 2022,
Vol. 31
2022,
Vol. 31



2. 国家远洋渔业工程技术研究中心, 上海 201306;
3. 大洋渔业资源可持续开发教育部重点实验室, 上海 201306;
4. 农业农村部大洋渔业开发重点实验室, 上海 201306;
5. 农业农村部大洋渔业资源环境科学观测实验站, 上海 201306
海洋是地球重要组成部分, 占地球表面面积的71%, 海洋渔业为2.6亿人类创造了工作机会[1], 同时也是人类重要的优质动物蛋白来源[2]。联合国粮农组织(Food and Agriculture Organization, FAO)将全球海域划分为19个主要渔区[3], 据FAO渔业统计, 1950—2019年累计渔获量最高的是西北太平洋渔区(FAO 61区), 平均年产量高达1678.3万t, 最低的为北冰洋渔区(FAO 18区), 平均年产量仅为217.5t;渔获量最高的类别为中上层鱼类, 达1291.54万t, 最低的为其他海洋动物, 年产量仅为16.66万t。不同海域间存在着生态系统、渔业资源量、鱼种类型等差异[4], 导致了各海域间渔获量的不同。有研究[5]表明, 不同海域海洋渔获物数量和种类组成受海洋温度变化影响显著, 同时又由于过度捕捞[6]使全球渔获量比半个世纪前下降了一半, 这进一步加剧了各渔区渔获量的差别。FAO报告显示, 截止到2017年有34.2%的海洋渔业处于过度捕捞状态[2]。有学者已对西北太平洋[7]、东南大西洋[8]、印度洋[9]等海域进行了渔获量和主要鱼种的分析研究, 但这些研究都仅限于单一渔区或海域, 缺少了大尺度视角的分析。因此, 为了更加合理利用开发全球海洋渔业资源, 有必要从全球海域角度, 探讨分析各渔区渔获量及主要鱼种渔获量的相互关系, 这对人类保护管理海洋渔业资源具有重要意义。
灰色理论系统是由我国学者邓聚龙教授创立的一门不确定系统理论, 其主要特点是能对已知的“少数据”和“贫信息”进行数据挖掘, 从而实现对整个系统行为和演变的正确描述和有效监控[10]。自创立以来, 该方法在航空航天、军事应用、地质勘探、企业金融、渔业产业等各个行业和领域均得到了广泛应用[10-13]。海洋渔获量具有受影响因素多、数据信息少等明显特点[13], 因此本文拟利用灰色关联聚类分析法, 研究全球各渔区渔获量与总渔获量之间的关系, 并确定同类型渔区中影响渔获量大小的主要鱼种类型, 为全球海洋渔业可持续发展提供参考。
1 材料与方法 1.1 数据来源据FAO统计, 全球累计渔获量自1950年起开始了持续增长模式, 但进入20世纪90年代后, 渔获量保持了稳定变动趋势, 为此本研究数据的时间范围选定为1990—2019年。全球海域渔获量数据来源于FAO网站(http://www.fao.org/fishery/statistics/global-production/zh), 运用FishStatJ软件按照FAOSTAT的格式下载获得, 包括19个渔区1990—2019年近30年的渔获量数据。其中渔获量按中上层鱼类、底层鱼类、软体动物(除头足类)、甲壳类、其他海洋鱼类、头足类、其他海洋动物、洄游性鱼类、水生植物等分为9大类。
1.2 研究方法 1.2.1 灰色关联分析法设Xm为母序列, Xm=(xm(1), xm(2), …, xm(n)); 设Xz为子序列, Xz=(xz(1), xz(2), …, xz(n)), 进行灰色关联分析, 可获得影响全球海域总渔获量的主要渔区类别或影响某一渔区总渔获量的主要鱼种类别。灰色关联系数计算公式[10]:
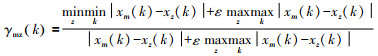 (1)
(1)
式中: ε为分辨系数; γmz(k)为母序列Xm与子序列Xz的灰色关联系数; xm(k)为母序列, xz(k)为子序列; k为年份; z为19大渔区或9大类渔获量种类。关联度计算:
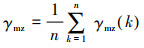 (2)
(2)
式中: γmz为母序列与子序列z的关联度; n为年份, 从1990年到2019年, 其值为30。在灰色关联计算中, 采用均值化进行初值化, 分辨系数取0.5。
1.2.2 灰色聚类分析法设有n个观测对象, 每个对象观测m个特征数据, 得到序列[10]如下:
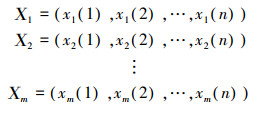 (3)
(3)
式中: 观测对象X为年份, 特征数据x为各渔区各年份的渔获量。
对所有的α≤β,α,β=1, 2,…,m, 计算出Χα与Χβ的灰色绝对关联度εαβ, 获得上三角关联聚类矩阵:
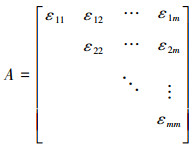 (4)
(4)
设临界值为r, 并要求r>0.5, 当εαβ≥r(α≠β)时, 则视Χα与Χβ为同类特征。根据灰色理论原理[10], 灰色关联聚类的临界值r越大, 其分类越精细, 设定各渔区间关联聚类矩阵的临界值r=0.975作为分类依据。
2 结果 2.1 全球各渔区渔获量与全球海域总渔获量的灰色关联分析根据FAO统计数据分析, 1990—2019年全球海洋渔业总渔获量为246796.29万t。全球各渔区渔获量排名前3位的渔区是西北太平洋、东南太平洋、中西太平洋, 其累计渔获量分别为63430.04万t、37218.92万t、31933.01万t, 依次占全球总渔获量的25.70%、15.08%、12.94%, 平均年渔获量分别为2114.33万t、1240.63万t、1064.43万t。渔获量排名最后3位的渔区分别是南极印度洋扇区、南极太平洋扇区、北冰洋, 其累计渔获量依次是35.60万t、6.30万t、2525t, 依次占全球总渔获量的0.014%、0.0025%、0.0001%。从各大洋分析可知, 太平洋海域渔获量最高, 达148726.00万t, 其次是大西洋海域, 渔获量为64030.81万t, 而两极的渔获量最低, 合计仅为599.12万t。
灰色关联分析表明(表 1), 全球大多数渔区渔获量与全球海域总渔获量关联度较大, 灰色关联度大于0.900的渔区有16个, 最大的为西北太平洋, 为0.988, 其次是地中海及黑海的0.983和东北太平洋的0.979;灰色关联度低于0.900的仅有3个渔区, 分别为南极大西洋扇区、南极太平洋扇区和北冰洋, 灰色关联度依次是0.893、0.838和0.739。

|
表 1 各渔区渔获量与全球海域总渔获量的灰色关联度 Tab.1 Grey correlation indices between the catch in each fishing area and the total catch of global marine area |
全球各渔区间的灰色聚类结果表明(表 2), 各渔区间的关联度均较高, 其中东印度洋和西印度洋的关联度最高, 为0.989;东北太平洋和北冰洋、中西大西洋和北冰洋, 它们的关联度最低, 均为0.737。另外, 除南极大西洋扇区、南极太平洋扇区和北冰洋外, 其余各渔区相互间的关联度均大于0.900;南极太平洋扇区与其余各渔区间的关联度均为0.820~0.859, 北冰洋与其余各渔区间的关联度均为0.737~0.782。

|
表 2 全球各渔区之间的灰色关联聚类系数 Tab.2 Grey relational clustering coefficients among fishing areas |
按设定的临界值r=0.975作为分类依据, 可获得2类聚类类群。类群Ⅰ包含6个渔区, 分别是西北太平洋、东北大西洋、东北太平洋、东南大西洋、中西大西洋、地中海及黑海; 类群Ⅱ包含4个渔区, 分别为中西太平洋、东印度洋、中东大西洋、西印度洋。其余9个渔区聚类效果不明显, 变化趋势不一致, 各自成类。
类群Ⅰ的6个渔区渔获量均在一定范围内波动(图 1a), 其渔业资源开发呈现稳定的状态, 处于充分开发阶段, 其中西北太平洋年渔获量为1964.28万~2402.09万t, 平均年渔获量为2114.33万t; 东北大西洋年渔获量为831.58万~1211.30万t, 平均年渔获量为990.03万t; 东北太平洋年渔获量为224.45万~337.72万t, 平均年渔获量为294.17万t; 中西大西洋年渔获量为111.00万~219.80万t, 平均年渔获量为158.34万t; 东南大西洋年渔获量为121.55万~196.64万t, 平均年渔获量为155.64万t; 地中海及黑海年渔获量为112.78万~170.93万t, 平均年渔获量为145.29万t。

|
图 1 1990—2019年同类群渔区渔获量变化趋势 Fig. 1 Variability in catch in homogeneous fishing areas from 1990 to 2019 |
类群Ⅱ的4个渔区年渔获量基本呈稳定增长趋势(图 1b), 其渔业资源开发处在未充分开发状态, 其中中东大西洋年渔获量在290.67万t(1994年)至547.57万t(2018年); 中西太平洋年渔获量在740.04万t(1990年)至1394.24万t(2019年); 东印度洋年渔获量在335.44万t(1990年)至697.20万t(2017年); 西印度洋年渔获量在327.26万t(1990年)至554.52万t(2018年)。
2.3 同类型渔区各大类鱼种渔获量的灰色关联分析 2.3.1 类群Ⅰ鱼种渔获量变化趋势及灰色关联分析1990—2019年, 类群Ⅰ的6大渔区总渔获量与9大类渔获量的灰色关联分析表明(表 3), 所有种类的灰色关联度均大于0.600, 小于0.900, 其中排名前两位的是中上层鱼类和底层鱼类, 其灰色关联度均大于0.800, 分别为0.892和0.852;灰色关联度最小的是头足类, 为0.601。

|
表 3 类群Ⅰ的各类别渔获量与总渔获量的灰色关联度 Tab.3 Grey correlation indices between each category of catch and total catch of Group Ⅰ |
1990—2019年, 类群Ⅰ的6大渔区中底层鱼类、中上层鱼类是主要鱼种, 其中西北太平洋、东北大西洋年平均渔获量排名前两位的鱼种为底层鱼类、中上层鱼类, 年平均渔获量分别为605.08万t、590.21万t和455.12万t、442.24万t, 占各渔区年平均渔获量的28.62%、27.91%和45.97%、44.67%;东南大西洋、地中海及黑海年平均渔获量排名前两位的鱼种为中上层鱼类、底层鱼类, 年平均渔获量分别为102.83万t、42.29万t和87.28万t、26.97万t, 占各渔区年平均渔获量的66.07%、27.17%和60.07%、18.56%;东北太平洋年平均渔获量排名前两位的鱼种为底层鱼类、洄游性鱼类, 年平均渔获量分别为232.50万t、38.12万t, 占该渔区年平均渔获量的79.04%、12.96%;中西大西洋年平均渔获量排名前两位的鱼种为中上层鱼类、甲壳类, 年平均渔获量分别为73.79万t、26.03万t, 占该渔区年平均渔获量的46.60%、16.44%。
2.3.2 类群Ⅱ鱼种渔获量变化趋势及灰色关联分析1990—2019年, 类群Ⅱ的4大渔区总渔获量与9大渔获量种类的关联分析表明(表 4), 大多数种类渔获量与总渔获量的关联度较高, 除其他海洋动物和水生植物外, 其余7个种类关联度均在0.800以上, 其中排名前五位种类的灰色关联度均大于0.900。中上层鱼类灰色关联度最大, 达0.960, 其余前五依次为底层鱼类、甲壳类、头足类、其他海洋鱼类。

|
表 4 类群Ⅱ的各渔获量种类与总渔获量的灰色关联度 Tab.4 Grey correlation indices between each category of catch and the total catch of Group Ⅱ |
1990—2019年, 类群Ⅱ的4大渔区中中上层鱼类、其他海洋鱼类、底层鱼类为主要种类, 其中中东大西洋和西印度洋年平均渔获量排名前两位的鱼种为中上层鱼类、底层鱼类, 年平均渔获量分别为294.95万t、55.91万t和199.15万t、114.70万t, 占各渔区年平均渔获量的73.09%、13.86%和46.25%、26.63%;中西太平洋年平均渔获量排名前两位的鱼种为中上层鱼类、其他海洋鱼类, 年平均渔获量分别为527.83万t、257.22万t, 占渔区年平均渔获量的49.59%、24.16%;东印度洋年平均渔获量排名前两位的鱼种是其他海洋鱼类、中上层鱼类, 年平均渔获量分别为190.19万t、168.86万t, 占渔区年平均渔获量的35.30%、31.34%。
3 讨论与分析 3.1 各渔区渔获量与全球总渔获量的关系有研究[14]表明, 全球海域通过海洋环流形成一个大整体, 海洋生物存在着连续分布, 海洋渔业捕捞的大量扩张覆盖了全球海域, 过度捕捞使全球渔获量经历了不断增长阶段后, 出现下降, 其增长受到了限制[15]。研究结果发现, 各渔区渔获量与全球总渔获量的灰色关联度均较大, 绝大多数的灰色关联度均在0.950以上, 各渔区的渔获量对总渔获量的贡献大; 灰色关联度最大的西北太平洋、地中海及黑海、东北太平洋等3个渔区, 均分布在温带区域, 其近30年的渔获量变化趋势也一致, 均稳定在一定范围内, 渔业资源被充分开发[2], 其中西北太平洋平均年渔获量是地中海及黑海、东北太平洋2个渔区年平均渔获量之和的4.8倍; 灰色关联度最小的南极大西洋扇区、南极太平洋扇区、北冰洋等3个渔区, 均分布在南北两极, 是渔获开发力度低, 渔获量较少的区域, 其中北冰洋的灰色关联度最低, 小于0.800, 其渔获量也是全球最低, 对全球渔获量的贡献度几乎可以忽略不计。
有研究指出, 全球气候变暖打破了北冰洋与北大西洋和北太平洋之间的屏障[16], 促进了不同海域间海洋生物的交流[17-18], 预计到2100年, 海洋鱼类种群间的交换将引发北大西洋和北太平洋生物多样性和食物网的变化, 其对生态系统产生的生态和经济效应会引起全球39%的渔获量的变化[18]。有学者[19-24]认为, 海水温度、海水含氧量水平、海洋热浪(MHWs)、南极海冰变化以及其他海洋环境因素的变化, 都深刻影响着海洋鱼类在各海域间的分布和交换。在后续探究不同渔区间渔获量以及各渔区渔获量和全球总渔获量关系时, 可以增加气候变化和海洋环境因素对渔场分布和资源变动影响的研究, 更好地完善研究的深度和广度。
3.2 各渔区海洋渔业开发状态评价对全球各渔区的灰色聚类分析发现, 各渔区间关联度高, 在选取0.975的临界值后, 形成了聚类明显的两类渔区类群Ⅰ和类群Ⅱ。类群Ⅰ除中西大西洋外, 主要分布在温带及亚热带向两极区域; 类群Ⅱ的渔区主要集中在热带或热带至温带区域。类群Ⅰ的6个渔区渔获量大小不同, 但均在各自一定范围内, 或达到最高峰值后波动下落, 或达到峰值后出现波动, 这与FAO报告的评估结论基本一致, 其渔业大多已充分开发[2]。造成类群Ⅰ各渔区渔获量变化趋势的因素可能主要和过度捕捞有关, 据FAO报告指出, 由于过度捕捞, 2017年, 地中海及黑海渔区生物可持续捕捞率为37.5%, 为全球所有渔区中最低[2]; 西北太平洋[7, 25]、东南大西洋[26-28]、东北大西洋[29]均因历史上主要鱼种被过度捕捞, 而影响了渔获量。各渔区实施严格的渔业管理措施, 大幅降低了东北大西洋的渔船数量[30], 加强了东南大西洋的资源养护[31]和配额管理[32], 限制了东北太平洋部分区域年度捕捞量[33]和主要鱼种的管理[34], 恢复了中西大西洋主要鱼种资源[2], 使得4个渔区的生物资源量得到了一定恢复, 其2017年可持续捕捞水平分别为79.3%、67.6%、83.9%、61.4%[2]; 另外, 各海区复杂的海洋环境, 影响了渔区渔业资源量分布和捕捞作业水平[23, 35], 也一定程度上影响了渔区的渔获量
类群Ⅱ的4个渔区渔获量大小不一, 但均呈现一致的增长趋势, 其渔业还具有一定开发潜力, 其中, 东印度洋、西印度洋、中西太平洋3个渔区的评估, 与FAO报告中持续上升姿态的评估结果一致[2]。有研究表明, 东印度洋和西印度洋渔区的岛屿众多, 寒暖流交汇、上升流机制形成了良好的渔场环境, 孕育了丰富的渔业资源[36-37], 同时合理的开发[36, 38]和管理[36]保证了渔获量的增长状态; 中西太平洋渔区岛国众多, 渔业资源充足, 是全球最大的金枪鱼产区[39], 因捕捞压力强度大[40], 以及3个区域组织[39]和各国的管理措施[41]保障了渔获量的增长趋势; 中东大西洋渔区寒暖流交汇及季节性上升流形成了良好的渔场, 在高强度捕捞和缺少科学管理情况下, 渔获量也保持了持续增加[42]。
研究发现, 类群Ⅰ和类群Ⅱ渔区的几大渔获物中, 与渔区总渔获量关联度最高的前两位鱼种均为中上层鱼类和底层鱼类, 同时FAO各渔区渔获量统计也表明, 该两种鱼种也是各渔区的主要鱼种, 其中有6个渔区的年平均渔获量的前两位是该两种鱼种, 另外4个渔区的年平均渔获量前两位中均有两者之一。有研究表明, 海洋小型中上层鱼类占海洋物种的50%, 并且多集中在上升流区域[43], 且FAO主要渔区的主要鱼种大多为中上层鱼类[2]。由本文研究可知, 类群Ⅰ的中上层鱼类和底层鱼类的年平均渔获量相近, 而类群Ⅱ中中上层鱼类年平均渔获量远高于底层鱼类, 这可能和类群Ⅱ渔业资源丰富[36-37, 39, 42]、近年来加大的捕捞强度[40, 42], 以及渔区内以中上层鱼类为主要目标渔获物[2, 44]等因素有关, 且也证明类群Ⅱ渔区的渔业资源未完全开发, 底层鱼类等还有开发潜力。
本文利用灰色理论系统的关联度和聚类方法, 对全球渔区渔获量及关联渔区鱼种的关联聚类进行了分析, 并得到了较好的结果, 这有利于从宏观角度, 大尺度的把控全球主要渔区和鱼种的渔获量变化动态及相互间关联度情况, 从而为各区域渔业组织、各远洋渔业国家和地区针对不同类型渔区和鱼种, 制定和执行各项针对性的渔业管理措施提供依据。建议今后结合气候变化、海洋环境、生态系统等因素, 开展全球渔业资源可持续利用评价、全球渔区渔获产量预测等方面的研究。
| [1] |
TEH L C L, SUMAILA U R. Contribution of marine fisheries to worldwide employment[J]. Fish and Fisheries, 2013, 14(1): 77-88. DOI:10.1111/j.1467-2979.2011.00450.x |
| [2] |
FAO. The state of world fisheries and aquaculture 2020[R]. Rome: FAO, 2020.
|
| [3] |
FAO. FAO major fishing areas[EB/OL]. https://www.fao.org/fishery/zh/area/search.
|
| [4] |
陈新军. 远洋渔业概论-资源与渔场[M]. 北京: 科学出版社, 2018. CHEN X J. Distant-water fishery[M]. Beijing: Science Press, 2018. |
| [5] |
CHEUNG W W L, WATSON R, PAULY D. Signature of ocean warming in global fisheries catch[J]. Nature, 2013, 497(7449): 365-368. DOI:10.1038/nature12156 |
| [6] |
WATSON R A, CHEUNG W W L, ANTICAMARA J A, et al. Global marine yield halved as fishing intensity redoubles[J]. Fish and Fisheries, 2013, 14(4): 493-503. DOI:10.1111/j.1467-2979.2012.00483.x |
| [7] |
丁琪, 陈新军, 李纲, 等. 基于渔获统计的西北太平洋渔业资源可持续利用评价[J]. 资源科学, 2013, 35(10): 2032-2040. DING Q, CHEN X J, LI G, et al. Catch statistics and the sustainable utilization of northwest Pacific Ocean fishery resources[J]. Resources Science, 2013, 35(10): 2032-2040. |
| [8] |
张忠, 杨文波, 陈新军, 等. 基于渔获量平均营养级的东南大西洋渔业资源状况分析[J]. 海洋渔业, 2015, 37(3): 197-207. ZHANG Z, YANG W B, CHEN X J, et al. On fisheries resources in the southeast Atlantic based on the mean trophic level of catches[J]. Marine Fisheries, 2015, 37(3): 197-207. DOI:10.3969/j.issn.1004-2490.2015.03.001 |
| [9] |
鲁泉, 方舟, 李楠, 等. 以灰色系统理论模型为基础构建印度洋海洋捕捞渔获量预测模型[J/OL]. 水产学报: 1-8[2021-12-17]. http://kns.cnki.net/kcms/detail/31.1283.S.20210617.1608.004.html. LU Q, FANG Z, LI N, et al. Prediction model of fisheries catch based on GM(1, N)in the Indian Ocean[J/OL]. Journal of Fisheries of China: 1-8[2021-12-17]. http://kns.cnki.net/kcms/detail/31.1283.S.20210617.1608.004.html. |
| [10] |
刘思峰, 杨英杰, 吴利丰, 等. 灰色系统理论及其应用[M]. 7版. 北京: 科学出版社, 2014. LIU S F, YANG Y J, WU L F, et al. Grey system theory and its application[M]. 7th ed. Beijing: China Science Press, 2014. |
| [11] |
刘思峰, 杨英杰. 灰色系统研究进展(2004—2014)[J]. 南京航空航天大学学报, 2015, 47(1): 1-18. LIU S F, YANG Y J. Advances in grey system research(2004-2014)[J]. Journal of Nanjing University of Aeronautics & Astronautics, 2015, 47(1): 1-18. DOI:10.3969/j.issn.1005-1120.2015.01.001 |
| [12] |
鲁泉, 陈新军. 改革开放40年来中国渔业产业发展及十四五产量预测[J]. 上海海洋大学学报, 2021, 30(2): 339-347. LU Q, CHEN X J. Development of Chinese fishery industry in 40 years of reform and opening up and production forecast in the 14th five-year plan[J]. Journal of Shanghai Ocean University, 2021, 30(2): 339-347. |
| [13] |
陈新军. 灰色系统理论在渔业科学中的应用[M]. 北京: 中国农业出版社, 2003: 1-32. CHEN X J. Grey system theory in fisheries science[M]. Beijing: China Agriculture Press, 2003: 1-32. |
| [14] |
陈新军, 刘必林. 渔业资源生物学[M]. 北京: 科学出版社, 2017. CHEN X J, LIU B L. Fishery resources biology[M]. Beijing: China Science Press, 2017. |
| [15] |
SWARTZ W, SALA E, TRACEY S, et al. The spatial expansion and ecological footprint of fisheries (1950 to present)[J]. PLoS One, 2010, 5(12): e15143. DOI:10.1371/journal.pone.0015143 |
| [16] |
REID P C, JOHNS D G, EDWARDS M, et al. A biological consequence of reducing Arctic ice cover: arrival of the Pacific diatom Neodenticula seminae in the North Atlantic for the first time in 800000 years[J]. Global Change Biology, 2007, 13(9): 1910-1921. DOI:10.1111/j.1365-2486.2007.01413.x |
| [17] |
DODSON J J, TREMBLAY S, COLOMBANI F, et al. Trans-Arctic dispersals and the evolution of a circumpolar marine fish species complex, the capelin (Mallotus villosus)[J]. Molecular Ecology, 2007, 16(23): 5030-5043. DOI:10.1111/j.1365-294X.2007.03559.x |
| [18] |
VERMEIJ G J, ROOPNARINE P D. The coming Arctic invasion[J]. Science, 2008, 321(5890): 780-781. DOI:10.1126/science.1160852 |
| [19] |
WISZ M S, BROENNIMANN O, GRØNKJÆRP, et al. Arctic warming will promote Atlantic-Pacific fish interchange[J]. Nature Climate Change, 2015, 5(3): 261-265. DOI:10.1038/nclimate2500 |
| [20] |
PÖRTNER H O, KNUST R. Climate change affects marine fishes through the oxygen limitation of thermal tolerance[J]. Science, 2007, 315(5808): 95-97. DOI:10.1126/science.1135471 |
| [21] |
PÖRTNER H O, FARRELL A P. Physiology and climate change[J]. Science, 2008, 322(5902): 690-692. DOI:10.1126/science.1163156 |
| [22] |
SUNDAY J M, BATES A E, DULVY N K. Global analysis of thermal tolerance and latitude in ectotherms[J]. Proceedings of the Royal Society B: Biological Sciences, 2011, 278(1713): 1823-1830. DOI:10.1098/rspb.2010.1295 |
| [23] |
CHEUNG WW L, FRÖLICHER T L. Marine heat waves exacerbate climate change impacts for fisheries in the northeast Pacific[J]. Scientific Reports, 2020, 10: 6678. DOI:10.1038/s41598-020-63650-z |
| [24] |
张忠, 刘赫威, 余为, 等. 南极海冰变化驱动的海水降温效应对西南大西洋阿根廷滑柔鱼栖息地的影响[J]. 上海海洋大学学报, 2021, 30(6): 1101-1112. ZHANG Z, LIU H W, YU W, et al. Effects of Antarctic sea ice variations-driven seawater cooling on habitat pattern of Argentine shortfin squid Illex argentinus in the southwest Atlantic Ocean[J]. Journal of Shanghai Ocean University, 2021, 30(6): 1101-1112. |
| [25] |
张立. 西北太平洋渔业资源声学评估[D]. 厦门: 国家海洋局第三海洋研究所, 2017. ZHANG L. Acoustic assessment of fishery resources in the northwest Pacific Ocean[D]. Xiamen: Third Institute of Oceanography, State Oceanic Administration, 2017. |
| [26] |
顾惠庭, 龙红宝. 安哥拉和纳米比亚外海的主要渔业资源[J]. 现代渔业信息, 1988(6): 8-12. GU H T, LONG H B. Main fishery resources off the coast of Angola and Namibia[J]. Fishery Information & Strategy, 1988(6): 8-12. |
| [27] |
王佳迪, 李天, 于瑞, 等. 纳米比亚渔业现状、问题和建议[J]. 世界农业, 2012(1): 86-90. WANG J D, LI T, YU R, et al. Present situation, problems and suggestions of fisheries in Namibia[J]. World Agriculture, 2012(1): 86-90. DOI:10.3969/j.issn.1002-4433.2012.01.021 |
| [28] |
房俊晗, 任航, 罗莹, 等. 非洲沿海国家海洋渔业资源开发利用现状[J]. 热带地理, 2019, 39(2): 288-297. FANG J H, REN H, LUO Y, et al. Evaluation utilization state of marine fishery resources of Africa[J]. Tropical Geography, 2019, 39(2): 288-297. |
| [29] |
焦敏, 高郭平, 陈新军. 东北大西洋海洋捕捞渔获物营养级变化研究[J]. 海洋学报, 2016, 38(2): 48-63. JIAO M, GAO G P, CHEN X J. Changes in trophic level of marine catches in the northeast Atlantic[J]. Haiyang Xuebao, 2016, 38(2): 48-63. DOI:10.3969/j.issn.0253-4193.2016.02.005 |
| [30] |
郭文路, 黄硕琳, 曹世娟. 个体可转让配额制度在渔业管理中的运用分析[J]. 海洋通报, 2002, 21(4): 72-78. GUO W L, HUANG S L, CAO S J. Analysis on applying individual transferable quota system to the fishery management[J]. Marine Science Bulletin, 2002, 21(4): 72-78. DOI:10.3969/j.issn.1001-6392.2002.04.010 |
| [31] |
ANDREW J. The convention on the conservation and management of fishery resources in the south east Atlantic Ocean, 2001: an introduction[J]. The International Journal of Marine and Coastal Law, 2002, 17(1): 33-77. DOI:10.1163/15718080220493316 |
| [32] |
SEAFO. Conservation measure CM-TAC-01 (2020): on total allowable catches and related conditions for Patagonian toothfish, deep-sea red crab, alfonsino, orange roughy and pelagic armourhead for 2021 in the SEAFO convention area. Enter into Force: 1 January 2021.
|
| [33] |
DICOSIMO J, METHOT R D, ORMSETH O A. Use of annual catch limits to avoid stock depletion in the Bering Sea and Aleutian Islands management area (Northeast Pacific)[J]. ICES Journal of Marine Science, 2010, 67(9): 1861-1865. DOI:10.1093/icesjms/fsq060 |
| [34] |
TRUMBLE R J. Northeast Pacific flatfish management[J]. Journal of Sea Research, 1998, 39(1/2): 167-181. |
| [35] |
BERGSTAD OA, HØIMES ÅS, SARRALDE R, et al. Bathymetry, substrate and fishing areas of southeast Atlantic high-seas seamounts[J]. African Journal of Marine Science, 2019, 41(1): 11-28. DOI:10.2989/1814232X.2019.1569160 |
| [36] |
葛慧. 印度洋沿岸典型国家渔业管理政策法规及过洋性渔业合作分析[D]. 舟山: 浙江海洋大学, 2018: 10-50. GE H. The research on fishery management in coastal areas in India and the analysis of the pelagic cooperation[D]. Zhoushan: Zhejiang Ocean University, 2018: 10-50. |
| [37] |
杨晓明, 陈新军, 周应祺, 等. 基于海洋遥感的西北印度洋鸢乌贼渔场形成机制的初步分析[J]. 水产学报, 2006, 30(5): 669-675. YANG X M, CHEN X J, ZHOU Y Q, et al. A marine remote sensing-based preliminary analysis on the fishing ground of purple flying squid Sthenoteuthis oualaniensis in the northwest Indian Ocean[J]. Journal of Fisheries of China, 2006, 30(5): 669-675. |
| [38] |
鲁泉, 苏雪, 方舟, 等. 基于渔获量平均营养级的东印度洋渔业资源可持续利用评价[J]. 海洋学报, 2021, 43(8): 118-127. LU Q, SU X, FANG Z, et al. Evaluation of sustainable utilization of fishery resources in the eastern Indian Ocean based on the mean trophic level[J]. Haiyang Xuebao, 2021, 43(8): 118-127. |
| [39] |
沈卉卉, 黄硕琳. 中西太平洋金枪鱼渔业管理现状分析[J]. 上海海洋大学学报, 2014, 23(5): 789-795. SHEN H H, HUANG S L. An analysis of the tuna fishery management status in the western and central Pacific Ocean[J]. Journal of Shanghai Ocean University, 2014, 23(5): 789-795. |
| [40] |
丁琪. 全球海洋渔业资源可持续利用及脆弱性评价[D]. 上海: 上海海洋大学, 2017: 5-60. DING Q. Evaluation on sustainable utilization and vulnerability of global marine fishery resources[D]. Shanghai: Shanghai Ocean University, 2017: 5-60. |
| [41] |
丁琪, 陈新军, 耿婷, 等. 基于渔获统计的太平洋岛国渔业资源开发利用现状评价[J]. 生态学报, 2016, 36(8): 2295-2303. DING Q, CHEN X J, GENG T, et al. Evaluationon utilization state of marine fishery resources of Pacific Islands based on catch statistics[J]. Acta Ecologica Sinica, 2016, 36(8): 2295-2303. |
| [42] |
陈新军, 周应祺. 渔业导论[M]. 北京: 科学出版社, 2018. CHEN X J, ZHOU Y Q. Introduction of fishery[M]. Beijing: China Science Press, 2018. |
| [43] |
FRÉONP, CURY P, SHANNON L, et al. Sustainable exploitation of small pelagic fish stocks challenged by environmental and ecosystem changes: areview[J]. Bulletin of Marine Science, 2005, 76(2): 385-462. |
| [44] |
鲁泉, 方舟, 陈新军. 印度洋渔业资源可持续利用灰色关联评价[J]. 广东海洋大学学报, 2021, 41(5): 61-66. LU Q, FANG Z, CHEN X J. Evaluation of sustainable utilization of fishery resources in the Indian Ocean based on grey relational analysis[J]. Journal of Guangdong Ocean University, 2021, 41(5): 61-66. DOI:10.3969/j.issn.1673-9159.2021.05.008 |
2. National Engineering Research Center for Oceanic Fisheries, Shanghai 201306, China;
3. Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai 201306, China;
4. Key Laboratory of Oceanic Fisheries Exploration, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China;
5. Scientific Observing and Experimental Station of Oceanic Fishery Resources, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China