, 其与IMF3呈现显著的负相关关系(r=-0.24, P < 0.05, 表 3)。而Niño1+2指数可对IMF2、IMF3和IMF4造成影响(表 4): Niño1+2指数与滞后0和0.5年的IMF2、IMF3和IMF4均呈显著的负交相关关系(P < 0.05), 其中Niño1+2指数与滞后0.5年的IMF3与的交相关系数最大, 为-0.39;此外, 滞前1年的Niño1+2指数还与IMF3和呈显著的负交相关关系(P < 0.05), 交相关系数分别为-0.29和-0.25;Niño1+2指数和滞后2.5年的IMF2与IMF3呈显著的正交相关关系(P < 0.05), 交相关系数分别为0.26和0.22。

|
表 3 海洋尼诺指数与各IMFi分量交相关分析结果 Tab.3 Cross-correlation results between each IMFi and oceanic Niño index (ONI) |

|
表 4 Niño 1+2指数与各IMFi分量交相关分析结果 Tab.4 Cross-correlation results between each IMFi and Niño1+2 Index |
因此, 基于不同滞后时间的交相关系数及其显著情况可知, 厄尔尼诺和拉尼娜事件可对IMF2、IMF3和IMF4的波动造成影响, 其中, IMF3与厄尔尼诺和拉尼娜事件的关系最大; 滞前1~1.5年内ONI和Niño1+2指数可作为判定秘鲁鳀资源波动的一个主要指标, 其变化可对秘鲁鳀资源量波动产生负影响; 但是, Niño1+2指数与ONI相比对秘鲁鳀资源量波动的表征作用更为明显。此外, Niño1+2指数和滞后2.5年的IMF2与IMF3呈显著的正交相关关系, 这可能与Niño1+2指数的周期性变动有关。
3 讨论渔业资源其资源量年间波动是学者讨论的热点问题, 但研究通常仅基于整体资源量时间序列的观测和分析。本研究以多年秘鲁鳀资源量波动为例, 利用CEEMDAN的方法对其波动的不同时间尺度特征进行分析, 分解出代表 5种不同频次周期性波动的IMF1~IMF5和一个趋势项RES(图 2), 其中周期性变动(IMF2~IMF4)与厄尔尼诺拉尼娜事件相关指数存在显著的交相关关系(表 2和表 3), 与前人的研究[14-15]结果一致。此外对秘鲁鳀资源量[16-17]和补充量[26]的时间序列观测和分析均表明, 在20世纪90年代末期, 由于生态系统的周期性变动, 秘鲁鳀资源存在着由低位至高位的“跃变”, 且近年来均保持在高位的状态, CEEMDAN分解出的趋势项RES也得到相应的趋势。上述结果证明了使用CEEMDAN方法分析秘鲁鳀资源量波动的时间特征的可行性。相关研究方法可在其他渔业资源种类资源量波动的研究中进行推广。
各IMFi分量准周期的时间跨度表明(表 1), IMF1表现出秘鲁鳀资源波动的季节和年间特征, IMF2、IMF3和IMF4主要表示年间特征和年际特征, IMF5则表示年际甚至年代际的特征。总体上随着IMFi分量等级的增加, 波动从高频转至低频。但是从各IMFi分量准周期的时间跨度来看, 分解结果还是存在模态混叠的现象, 尤其是IMF3, 准周期的时间跨度为1~6年, 准周期平均值与IMF2相似(表 1), 模态混叠是目前EMD方法尚未完全解决的问题, 研究认为时间序列信号中的噪声、其他频次信号过强或频率接近都有可能造成模态混叠[27-28]。此外, 影响因素的多周期性也可能是造成模态混叠的1个原因。根据ONI数值和厄尔尼诺拉尼娜事件的定义, 1985年以来, 厄尔尼诺或拉尼娜事件的持续时间可为5~32个月, 两事件出现的时间间隔不一(例如2010年3月, 厄尔尼诺事件结束, 6月开始发生拉尼娜事件; 而2012年4月, 拉尼娜事件结束后, 直至2014年10月才开始发生厄尔尼诺事件); 且厄尔尼诺和拉尼娜事件并非交替出现(例如2020年8月—2021年5月发生了1次拉尼娜事件, 而随后8月拉尼娜事件再次发生), 这表明历史上厄尔尼诺拉尼娜事件的发生也不具备稳定时间跨度的周期。本研究发现, 厄尔尼诺拉尼娜事件时影响秘鲁鳀年间和年际波动的主要因素之一(表 3和表 4), 其中Niño1+2指数可对IMF2、IMF3和IMF4造成影响(表 4), 表明多周期性的厄尔尼诺和拉尼娜事件影响在不同时间尺度的秘鲁鳀资源波动上都可呈现, 这为EMD相关方法完全避免模态混叠的问题造成困难。目前学者针对EMD结果中模态混叠问题提出了多种办法, 如掩膜信号法[29]、多元变分模态分解法[30]、二元经验模态分解法[31]等, 在今后的研究中可利用多种办法对资源量时间序列进行分解, 比较不同方法的优劣性。
从各IMFi分量及RES的贡献程度分析中(表 2)可以得知, 秘鲁鳀的资源量变动主要受到季节或年间为表征的IMF1波动和代表其十年代际变动的趋势项RES所控制。其中RES的趋势可以用东南太平洋洪堡洋流生态系统周期性变动解释[16]。季节或年间波动为主导则主要与秘鲁鳀的生物学特征及其影响其资源变动的其他因素相关, 秘鲁鳀是一种短生命周期的r型种类[32-33], 资源变动对环境变化响应迅速[12], 如上升流强弱[34]、涡的形成[35]和饵料生物的丰度[36]等, 这些因素在年内就存在着较大的波动, 存在明显的季节性, 因此, 为了解释秘鲁鳀资源量波动, 季节性相关因素不可忽视, 需要对多种因素进行综合考虑。另外, CEEMDAN同时分解出IMF5分量(图 2), 其表现出年际甚至年代际的特征, 虽然其对总资源量时间序列波动的贡献程度相对较弱, 但表明可能存在其他大尺度因素对其资源量波动造成影响。这有待于后续结合多种气候因子进行探究。
如前所述, 厄尔尼诺和拉尼娜事件是影响秘鲁鳀资源波动的一个重要气候因子, 其可影响秘鲁鳀资源量季节至年际的波动, 其中对年间和年际波动影响最多(表 2和表 3)。通过交相关分析发现, Niño1+2指数与ONI相比, 其对秘鲁鳀资源量波动的表征作用更为明显。可能是由于厄尔尼诺事件存在不同种类型导致的。厄尔尼诺发生时暖池向东太平洋偏移, KUG等[37]研究发现, 厄尔尼诺按照暖池的分布可分为两种: 当偏移程度不足时, 水温升高区域主要在中部太平洋, 称为中部型厄尔尼诺(Central Pacific El-ni?o), 此时Niño1+2指数升高不明显; 当偏移程度足够向东时, 水温升高区域主要在东部太平洋, 称为东部型厄尔尼诺(Eastern Pacific El-Niño), 此时Niño1+2指数升高明显。Niño1+2指数升高表示秘鲁渔场北部的东太平洋赤道暖水增多, 且可开始向南部扩张至秘鲁沿岸, 结合陈芃等[38]的研究表明, 外海暖水入侵秘鲁沿岸会导致秘鲁鳀适宜生存温度区域减少, 压缩生存空间, 同时食物减少, 伴随暖水而至的天敌数量增多, 这将不利于秘鲁鳀的生存。中部型厄尔尼诺发生时则情况不同: 渔场北部的东太平洋赤道暖水向秘鲁沿岸扩张不明显, 因此对秘鲁鳀生存的危害较小。也要指出, 与厄尔尼诺事件相反, KUG等[37]后续的研究发现, 拉尼娜事件的发生不存在两种类型, 这可能是造成Niño1+2指数与ONI与各IMFi分量的交相关系数都较低的一个原因(表 3和表 4)。但是, 综合本研究的相关结果及上述厄尔尼诺事件影响秘鲁鳀资源变动的相关机理, 我们建议相关研究及国内鱼粉市场在后续估计某次渔汛季度开始前秘鲁鳀的资源状况时, 需重点关注前1~1.5年内的Niño1+2指数的变动情况, 其值的偏高可能会对秘鲁鳀资源量产生负影响。
4 小结和展望研究利用CEEMDAN的方法对1985—2021年秘鲁鳀资源量波动的时间尺度特征进行分析, 结果可与厄尔尼诺拉尼娜事件及东南太平洋生态系统周期性变动相印证; 秘鲁鳀的资源量变动主要受到季节或年间的波动(IMF1)及年代际变动的趋势项(RES)所控制; 研究并进一步讨论了厄尔尼诺拉尼娜事件对秘鲁鳀资源波动的影响, 认为Niño1+2指数对秘鲁鳀资源量波动的表征作用更为明显。相关研究结果可为学者研究秘鲁鳀及其它种类渔业资源波动及其成因提供理论和方法参考, 也为我国鱼粉进口企业把握秘鲁鳀渔业生产情况提供决策支撑。研究表明, 为解释秘鲁鳀资源量波动的不同时间尺度特征, 后续研究应综合考虑多种因素。
| [1] |
詹秉义. 渔业资源评估[M]. 北京: 中国农业出版社, 1995: 10-12. ZHAN B Y. Fish stock assessment[M]. Beijing: China Agriculture Press, 1995: 10-12. |
| [2] |
陈新军. 渔业资源与渔场学[M]. 2版. 北京: 海洋出版社, 2014: 1-173. CHEN X J. Fisheries biology and oceanography[M]. 2nd ed. Beijing: China Ocean Press, 2014: 1-173. |
| [3] |
张瑛瑛, 徐博, 张衡, 等. 南极磷虾渔场(48渔区)CPUE的年、月变化及其与海表温度、叶绿素浓度的关系[J]. 生态学杂志, 2020, 39(5): 1685-1694. ZHANG Y Y, XU B, ZHANG H, et al. Inter annual and monthly variations of catch per unit effort and the relation with sea surfacetemperature and chlorophyll concentration in fishing grounds (fishing area 48) of Antarctic krill[J]. Chinese Journal of Ecology, 2020, 39(5): 1685-1694. |
| [4] |
张志敏, 徐年军. 东海渔获量受捕捞努力与气候因子影响的年际变动分析[J]. 宁波大学学报(理工版), 2016, 29(4): 112-116. ZHANG Z M, XU N J. Effect of climate factors and fishing pressure on total fishery resources in East China Sea[J]. Journal of Ningbo University (NSEE), 2016, 29(4): 112-116. |
| [5] |
MANTUA N J, HARE S R, ZHANG Y, et al. A Pacific inter decadal climate oscillation with impacts on salmon production[J]. Bulletin of the American Meteorological Society, 1997, 78(6): 1069-1080. DOI:10.1175/1520-0477(1997)078<1069:APICOW>2.0.CO;2 |
| [6] |
CHEN P, CHEN X J, YU W, et al. Inter annual abundance fluctuations of two oceanic squids in the Pacific Ocean can be evaluated through their habitat temperature variabilities[J]. Frontiers in Marine Science, 2021, 8: 770224. DOI:10.3389/fmars.2021.770224 |
| [7] |
BRU N, PROUZET P, LEJEUNE M. Daily and seasonal estimates of the recruitment and biomass of glass eels runs (Anguilla anguilla) and exploitation rates in the Adour open estuary (Southwestern France)[J]. Aquatic Living Resources, 2009, 22(4): 509-523. DOI:10.1051/alr/2009050 |
| [8] |
MUHLING B A, BRODIE S, SMITH J A, et al. Predictability of species distributions deteriorates under novel environmental conditions in the California Current System[J]. Frontiers in Marine Science, 2020, 7: 589. DOI:10.3389/fmars.2020.00589 |
| [9] |
NATANIEL A, LOPEZ J, SOTO M. Modelling seasonal environmental preferences of tropical tuna purse seine fisheries in the Mozambique Channel[J]. Fisheries Research, 2021, 243: 106073. DOI:10.1016/j.fishres.2021.106073 |
| [10] |
ERIK G, DAGˇTEKIN M. Effect of fishery closure on discard composition of rapa whelk, Rapanavenosa (Valenciennes 1846) beam trawl fisheries in Black Sea[J]. Turkish Journal of Fisheries and Aquatic Sciences, 2021, 22(5): TRJFAS19046. |
| [11] |
HERRAIZ I G, TORRES M A, FARIÑA A C, et al. The NAO index and the long-term variability of Nephrops norvegicus population and fishery off West of Ireland[J]. Fisheries Research, 2009, 98(1/3): 1-7. |
| [12] |
陈芃, 汪金涛, 陈新军. 秘鲁鳀资源变动及与海洋环境要素的关系研究进展[J]. 海洋渔业, 2016, 38(2): 206-216. CHEN P, WANG J T, CHEN X J. Review on relationship between oceanic environment factors and population dynamics of anchoveta Engraulis ringens[J]. Marine Fisheries, 2016, 38(2): 206-216. DOI:10.3969/j.issn.1004-2490.2016.02.012 |
| [13] |
韦震. 鱼粉: 市场静待秘鲁配额公布, 国内外鱼粉价格稳定观望[J]. 当代水产, 2016, 41(11): 72-73. WEI Z. Fish meat: Market is waiting for the promulgate of anchoveta allowable catch, so the price of fish meal tends to be stable[J]. Current Fisheries, 2016, 41(11): 72-73. DOI:10.3969/j.issn.1674-9049.2016.11.025 |
| [14] |
BERTRAND A, SEGURA M, GUTIÉRREZ M, et al. From small-scale habitat loopholes to decadal cycles: a habitat-based hypothesis explaining fluctuation in pelagic fish populations off Peru[J]. Fish and Fisheries, 2004, 5(4): 296-316. DOI:10.1111/j.1467-2679.2004.00165.x |
| [15] |
ÑIQUEN M, BOUCHON M. Impact of El Niño events on pelagic fisheries in Peruvian waters[J]. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 2004, 51(6/9): 563-574. |
| [16] |
ALHEIT J, NIQUEN M. Regime shifts in the Humboldt Current ecosystem[J]. Progress in Oceanography, 2004, 60(2/4): 201-222. |
| [17] |
CHAVEZ F P, MESSIÉ M. A comparison of eastern boundary upwelling ecosystems[J]. Progress in Oceanography, 2009, 83(1/4): 80-96. |
| [18] |
TORRES M E, COLOMINAS M A, SCHLOTTHAUER G, et al. A complete ensemble empirical mode decomposition with adaptive noise[C]//Proceedings of 2011 IEEE International Conference on Acoustics, Speech and Signal Processing (ICASSP). Prague: IEEE, 2011: 4144-4147.
|
| [19] |
IMARPE. Situación del stock Norte-Centro de la Anchoveta peruana (Engraulis ringens) a Abril de 2021γperspectivas de explotaciónpara la primeratemporada de pesca del a?o[EB/OL]. (2022-01-18)[2021-04-01]. https://cdn.www.gob.pe/uploads/document/file/1832232/Anexo%20-%20Proyecto%20de%20Oficio%20339-2021-IMARPE.pdf.
|
| [20] |
SIMMONDS E J, GUTIÉRREZ M, CHIPOLLINI A, et al. Optimizing the design of acoustic surveys of Peruvian anchoveta[J]. ICES Journal of Marine Science, 2009, 66(6): 1341-1348. DOI:10.1093/icesjms/fsp118 |
| [21] |
SCHREIBER M A. The evolution of legal instruments and the sustainability of the Peruvian anchovy fishery[J]. Marine Policy, 2012, 36(1): 78-89. DOI:10.1016/j.marpol.2011.03.010 |
| [22] |
HUANG N E, SHEN Z, LONG S R, et al. The empirical mode decomposition and the Hilbert spectrum for nonlinear and non-stationary time series analysis[J]. Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences, 1998, 454(1971): 903-995. DOI:10.1098/rspa.1998.0193 |
| [23] |
ZHU B Z, WANG P, CHEVALLIER J, et al. Carbon price analysis using empirical mode decomposition[J]. Computational Economics, 2015, 45(2): 195-206. DOI:10.1007/s10614-013-9417-4 |
| [24] |
段丁毓, 陈芃, 陈新军, 等. 基于灰色系统的秘鲁鳀资源量预测模型的构建[J]. 上海海洋大学学报, 2018, 27(2): 284-290. DUAN D Y, CHEN P, CHEN X J, et al. The construction of biomass forecasting model for the anchoveta (Engraulis ringens) by the grey system model[J]. Journal of Shanghai Ocean University, 2018, 27(2): 284-290. |
| [25] |
陈芃, 陈新军, 雷林, 等. 东南太平洋秘鲁鳀资源量预报模型的构建[J]. 海洋学报, 2018, 40(1): 77-86. CHEN P, CHEN X J, LEI L, et al. Forecasting model to the anchoveta (Engraulis ringens) biomass in the Southeastern Pacific Ocean[J]. Haiyang Xuebao, 2018, 40(1): 77-86. DOI:10.3969/j.issn.0253-4193.2018.01.009 |
| [26] |
CAHUIN S M, CUBILLOS L A, ÑIQUEN M, et al. Climatic regimes and the recruitment rate of anchoveta, Engraulis ringens, off Peru[J]. Estuarine, Coastal and Shelf Science, 2009, 84(4): 591-597. DOI:10.1016/j.ecss.2009.07.027 |
| [27] |
WU Z H, HUANG N E. Ensemble empirical mode decomposition: a noise-assisted data analysis method[J]. Advances in Adaptive Data Analysis, 2009, 1(1): 1-41. DOI:10.1142/S1793536909000047 |
| [28] |
刘少康, 武英杰, 安伟伦, 等. 基于声音信号和改进MS-LMD的风电齿轮箱故障诊断[J]. 振动与冲击, 2021, 40(11): 230-239, 251. LIU S K, WU Y J, AN W L, et al. Wind turbine gearbox fault diagnosis based on sound signal and improved MS-LMD[J]. Journal of Vibration and Shock, 2021, 40(11): 230-239, 251. |
| [29] |
DEERING R, KAISER J F. The use of a masking signal to improve empirical mode decomposition[C]//Proceedings of IEEE International Conference on Acoustics, Speech, and Signal Processing, 2005. Philadelphia: IEEE, 2005: iv/485-iv/488.
|
| [30] |
REHMAN N U, AFTAB H. Multivariate variational mode decomposition[J]. IEEE Transactions on Signal Processing, 2019, 67(23): 6039-6052. DOI:10.1109/TSP.2019.2951223 |
| [31] |
RILLING G, FLANDRIN P, GONCALVES P, et al. Bivariate empirical mode decomposition[J]. IEEE Signal Processing Letters, 2007, 14(12): 936-939. DOI:10.1109/LSP.2007.904710 |
| [32] |
BERTRAND A, GERLOTTO F, BERTRAND S, et al. Schooling behaviour and environmental forcing in relation to anchoveta distribution: an analysis across multiple spatial scales[J]. Progress in Oceanography, 2008, 79(2/4): 264-277. |
| [33] |
BAKUN A. Active opportunist species as potential diagnostic markers for comparative tracking of complex marine ecosystem responses to global trends[J]. ICES Journal of Marine Science, 2014, 71(8): 2281-2292. DOI:10.1093/icesjms/fst242 |
| [34] |
陈芃, 陈新军, 雷林. 秘鲁上升流对秘鲁鳀渔场的影响[J]. 水产学报, 2018, 42(9): 1367-1377. CHEN P, CHEN X J, LEI L. Influence of Peruvian upwelling on the Anchoveta (Engraulis ringens) fishing ground[J]. Journal of Fisheries of China, 2018, 42(9): 1367-1377. |
| [35] |
QUINOÑES J, CHIAVERANO L M, AYÑN P, et al. Spatial patterns of large jellyfish Chrysaora plocamia blooms in the Northern Humboldt Upwelling System in relation to biological drivers and climate[J]. ICES Journal of Marine Science, 2018, 75(4): 1405-1415. DOI:10.1093/icesjms/fsy004 |
| [36] |
MOLINA-VALDIVIA V, LANDAETA M F, CASTILLO M I, et al. Short-term variations in the early life history traits of common sardine Strangomera bentincki and anchoveta Engraulis ringens off central Chile[J]. Fisheries Research, 2020, 224: 105460. DOI:10.1016/j.fishres.2019.105460 |
| [37] |
KUG J S, HAM Y G. Are there two types of La Nina?[J]. Geophysical Research Letters, 2011, 38(16): L16704. |
| [38] |
陈芃, 陈新军. 表层水温结构变化对东南太平洋秘鲁鳀渔场的影响[J]. 海洋学报, 2017, 39(4): 79-88. CHEN P, CHEN X J. Evaluating the effects of sea surface temperature variations on fishing ground of anchoveta (Engraulis ringens) in the Southeast Pacific Ocean[J]. Haiyang Xuebao, 2017, 39(4): 79-88. DOI:10.3969/j.issn.0253-4193.2017.04.008 |
2. Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai 201306, China;
3. National Engineering Research Center for Oceanic Fisheries, Shanghai 201306, China;
4. Key Laboratory of Oceanic Fisheries Explotation, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China