 2022,
Vol. 31
2022,
Vol. 31



2. 上海海洋大学 海洋科学学院, 上海 201306;
3. 山东省远洋渔业协会, 山东 济南 250000
大青鲨(Prionace glauca)广泛分布于太平洋、大西洋和印度洋的热带及温带海域,是海洋中的顶级捕食者, 其在大洋中上层生态系统中扮演重要角色[1-4]。由于大量的兼捕和捕捞等因素所致, 目前大青鲨已被世界自然保护联盟(International Union for Conservation of Nature, IUCN)列为近危物种。作为金枪鱼延绳钓渔业的中最常见的兼捕鲨鱼, 大青鲨已被金枪鱼区域渔业管理组织列为重点管理和养护物种, 但由于数据的限制, 其兼捕总量的不确定性仍是当前大青鲨资源评估和管理的焦点问题之一[5-6]。
目前, 大青鲨兼捕量信息主要来源于科学观察员计划。但由于成本较高, 很少有渔业能够实现100%的观察员覆盖率。因此, 在兼捕量估算时, 常假设渔业中被观测部分的兼捕模式能够代表未被观测部分, 利用比率估计方法(ratio estimators)估算未被观测部分的兼捕量。在兼捕率存在一定的层次差异(如季节、作业深度、经度或纬度等)的情况下, 则可采用分层估计的方法, 将比率估计法单独应用于每个层次。这种分层比率估计(stratified ratio estimators)已被广泛应用到世界多个渔业的兼捕渔获量估计中[7-11]。但是, 比率估计法隐含地假设兼捕量与捕捞努力量之间存在简单线性关系[12],大多兼捕渔获量受时间、空间、环境和作业方式影响, 与捕捞努力量之间线性关系不显著, 尤其是当兼捕物种的渔获量出现大量零值时, 采用比率估计量估算时往往会导致结果偏差较大[13]。而基于模型的估算方法能够综合考虑多个因素对兼捕量的影响, 从而提高估计值的准确性[13-14]。
广义加性模型(generalized additive model, GAM)采用平滑函数研究高维数据中响应变量与解释变量之间的非线性关系, 在渔业中常被用来分析渔业资源数量分布与环境因子之间的关系, 以及多个因子的重要程度[15]。Tweedie类分布较适合拟合解释变量零值较多的情况, 能够更好地拟合响应变量和解释变量之间的关系[13, 16-17]。因此, 鉴于金枪鱼延绳钓渔业大青鲨兼捕量零值较多的现象[18], 本研究应用基于Tweedie分布的GAM模型, 分析大青鲨兼捕率与时空因素、环境因素和作业因素之间的关系, 筛选出影响大青鲨兼捕量的关键因子进行建模, 并探讨基于模型的兼捕量估计方法是否优于基于调查设计的估计方法(分层比率估计), 以期为大青鲨资源评估与管理提供更准确的兼捕量信息。
1 材料与方法 1.1 数据来源 1.1.1 作业数据作业数据来源于中国金枪鱼延绳钓渔业观察员计划(2012—2018年), 观察期间, 要求观察员记录关于渔船作业的基本信息, 包括作业位置(经纬度)、时间(具体到天)、下钩数量和下钩开始时间等信息; 同时还包括渔获种类、数量等渔获信息。共统计2785钩次, 覆盖海域为20°S~20°N, 150°E~170°W(图 1)。

|
图 1 2012—2018年中西太平洋金枪鱼延绳钓渔业观测钩次分布 Fig. 1 Observed sets distribution of tuna longline fisheries in the Western and Central Pacific Ocean from 2012 to 2018 |
大青鲨主要生活在中上层, 因此采用表层环境因子来分析。海洋环境数据来源于NOAA Ocean Watch Central Pacific Node (https://oceanwatch.pifsc.noaa.gov/doc.html)。其中, 海表面温度(sea surface temperature, FSST, 单位: ℃)以天为周期, 空间分辨率为5km; 叶绿素a质量浓度(chlorophyll, FChl.a, 单位: mg/m3)数据以月为周期, 空间分辨率为4km; 海平面距平值(sea level anomaly, FSLA, 单位: m)以天为周期, 空间分辨率为20km。选择与每钩次时空位置最接近的点的环境数据作为该钩次作业时所在海域的环境特征。
1.2 分析方法 1.2.1 影响因子分析根据大青鲨兼捕零值较多的特征, 采用Tweedie-GAM模型对大青鲨单钩次兼捕数量(bycatch, B)与影响因子之间的关系进行建模[16-17]。初步选择的影响因子包括时空因素: 经度(Flon)、纬度(Flat)、季节(season, Fseason)和月相(lunar, Flunar), 月光亮度随月相变化而变化, 在月初和月末亮度较低, 满月日亮度最大[19], 因此根据其亮度可分为3个阶段: 农历1—5日和26—30日为L1, 6—10日和21—25日为L2, 11—20日为L3;环境因素: FSST、FChl.a和FSLA[20-22]; 作业因素: 选择被要求记录在渔捞日志中的3个因素, 包括下钩开始时间(Ftime)、下钩数量(Fhooks)和单筐钩数(hooks between floats, FHBF, 常被用于指示作业深度)。
初始公式构建如下所示:
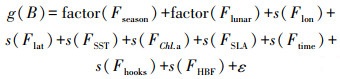 (1)
(1)
式中: g为关联函数;s为自然立方样条平滑;ε为误差项。
Tweedie-GAM中能效参数p采用Tweedie软件包中的tweedie.profile函数估算, 确定大青鲨的单钩次兼捕量服从Tweedie类的分布类型。
利用方差膨胀因子(variance inflation factor, VIF)对解释变量进行筛选, 一般认为VIF>4即存在多重共线性[17]。利用逐步回归法进行因子筛选, 最适模型选择基于赤池信息准则(akaike information criterion, AIC)、利用F检验评估因子的显著性[23]。
1.2.2 兼捕量预测效果分析受试者工作特征(receiver operating characteristic, ROC)曲线由横坐标的特异性和纵坐标的敏感性组成。ROC曲线与横坐标之间的面积为曲线下面积(area under the curve, AUC), 可以用来评价模型的整体性能和准确性。AUC值为[0.5, 0.6)表示模型预测效果不佳, [0.6, 0.7)表示效果较差, [0.7, 0.8)表示效果中等, [0.8, 0.9)表示效果较好, [0.9, 1]表示效果较好[24]。AUC值越接近1, 影响因素与预测的单钩次大青鲨兼捕量的相关性越大。
同时利用传统的分层比率估计法对大青鲨兼捕量进行估算, 以进行比较分析, 评价基于最优模型的兼捕量预测效果[13]。在金枪鱼延绳钓渔业中由于渔船时空分布特征和作业方式的差异, 季节和不同目标物种的船队[长鳍金枪鱼(Thunnus alalunga)船队和大眼金枪鱼(Thunnus obesus)船队]之间的渔获组成往往存在较大的差异[18]。因此, 按照季度和船队进行分层估算大青鲨兼捕量(表 1)。计算公式如下[13, 25]:
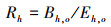 (2)
(2)
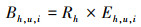 (3)
(3)

|
表 1 大青鲨兼捕量估算分层依据及各层次观测钩次数量 Tab.1 Stratified basis for estimating blue shark bycatch and the number of observed sets for each stratum |
式中: h为第h层次; o为被观测部分; u为未被观测部分; i为第i个钩次; R为兼捕率, 尾/千钩; B为兼捕量, 尾; E为捕捞努力量即下钩数量, 千钩。
预测效果分析采用交叉验证的方法, 交叉验证次数为200次, 每次随机选取80%的钩次作为观测数据, 然后利用比率估计法和建模的方法估算剩余20%的钩次的兼捕量, 最后将预测结果与观测结果进行比较。选择均方根误差(root mean square error, RMSE)比较两种估计方法之间的差异。RMSE越接近0, 则表明估计值和观测值之间的差异越小, 即表明估计效果越好[26-27]。RMSE计算公式:
 (4)
(4)
式中: n为样本数量; Bp, i为第i个估计值, Bo, i为第i个观测值。
所有的分析都在R-3.5.3语言中进行。
2 结果 2.1 因子筛选和最优模型的选择通过VIF度量对选取的初始因子进行多重共线性检验(表 2)。结果显示, 大青鲨兼捕量与10个候选变量的VIF值均小于4。故表 2中的因子均可作为初始变量。通过计算得出大青鲨兼捕量的Tweedie分布中能效参数P为1.39, 1 < P < 2, 服从复合泊松分布。

|
表 2 影响因子的多重共线性检验 Tab.2 Test of multicollinearity of factors |
利用AIC最小原则, 通过逐步回归法, 筛选后最佳Tweedie-GAM模型及各影响因子的相关参数如表 3所示。最优模型包含除季节和海平面距平值之外的所有因子, 累积偏差解释率为51.468%, 其中偏差解释率最大的因子为单筐钩数(38.238%), 其次是空间因素纬度(4.313%)和经度(3.460%)。

|
表 3 GAM模型拟合结果的偏差分析 Tab.3 Summary analysis of deviance for generalized additive models |
各影响因子对中西太平洋延绳钓渔业单钩次大青鲨兼捕量的影响如图 2所示。

|
图 2 不同因子对中西太平洋金枪鱼延绳钓渔业大青鲨单钩次兼捕量的影响 Fig. 2 Effects of different factors on the bycatch per set of blue shark in the Western and Central Pacific tuna longline fishery |
时空分析结果显示, 单钩次大青鲨兼捕量的季节差异不显著, 但受月相影响(解释率为0.163%), L3阶段兼捕量显著高于L2和L1阶段; 在纬度上, 单钩次兼捕量在低纬度(10°S~5°N)海域差异随纬度变化较小, 随后呈现出纬向递增的趋势; 在经度上, 总体呈现出东高西低的趋势(图 2)。
环境因子中影响最大的是海表面温度, 在25.0~29.5℃时, 单钩次兼捕量随着海表温的升高而降低, 之后(>29.5℃)随着温度的升高而降低; 其次是叶绿素质量浓度, 在质量浓度较高的海域单钩次兼捕量相对较高; 海平面距平值对单钩次兼捕量的影响不显著(图 2)。
作业因素影响分析表明单钩次兼捕量随着单筐钩数的增加呈现波动降低的趋势, 单筐钩数为15~19时, 兼捕量较高, 且相对变化较小, 为20~24时显著下降, 为25~35时呈现先上升后下降的趋势; 下钩数量与单钩次兼捕量总体线性关系不显著, 在单钩次钩数较少时(1000~2000)线性增加的趋势较为显著, 随着单钩次下钩数量的增加(2000~3000), 兼捕量的增加趋势逐渐降低, 当单钩次下钩数量超过3000之后, 对兼捕量的影响不显著。中西太平洋金枪鱼延绳钓渔业下钩开始时间为00:00~12:00, 其中多数钩次集中在02:00~10:00开始下钩, 模型分析结果显示, 下钩开始时间对单钩次兼捕量的影响并不显著(表 3, 图 2)。
2.3 预测效果ROC分析结果显示, AUC值大于0.7(0.749), 表明模型的预测效果中等。
交叉验证结果显示, 分层比率估计法和基于Tweedie-GAM的大青鲨兼捕量估算方法的RMSE均值分别为2.102和1.501(图 3)。表明基于最优Tweedie-GAM模型的太平洋金枪鱼延绳钓渔业大青鲨兼捕量估算准确度和精确度高于分层比率估计法。

|
图中数字为RMSE均值。 The values in the plot represent mean RMSE. 图 3 不同兼捕估计方法的均方根误差比较 Fig. 3 Comparison of RMSE of different bycatch estimation methods |
延绳钓是一种在一根干线上系结许多等距离的支线, 末端结有钓钩和饵料, 利用浮、沉子装置, 将其敷设于表、中和底层的作业方式。该种捕捞模式表明捕捞能力很大程度上取决于下钩深度与资源垂直分布的重叠程度, 但受信息可获得性限制, 渔业研究者通常用HBF近似地代表最大作业深度(一般认为, HBF越少, 最大深度相对越浅)[28-29]。目前HBF仍是渔捞日志中唯一与作业深度有关的因素, 也是依据观察员数据估算大青鲨总兼捕量时的重要依据[30]。因此本研究选择HBF近似地代表不同钩次之间作业深度的差异。相对于目标物种, 鲨鱼分布水层较浅, HBF越少, 兼捕到的鲨鱼渔获率越高[20-22, 31-32]。本研究中HBF作为影响大青鲨单钩兼捕量的主要因素, 也表达了类似的特征, 尤其是增加HBF(≥20), 其兼捕量显著下降。
月相是影响许多物种渔获率的重要因素, 饵料生物具有昼夜移动的习性, 月光穿透的水层影响饵料生物所处水深和密度[33]。本研究发现, 当船只在满月日(L3)作业时大青鲨单钩次兼捕量相对较高, 与马绍尔群岛以及在澳大利亚新南威尔士州北部的延绳钓渔业的大青鲨兼捕模式一致[21, 34]。大青鲨在满月日的夜间平均栖息深度更高, 主要与猎物在月光较强的夜间下潜深度更深有关[35]。为了提高渔获效率同时降低鲨鱼的兼捕, 以金枪鱼为目标物种的延绳钓渔业作业深度相对较深[33], 满月日大青鲨随饵料分布而下潜, 增加了遭遇渔具的概率[21, 34]。但POISSON等对留尼旺岛以及BIGELOW等对北太平洋的剑鱼延绳钓渔业中大青鲨的兼捕研究认为, 大青鲨是机会主义捕食者, 与月相没有相关关系[19-20]。这种差异可能是由于剑鱼渔业作业深度相对较浅有关, 鲨鱼随着月相变化潜浮对兼捕率影响可能不显著。
3.2 影响兼捕量的空间和环境特征WORM等[36-37]研究认为海洋捕食者多样性热点区域多集中在中纬度地区(20°N~30°N和20°S~30°S), 与初级生产力和浮游动物的纬向分布特征显著相关, 同时, 中纬度海域也是热带和温带物种分布范围重叠的区域, 其饵料资源丰富, 吸引捕食者在此聚集。全球鲨鱼的分布也在中纬度地区物种丰富度最高[38]。本研究中大青鲨兼捕量的纬向分布特征验证了中西太平洋水域大青鲨资源分布的空间异质性, 且以中纬度地区较高[20-22]。兼捕量的季节特征以及在10°S~5°N海域之间纬向分布特征, 进一步反映了在该水域范围内大青鲨资源时空分布相对比较均匀[39]。除此之外, 研究区域东部(178°E~170°W)较高的兼捕量表明在热带水域大青鲨资源分布存在一定的经向梯度。这种分布模式可能与研究区域内岛屿和海底火山分布的东西差异有关。有研究[40]发现, 延绳钓6渔业中多数物种的渔获率都与离岸距离和离海底火山距离显著相关, 主要受不同地理区域的饵料丰富程度和活动空间大小影响, BROMHEAD等[21]研究发现在远离海底火山的水域,大青鲨兼捕率相对较高, 相较于西部海底火山和岛屿遍布的水域, 研究区域内东部水域则更加开阔。
水温是影响鱼类生存、生长和繁殖的重要因子之一。大青鲨虽然分布广泛, 但其95%的生活史阶段栖息于水温为9.7~26.9℃的海域[41]。这也是导致相较于温带水域, 大青鲨在热带海域的栖息水层更深的主要因素[39]。较高的海表温可能会导致大青鲨向较深的适温水层垂直迁移, 因此增加了与钓钩的遭遇率, 而过高的海表温可能不适宜大青鲨栖息或导致大青鲨下潜至更深的水层。
3.3 基于最优模型的兼捕量预测效果STOCK等[13-14]对多个渔业中兼捕物种渔获量估算方法比较分析中发现, 绝大多数兼捕渔获量与捕捞努力量呈现非线性关系或较弱的线性关系, 在只考虑时空因素和作业因素(作业深度和渔具类型等)的情况下, 基于模型的估算方法(如GAM和随机森林等)都要优于分层比率估计方法。本研究中, 进一步验证了金枪鱼延绳钓渔业中大青鲨兼捕量与每钩次下钩数量的非线性关系, 这可能导致利用基于线性假设的比率估计法获得的估计值将存在较大的误差。月相、作业深度、海表面温度等环境因素对大青鲨兼捕量的显著影响以及不规则的时空分布特征[18]也进一步表明简略的时空分层并不能有效考虑大青鲨兼捕量在不同层次之间的差异。因此, 本研究建议后续对兼捕总量的估计均采用模型估算方法。
3.4 总结与展望综上所述, 基于Tweedie-GAM分析的延绳钓渔业对大青鲨兼捕量的影响因素得出的结果与相关研究得出的大青鲨的生境、生物学和行为的研究结果基本一致。为降低大青鲨兼捕, 建议中西太平洋延绳钓渔业单筐钩数不低于20, 应控制在兼捕量较高的中纬度以及178°E以东水域作业的渔船数量, 并重点关注其兼捕减缓措施的执行情况。虽然本研究显示基于模型的方法能够获得更准确的估计值, 但模型解释率和AUC结果表明基于当前渔捞日志中可获得信息的模型仍有较大的提升空间。因此, 将更多的与兼捕率相关的因素(如饵料类型、渔具类型[42]以及与实际作业深度有关的信息)纳入渔捞日志记录要求中, 为分层比率估计或建模估计提供更多依据, 可能是未来除了加强监测力度[9]之外降低兼捕量估计不确定性的另一重要途径。
| [1] |
朱江峰, 戴小杰, 陈彦. 运用生产力-易捕率指数对10种热带太平洋鲨鱼种群的研究[J]. 南方水产科学, 2013, 9(6): 8-13. ZHU J F, DAI X J, CHEN Y. Productivity-susceptibility analysis of 10 shark populations in tropical Pacific Ocean[J]. South China Fisheries Science, 2013, 9(6): 8-13. DOI:10.3969/j.issn.2095-0780.2013.06.002 |
| [2] |
高春霞, 戴小杰, 吴峰, 等. 北大西洋大青鲨年龄和生长初步研究[J]. 上海海洋大学学报, 2013, 22(1): 120-127. GAO C X, DAI X J, WU F, et al. A preliminary study on age and growth of the blue shark (Prionace glauca)in the North Atlantic Ocean[J]. Journal of Shanghai Ocean University, 2013, 22(1): 120-127. |
| [3] |
高春霞, 戴小杰, 田思泉, 等. 北大西洋大青鲨种群统计分析[J]. 应用生态学报, 2016, 27(2): 622-628. GAO C X, DAI X J, TIAN S Q, et al. Demographic analysis of the blue shark, Prionace glauca, in the North Atlantic Ocean[J]. Chinese Journal of Applied Ecology, 2016, 27(2): 622-628. |
| [4] |
李云凯, 高小迪, 王琳禹, 等. 东太平洋中部中上层鲨鱼群落营养生态位分化[J]. 应用生态学报, 2018, 29(1): 309-313. LI Y K, GAO X D, WANG L Y, et al. Trophic niche partitioning of pelagic sharks in Central Eastern Pacific inferred from stable isotope analysis[J]. Chinese Journal of Applied Ecology, 2018, 29(1): 309-313. |
| [5] |
耿喆, 朱江峰, 夏萌, 等. 运用数据缺乏方法估算印度洋大青鲨可持续渔获量[J]. 中国水产科学, 2017, 24(5): 1099-1106. GENG Z, ZHU J F, XIA M, et al. Estimate of sustainable yield of blue shark (Prionace glauca) in the Indian Ocean using data-poor approach[J]. Journal of Fishery Sciences of China, 2017, 24(5): 1099-1106. |
| [6] |
CLARKE S C, SATO M, SMALL C, et al. Bycatch in longline fisheries for tuna and tuna-like species: a global review of status and mitigation measures[R]. FAO Fisheries and Aquaculture Technical Paper 588. Rome: FAO, 2014: 1-199.
|
| [7] |
AMANDÈ M J, ARIZ J, CHASSOT E, et al. Bycatch of the European purse seine tuna fishery in the Atlantic Ocean for the 2003-2007 period[J]. Aquatic Living Resources, 2010, 23(4): 353-362. DOI:10.1051/alr/2011003 |
| [8] |
AMANDÈ M J, CHASSOT E, CHAVANCE P, et al. Precision in bycatch estimates: the case of tuna purse-seine fisheries in the Indian Ocean[J]. ICES Journal of Marine Science, 2012, 69(8): 1501-1510. DOI:10.1093/icesjms/fss106 |
| [9] |
BABCOCK E A, PIKITCH E K, HUDSON C G. How much observer coverage is enough to adequately estimate bycatch?[M]. Washington, DC: Pew Institute for Ocean Science, 2003.
|
| [10] |
HUANG H W. Incidental catch of seabirds and sea turtles by Taiwanese longline fleets in the Pacific Ocean[J]. Fisheries Research, 2015, 170: 179-189. DOI:10.1016/j.fishres.2015.06.004 |
| [11] |
HUANG H W, LIU K M. Bycatch and discards by Taiwanese large-scale tuna longline fleets in the Indian Ocean[J]. Fisheries Research, 2010, 106(3): 261-270. DOI:10.1016/j.fishres.2010.08.005 |
| [12] |
FONTENEAU A, RICHARD N. Relationship between catch, effort, CPUE and local abundance for non-target species, such as billfishes, caught by Indian Ocean longline fisheries[J]. Marine and Freshwater Research, 2003, 54(4): 383-392. DOI:10.1071/MF01268 |
| [13] |
STOCK B C, WARD E J, THORSON J T, et al. The utility of spatial model-based estimators of unobserved bycatch[J]. ICES Journal of Marine Science, 2019, 76(1): 255-267. DOI:10.1093/icesjms/fsy153 |
| [14] |
STOCK B C, WARD E J, EGUCHI T, et al. Comparing predictions of fisheries bycatch using multiple spatiotemporal species distribution model frameworks[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2020, 77(1): 146-163. DOI:10.1139/cjfas-2018-0281 |
| [15] |
HASTIE T J, TIBSHIRANI R J. Generalized additive models[M]. London: Chapman & Hall, 1990.
|
| [16] |
王浩展, 戴小杰, 官文江, 等. 基于Tweedie-GAM模型的热带大西洋拟锥齿鲨单位捕捞努力量渔获量与环境因子的关系[J]. 应用生态学报, 2017, 28(6): 2024-2032. WANG H Z, DAI X J, GUAN W J, et al. Relationship between the environmental factors and the CPUE (catch per unit effort) of Pseudocarcharias kamoharai in tropical Atlantic Ocean based on the GAM-Tweedie model[J]. Chinese Journal of Applied Ecology, 2017, 28(6): 2024-2032. |
| [17] |
张云雷, 徐宾铎, 张崇良, 等. 基于Tweedie-GAM模型研究海州湾小黄鱼资源丰度与栖息环境的关系[J]. 海洋学报, 2019, 41(12): 78-89. ZHANG Y L, XU B D, ZHANG C L, et al. Relationship between the habitat factors and the abundance of small yellow croaker (Larimichthys polyactis) in Haizhou Bay based on the Tweedie-GAM model[J]. Haiyang Xuebao, 2019, 41(12): 78-89. DOI:10.3969/j.issn.0253-4193.2019.12.008 |
| [18] |
WANG J Q, GAO C X, WU F, et al. The discards and bycatch of Chinese tuna longline fleets in the Pacific Ocean from 2010 to 2018[J]. Biological Conservation, 2021, 255: 109011. DOI:10.1016/j.biocon.2021.109011 |
| [19] |
POISSON F, GAERTNER J C, TAQUET M, et al. Effects of lunar cycle and shing operations on longline-caught pelagic fish: fishing performance, capture time, and survival of fish[J]. Fishery Bulletin, 2010, 108(3): 268-281. |
| [20] |
BIGELOW K A, BOGGS C H, HE X. Environmental effects on swordfish and blue shark catch rates in the US North Pacific longline fishery[J]. Fisheries Oceanography, 1999, 8(3): 178-198. DOI:10.1046/j.1365-2419.1999.00105.x |
| [21] |
BROMHEAD D, CLARKE S, HOYLE S, et al. Identification of factors influencing shark catch and mortality in the Marshall Islands tuna longline fishery and management implications[J]. Journal of Fish Biology, 2012, 80(5): 1870-1894. DOI:10.1111/j.1095-8649.2012.03238.x |
| [22] |
WALSH W A, KLEIBER P. Generalized additive model and regression tree analyses of blue shark (Prionace glauca) catch rates by the Hawaii-based commercial longline fishery[J]. Fisheries Research, 2001, 53(2): 115-131. DOI:10.1016/S0165-7836(00)00306-4 |
| [23] |
徐国强, 朱文斌, 张洪亮, 等. 基于GAM模型分析印度洋大眼金枪鱼和黄鳍金枪鱼渔场分布与不同环境因子关系[J]. 海洋学报, 2018, 40(12): 68-80. XU G Q, ZHU W B, ZHANG H L, et al. Relationship between fishing grounds of Thunnus obesus and Thunnus albacores with environmental factors in the Indian Ocean based on generalized additive model[J]. Haiyang Xuebao, 2018, 40(12): 68-80. DOI:10.3969/j.issn.0253-4193.2018.12.008 |
| [24] |
SWETS J A. Measuring the accuracy of diagnostic systems[J]. Science, 1988, 240(4857): 1285-1293. DOI:10.1126/science.3287615 |
| [25] |
COCHRAN W G. Sampling techniques[M]. 3rded. New York: John Wiley & Sons, 1977.
|
| [26] |
戴黎斌, 陈锦辉, 田思泉, 等. 基于CART算法的长江口鱼种丰富度预测[J]. 中国水产科学, 2018, 25(5): 1082-1090. DAI L B, CHEN J H, TIAN S Q, et al. Prediction of fish species richness in the Yangtze River estuary using CART algorithm[J]. Journal of Fishery Sciences of China, 2018, 25(5): 1082-1090. |
| [27] |
LYNCH D R, MCGILLICUDDY D J JR, WERNER F E. Skill assessment for coupled biological/physical models of marine systems[J]. Journal of Marine Systems, 2009, 76(1/2): 1-3. |
| [28] |
BIGELOW K, MUSYL M K, POISSON F, et al. Pelagic longline gear depth and shoaling[J]. Fisheries Research, 2006, 77(2): 173-183. DOI:10.1016/j.fishres.2005.10.010 |
| [29] |
WARD P, MYERS R A. Inferring the depth distribution of catchability for pelagic fishes and correcting for variations in the depth of longline fishing gear[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2005, 62(5): 1130-1142. DOI:10.1139/f05-021 |
| [30] |
PEATMAN T, BELL T, ALLAIN V, et al. Summary of longline fishery bycatch at a regional scale, 2003-2017[C]//Proceedings of the 14th Regular Session of the Western and Central Pacific Fisheries Commission Scientific Committee. Busan, Republic of Korea: WCPFC, 2018.
|
| [31] |
张艳波, 戴小杰, 朱江峰, 等. 东南太平洋金枪鱼延绳钓主要渔获种类垂直分布[J]. 应用生态学报, 2015, 26(3): 912-918. ZHANG Y B, DAI X J, ZHU J F, et al. Vertical distribution of main species captured by tuna longline fishery in the southeast Pacific Ocean[J]. Chinese Journal of Applied Ecology, 2015, 26(3): 912-918. |
| [32] |
GILMAN E, CLARKE S, BROTHERS N, et al. Shark interactions in pelagic longline fisheries[J]. Marine Policy, 2008, 32(1): 1-18. DOI:10.1016/j.marpol.2007.05.001 |
| [33] |
宋利明, 张智. 月相和延绳钓不同投绳时间对大眼金枪鱼和黄鳍金枪鱼渔获率的影响[J]. 大连海洋大学学报, 2010, 25(3): 214-218. SONG L M, ZHANG Z. Effects of lunar phases and setting time on the catch rates of bigeye tuna and yellowfin tuna[J]. Journal of Dalian Ocean University, 2010, 25(3): 214-218. DOI:10.3969/j.issn.1000-9957.2010.03.005 |
| [34] |
LOWRY M, WILLIAMS D, METTI Y. Lunar landings-Relationship between lunar phase and catch rates for an Australian gamefish-tournament fishery[J]. Fisheries Research, 2007, 88(1/3): 15-23. |
| [35] |
CAMPANA S E, DOREY A, FOWLER M, et al. Migration pathways, behavioural thermoregulation and overwintering grounds of blue sharks in the Northwest Atlantic[J]. PLoS One, 2011, 6(2): e16854. DOI:10.1371/journal.pone.0016854 |
| [36] |
WORM B, LOTZE H K, MYERS R A. Predator diversity hotspots in the blue ocean[J]. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(17): 9884-9888. DOI:10.1073/pnas.1333941100 |
| [37] |
WORM B, SANDOW M, OSCHLIES A, et al. Global patterns of predator diversity in the open oceans[J]. Science, 2005, 309(5739): 1365-1369. DOI:10.1126/science.1113399 |
| [38] |
LUCIFORA L O, GARCÍA V B, WORM B. Global diversity hotspots and conservation priorities for sharks[J]. PLoS One, 2011, 6(5): e19356. DOI:10.1371/journal.pone.0019356 |
| [39] |
COMPAGNO L J V. FAO species catalogue. Vol. 4. Sharks of the world. An annotated and illustrated catalogue of shark species known to date. Part 1. Hexanchiformes to Lamniformes[J]. FAO Fishery Synopsis 125, 1984, 4: 1-249. |
| [40] |
GILMAN E, CHALOUPKA M, READ A, et al. Hawaii longline tuna fishery temporal trends in standardized catch rates and length distributions and effects on pelagic and seamount ecosystems[J]. Aquatic Conservation: Marine and Freshwater Ecosystems, 2012, 22(4): 446-488. DOI:10.1002/aqc.2237 |
| [41] |
MUSYL M K, BRILL R W, CURRAN D S, et al. Postrelease survival, vertical and horizontal movements, and thermal habitats of five species of pelagic sharks in the central Pacific Ocean[J]. Fishery Bulletin, 2011, 109(4): 341-368. |
| [42] |
GILMAN E, CHALOUPKA M, MUSYL M. Effects of pelagic longline hook size on species- and size-selectivity and survival[J]. Reviews in Fish Biology and Fisheries, 2018, 28(2): 417-433. DOI:10.1007/s11160-017-9509-7 |
2. College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China;
3. Shandong Overseas Fisheries Associatic, Jinan 250000, Shandong, China