 2022,
Vol. 31
2022,
Vol. 31



2. 海州湾渔业生态系统教育部野外科学观测研究站, 山东 青岛 266003;
3. 青岛海洋科学与技术试点国家实验室 海洋渔业科学与食物产出过程功能实验室, 山东 青岛 266071;
4. 日照山海天旅游度假区渔政监督管理站, 山东 日照 276800
体长、体质量是鱼类最基础的生物学特征, 可以反应鱼类的生长模式以及生长环境的优劣。而体长-体质量关系参数也是一些资源评估模型的必需参数, 对渔业资源评估和管理具有重要意义。此外, 鱼类体长-体质量关系包含了丰富的生态学信息, 其时空差异性可能反映了生态系统特征的变化。许多研究[1]表明, 鱼类体长-体质量关系受到季节和地理空间以及环境因子的影响, 因此其关系参数可能存在异质性。而体长-体质量相关参数的时空异质性可能会对部分鱼类资源评估模型的评估精确性产生影响[1], 进而影响鱼类资源的有效管理。因此, 研究鱼类体长-体质量关系的时空异质性, 可以更好地了解鱼类种群动态的时空差异, 为渔业资源的管理与保护提供更精确的基础资料。以往异质性研究的基础方法是根据数据来源的不同构建多个模型, 然后对模型的参数进一步进行差异性比较, 分析过程比较繁琐。而线性混合效应模型(linear mixed-effects model, LMEM)作为一般线性模型的拓展, 通过加入多个随时间和空间等因子变化产生的随机效应[2], 能将所有数据来源产生的随机效应用一个模型表示, 因此可以分析多样化的数据, 近年来已广泛应用于渔业领域研究[3-7]。
星康吉鳗(Conger myriaster)隶属于鳗鲡目(Anguilliformes)康吉鳗科(Congtidae)康吉鳗属(Conger)[8]。该种在我国东海、黄海以及日本和朝鲜半岛海域均有分布[9]。由于其具有较高的生态价值和经济价值, 是我国东、黄海等海域的重要捕捞对象之一。近年来随着捕捞强度的增加和海洋环境的恶化, 星康吉鳗渔业产量呈波动下降趋势[10]。因此, 亟需对其渔业生物学特征与资源评估开展研究, 为该资源的合理开发和管理提供理论依据。有研究表明山东近海星康吉鳗的摄食强度[11-13]、资源密度以及空间分布[14-15]具有明显的季节变化, 且其生长会受到水域温度和饵料可获得性的共同调控[16], 由此推断其体长-体质量关系可能存在时空异质性。因此, 本研究基于2016—2017年山东近海4个航次的底拖网调查数据, 研究了山东近海星康吉鳗的体长、体质量和肥满度的时空规律, 并使用线性混合效应模型分析其体长-体质量关系的时空异质性, 以期加深对星康吉鳗生物学特征的理解, 为其渔业管理和资源的合理开发提供科学支撑。
1 材料与方法 1.1 数据来源星康吉鳗体长、体质量数据来自2016年10月和2017年1月、5月、8月在山东近海进行的渔业资源底拖网调查。调查海域为35.0°N~38.5°N和119°E~124°E, 共设置99个调查站位(图 1)。调查船为功率220 kW的单拖渔船, 其网口扩张宽度和高度分别为15 m和7.53 m, 囊网网目17 mm。调查站位拖网时间均为1 h, 拖速为3 kn。4个航次共捕获949尾星康吉鳗样品, 其中春季255尾, 夏季254尾, 秋季289尾, 冬季151尾(表 1)。根据水深, 将调查海域分为3个水域(图 1), 其中Ⅰ区水深为30 m以浅, Ⅱ区水深为30~50 m, Ⅲ区水深为50 m以深。按照《海洋调查规范第6部分: 海洋生物调查》(GB/T 12763.6—2007)进行样品的收集、分析与测定[17], 体长测量精确到0.1 cm, 体质量测量精确到0.01 g。文中所有体长均为星康吉鳗的全长。

|
Ⅰ、Ⅱ、Ⅲ区的水深分别为30 m以浅, 30~50 m, 50 m以深。 The depth of areaⅠ is less than 30 m, the depth of area Ⅱ is 30-50 m, and the depth of area Ⅲ is deeper than 50 m. 图 1 山东近海星康吉鳗的调查站位 Fig. 1 Survey station of Conger myriaster in the coastal waters of Shandong Province |

|
表 1 山东近海星康吉鳗在不同季节和水域的样本数量 Tab.1 Sample size of Conger myriaster in different seasons and regions in the coastal waters of Shandong Province |
根据生物学测定数据分析山东近海星康吉鳗在各个季节体长和体质量范围、优势组及其占比、中位数等参数。运用t检验分析各个季节之间的差异显著性。体长-体质量关系用幂指数方程表示[18]:
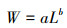 (1)
(1)
式中: W为星康吉鳗的体质量, g; L为星康吉鳗的体长, cm; a为条件因子, b为异速生长参数。当b < 3, 说明鱼体呈负异速生长, 即随鱼体长成, 肥满度下降, 身体趋于细长; 当b=3, 说明鱼体呈等速生长; 当b>3, 说明鱼体呈正异速生长, 即随鱼体长成, 肥满度升高。
采用Fulton状态指数K计算[19]:
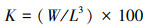 (2)
(2)
式中: W与L公式1。肥满度的季节差异显著性使用方差分析, 运用Surfer13软件制作4个季节个体肥满度的空间分布图。
1.2.2 线性混合效应模型线性混合效应模型的模型结构不仅包含固定效应, 还可以加入多个随时间和空间等因子变化产生的随机效应[5], 本研究拟合1个广义线性模型(generalized linear model, GLM)和9个线性混合效应模型(linear mixed-effects model, LMEM)来描述星康吉鳗体长与体质量的关系。线性混合效应模型用季节和水域的影响作为参数a、b的随机效应, 来阐明星康吉鳗体长与体质量关系的时空差异。9个LMEM模型在参数a或b是否包含水域或季节产生的随机效应的影响上均不同, 模型具体结构如表 4所示。模型构建过程通过R-4.0.2软件中“lme4”程序包完成[20], 并通过使用isSingular函数进一步确定模型是否出现过拟合现象。
利用赤池信息量准则(akaike Information criterion, AIC)筛选最优模型, 选取AIC最小的模型作为最优模型[21]。另外, 本文采用交叉验证来评估模型的预测性能, 随机抽取80%的数据作为训练集, 剩余20%为验证集, 使用训练集进行模型拟合, 并用训练集对所拟合模型进行预测, 将预测值和观测值进行对比检验, 重复以上过程100次, 得到该模型预测性能的评估结果[22]。再利用决定系数(R2)判断模型预测的性能, R2越接近1, 说明模型的预测值与样品的真实值越接近, 即表明模型的预测性能越好。
2 结果与分析 2.1 体长、体质量组成根据山东近海星康吉鳗体长、体质量组成, 分别分析了优势体长、体质量和均值等特征参数。春季优势体长为23.2~33.2 cm, 平均体质量为51.69 g;夏季优势体长为28.2~38.2 cm, 平均体质量为54.57 g;秋季优势体长为33.2~43.2 cm, 平均体质量为54.25 g;冬季优势体长为18.2~28.2 cm, 平均体质量为22.22 g;4个季节优势体长组占比均在45%以上。春、夏、秋、冬季样本的体长变异系数分别为22.94%、25.31%、24.53%、17.70%, 体质量变异系数分别为87.70%、83.07%、92.98%、78.21%。夏季和秋季航次的体长变异程度较其他季节大, 秋季航次的体质量变异程度最大。分析其不同季节体长、体质量频率分布图(图 2, 图 3), 由t检验得出, 冬季航次的体长和体质量显著小于其他季节(P<0.05),见表 2和表 3。

|
图 2 山东近海星康吉鳗各季节体长频率分布图 Fig. 2 Distribution of body length frequency of Conger myriaster in different seasons in the coastal waters of Shandong Province |

|
图 3 山东近海星康吉鳗各季节体质量频率分布图 Fig. 3 Distribution of body mass frequency of Conger myriaster in different seasons in the coastal waters of Shandong Province |

|
表 2 山东近海星康吉鳗各个季节体长t检验结果 Tab.2 Results of the t test for body length of Conger myriaster in each season in the coastal waters of Shandong Province |

|
表 3 山东近海星康吉鳗各个季节体质量t检验结果 Tab.3 Results of the t test for body mass of Conger myriaster in each season in the coastal waters of Shandong Province |
春季山东近海星康吉鳗的平均肥满度为0.120±0.015 9, 夏季为0.143±0.020 0, 秋季为0.150 2±0.053 4, 冬季为0.119 9±0.012 1。由t检验得出, 夏秋季航次个体的肥满度显著高于春冬季航次(P<0.01),见表 4。因此, 山东近海星康吉鳗的的季节间肥满度具有较大差异(图 4)。

|
表 4 山东近海星康吉鳗各个季节肥满度t检验结果 Tab.4 Results of the t test for condition factor of Conger myriaster in each season in the coastal waters of Shandong Province |

|
图 4 山东近海星康吉鳗各个季节平均肥满度的时间变化 Fig. 4 Seasonal variation of relative fatness of Conger myriaster in the coastal waters of Shandong Province |
基于对山东近海采集的949尾星康吉鳗样品的分析, 得出全部星康吉鳗样品的体长-体质量的幂函数关系式为W=0.000 4×L3.34(R2=0.96, n=949)。通过AIC值最小原则筛选最优模型, 结果表明: LMEM(R&T.I)为最优模型, 其AIC值为-1 098(表 5)。即表明在拟合的线性混合效应模型中, 包含季节和水域对参数a的随机效应的混合效应模型对星康吉鳗的体长-体质量关系的拟合效果最优。该模型中, 参数b的固定值为3.27, 参数a为4.78×10-4~5.68×10-4, 夏季的a值最大, 为5.68×10-4, 秋季和冬季次之, 在春季最小, 为4.78×10-4; Ⅲ区(50 m以深海域)最大, 为5.20×10-4; Ⅰ区(30 m以浅海域)次之, 为5.16×10-4; Ⅱ区(30 m~ 50 m海域)最小, 为5.06×10-4。另外, 交叉验证的结果(表 5): 其R2最接近1, 也表明此模型的预测效果最好, 且9个线性混合效应模型的R2值均大于GLM, 说明线性混合效应模型的预测效果优于传统模型。

|
表 5 山东近海星康吉鳗体长-体质量关系模型 Tab.5 Length-weight relationship models of Conger myriaster in the coastal waters of Shandong Province |
当星康吉鳗体长小于20 cm时(图 5), 其体质量的季节和水域差异均较小; 当体长大于20 cm时, 其体质量的时空差异较大。当体长为20~40 cm时, 夏季的体质量增长率明显大于其他三个季节, 而Ⅰ、Ⅱ、Ⅲ区星康吉鳗的体质量增长率的曲线几乎重合。当体长大于40 cm时, 其体质量增长率在季节上的大小: 夏季>秋季>冬季>春季, 在空间上的大小: Ⅲ区>Ⅰ区>Ⅱ区。

|
图 5 山东近海星康吉鳗在不同季节和水域的体长-体质量关系差异 Fig. 5 Variations of length-mass relationships for Conger myriaster among seasons and regions in the coastal waters of Shandong Province |
山东近海星康吉鳗体长、体质量、体长-体质量关系参数以及肥满度在时空上均有较大波动, 且季节间差异显著。冬季个体的平均体长、体质量显著小于其他3个航次; 肥满度季节间差异显著, 且夏季肥满度显著大于其他季节。其体长与体质量关系式为W=0.000 4×L3.34, b的固定值大于3, 表明星康吉鳗呈正异速生长, 此研究结果与东、黄海星康吉鳗群体的体长-体质量参数的相关研究[23]一致。即说明星康吉鳗随鱼体的增长, 肥满度呈升高的趋势。线性混合效应模型表明, 星康吉鳗体长-体质量关系参数条件因子a具有显著的时空差异: 夏季的a值最大, 秋季和冬季次之, 春季最小; Ⅲ区(30 m以深海域)最大, Ⅰ区(30 m以浅海域)次之, Ⅱ区(30 m~50 m海域)最小。山东近海星康吉鳗体长-体质量分布特征及其相关参数的季节和水域变化可能与其洄游习性、摄食强度以及饵料丰度等有关。
3.1 体长、体质量、肥满度的时空变化山东近海星康吉鳗体长、体质量组成存在明显的时空变化。其体长、体质量存在一定的季节差异, 冬季群体的体长和体质量显著小于其他3个季节(P<0.05)。冬季的星康吉鳗群体个体较小可能与其洄游习性有关。春、夏季近岸水温较高、饵料丰富, 星康吉鳗聚集在近岸栖息、索饵。秋后, 星康吉鳗随鱼体长成并向深水海域迁移, 越冬或参加产卵洄游[24], 而幼鳗在近海海域随底层水温变化做适温迁移, 所以冬季群体可能主要由个体较小的幼鱼组成。星康吉鳗体长、体质量组成的空间分布上总体呈现远岸深水区大于近岸浅水海域的特征, 进一步验证了该鱼种随鱼体长成向深水区洄游的生活史特性[24]。
山东近海星康吉鳗肥满度存在明显的季节变化。夏秋季节肥满度显著高于春冬季, 这可能与水温以及饵料丰度有关。海水温度与饵料丰度是影响鱼类生长的主要环境因子。在鱼类的适温范围内, 温度的升高会促进鱼体的新陈代谢, 促进鱼类的摄食行为。相关研究指出当水温低于8 ℃时, 星康吉鳗的摄食和游泳速度会受到影响[25]。在水温和食物丰度相对较低的春、冬季节, 星康吉鳗摄食强度较低[11], 为维持机体的正常代谢和活动消耗, 须依靠自身体内的营养来补充机体活动所需的能量, 从而个体肥满度较低。夏秋季节, 水温较高、食物充沛, 星康吉鳗摄食强度达到一年之中最大值[11], 此时积极索饵育肥, 为洄游和越冬储存能量, 此时个体肥满度较高。
3.2 混合效应模型研究体长-体质量关系时空异质性的优势异质性研究的基础方法是根据数据来源的不同构建多个模型, 然后对模型的参数进一步进行差异性比较。如栾静等[1]采用海州湾8个航次的渔业资源底拖网调查数据, 拟合了8个体长体质量模型, 来研究海州湾方氏云鳚(Pholis fangi)体长与体质量分布特征及其关系; GONZÃLEZ等[26]依据2014—2017年在密苏河采集的密苏里铲鲟(Scaphirhynchus Sturgeon)的生物学数据, 建立了8个生长方程; 这些单独构建的模型得出的估计值, 还需要进一步对其进行异质性比较, 而混合效应模型可以在单个模型中包含多个随时间和空间等因子变化产生的随机效应, 能更简单、准确地进行异质性研究。衷思剑等[4]根据山东近海2016—2017年4个季节的调查数据, 在模型中考虑季节和水域因素, 构建了1个广义线性模型和9个线性混合效应模型, 发现季节和纬度对黄(Lophius litulon)的体长-体质量关系具有显著的影响。陈锦辉等[5]用2013—2018年采集的印度洋大眼金枪鱼(Thunnus obesus)的相关数据, 拟合了6个线性混合效应模型来分析不同年份、季度和海域间对生长的影响。朱文斌等[7]根据2019年4—6月在浙江沿岸海域日本鳀(Engraulis japonicus)幼鱼样品, 拟合1个广义线性模型和9个线性混合效应模型发现日本鳀幼鱼的叉长与体质量关系会随着月份和水域变化。研究也发现山东近海星康吉鳗的体长-体质量关系具有时空差异。另外, 9个线性混合效应模型的R2均大于GLM, 说明线性混合效应模型的预测效果优于传统模型, 进一步验证了线性混合效应模型在数据来源异质性研究中的优势。
基于LMEM(R&T.I)的结果, 得出星康吉鳗体长与体质量关系呈现明显的时空变化, 这与在小黄鱼(Larimichthys polyactis)[3]和黄[4]的研究结果相同。其中条件因子a为6.28×10-4~7.44×10-4, 均在前人研究[27]的鳗形鱼类的a值范围(4.64×10-4~2.25×10-3)之内。研究表明, 夏季和秋季的a值显著高于春季和冬季, 50 m以深海域的a值最高。条件因子a与鱼体的肥满度、生长环境等有关[28], 造成参数a的季节性差异可能与星康吉鳗的摄食强度、季节性洄游以及饵料丰度的季节变化有关。有学者[11]曾发现, 星康吉鳗夏季和秋季的摄食强度较高, 积极索饵, 造成星康吉鳗夏、秋季的a值高于春、冬季。另外, 夏、秋季节的水温高, 饵料更为丰富, 丰富的饵料更有利于星康吉鳗的生长, 并且秋后, 星康吉鳗随鱼体长成向深水海域迁移, 越冬或参加产卵洄游。研究发现, a值在Ⅰ区(30 m以浅海域)和Ⅲ区(50 m以深海域)中较Ⅱ区(30~50 m海域)高, 这表明Ⅰ区和Ⅲ区的星康吉鳗处于适宜的生长环境, 李明坤等[14]认为星康吉鳗渔获率随水深的增加, 呈现先增加后减小再增加的变化趋势, 也反映出Ⅰ区和Ⅲ区的星康吉鳗具有更好的生长条件。
采用线性混合效应模型研究山东近海星康吉鳗体长、体质量及其关系参数的时空异质性。结果表明, 季节和水域对山东近海星康吉鳗体长-体质量关系参数a有显著影响。因此, 一些种群评估模型使用体长-体质量等相关参数时, 应当充分考虑季节和水域对于相关参数的影响, 以提高评估精确性。如在评估鱼类的可持续产量、制定生物学管理参考点时, 应该充分考虑体长-体质量参数的时空变化, 减少评估的不确定性, 为渔业资源的保护和管理制定更加科学合理的管理策略。文中未检验季节与空间对体长-体质量关系式参数b的影响, 原因是我们当前的数据量不支持复杂的随机效应结构(如随机斜率项, 即对参数b的影响)导致复杂的模型因随机效应间的共线性而出现过度拟合。此外, 研究只考虑季节和水域的随机效应, 而年间小尺度环境变化也可能对鱼类生长产生一定影响, 因此, 未来可考虑星康吉鳗体长-体质量关系的年际变化, 探讨环境因子对其关系参数的影响, 进一步为星康吉鳗资源的合理开发和管理提供科学依据和参数支撑。
| [1] |
栾静, 徐宾铎, 薛莹, 等. 海州湾方氏云鳚体长与体重分布特征及其关系[J]. 中国水产科学, 2017, 24(6): 1323-1331. LUAN J, XU B D, XUE Y, et al. Size distribution and length-weight relationships in Pholis fangi in Haizhou Bay[J]. Journal of Fishery Sciences of China, 2017, 24(6): 1323-1331. |
| [2] |
XU H, SUN Y J, WANG X J, et al. Linear mixed-effects models to describe individual tree crown width for China-fir in Fujian province, southeast China[J]. PLoS ONE, 2015, 10(4): e0122257. DOI:10.1371/journal.pone.0122257 |
| [3] |
MA Q Y, JIAO Y, REN Y P. Linear mixed-effects models to describe length-weight relationships for yellow croaker (Larimichthys polyactis) along the north coast of China[J]. PLoS ONE, 2017, 12(2): e0171811. DOI:10.1371/journal.pone.0171811 |
| [4] |
衷思剑, 麻秋云, 刘淑德, 等. 基于线性混合效应模型的黄体长体重关系的时空差异[J]. 中国水产科学, 2018, 25(6): 1299-1307. ZHONG S J, MA Q Y, LIU S D, et al. Linear mixed-effects models for estimating spatiotemporal variations of length-weight relationships for Lophius litulon[J]. Journal of Fishery Sciences of China, 2018, 25(6): 1299-1307. |
| [5] |
陈锦辉, 王学昉, 田思泉, 等. 基于混合效应模型的印度洋大眼金枪鱼生长特征异质性分析[J]. 中国水产科学, 2020, 27(5): 570-578. CHEN J H, WANG X F, TIAN S Q, et al. Growth heterogeneity of bigeye tuna (Thunnus obesus) in the Indian Ocean explored by the mixed effects model[J]. Journal of Fishery Sciences of China, 2020, 27(5): 570-578. |
| [6] |
王杨楠, 韩东燕, 麻秋云, 等. 基于线性混合效应模型的镰鲳叉长-体质量关系的异质性分析[J]. 上海海洋大学学报, 2021, 30(1): 135-143. WANG Y N, HAN D Y, MA Q Y, et al. Linear mixed-effect models for Heterogeneity analysis of fork length-mass relationships for Pampus echinogaster[J]. Journal of Shanghai Ocean University, 2021, 30(1): 135-143. |
| [7] |
朱文斌, 朱海晨, 王雅丽, 等. 基于线性混合效应模型的日本鳀幼鱼叉长-体重关系异质性[J]. 应用生态学报, 2021, 32(12): 4532-4538. ZHU W B, ZHU H C, WANG Y L, et al. Heterogeneity of fork length-weight relationship for juvenile Engraulis japonius based on linear mixed-effects models[J]. Chinese Journal of Applied Ecology, 2021, 32(12): 4532-4538. |
| [8] |
李明德. 中国经济鱼类生态学[M]. 天津: 天津科学技术出版社, 2005: 12-13. LI M D. Ecology of Chinese economic fishes[M]. Tianjin: Tianjin Science and Technology Press, 2005: 12-13. |
| [9] |
唐逸民, 吴常文. 星鳗Astroconger myriaster (Brevoort)的生物学特性与资源分布[J]. 浙江水产学院学报, 1988, 7(1): 19-26. TANG Y M, WU C W. On biologic habits and resources distribution of common Japanese conger Astroconger myriaster (Brevoort)[J]. Journal of Zhejiang College of Fisheries, 1988, 7(1): 19-26. |
| [10] |
娄甜甜, 齐兴柱, 尹绍武, 等. 鳗鲡种质资源的研究进展[J]. 水产科学, 2007, 26(6): 366-369. LOU T T, QI X Z, YIN S W, et al. Recent research on germplasm resources in eels (Genus anguilla)[J]. Fisheries Science, 2007, 26(6): 366-369. DOI:10.3969/j.issn.1003-1111.2007.06.015 |
| [11] |
刘西方, 刘贺, 薛莹, 等. 海州湾星康吉鳗的摄食生态特征[J]. 中国水产科学, 2015, 22(3): 517-527. LIU X F, LIU H, XUE Y, et al. Feeding ecology of Conger myriaster in Haizhou Bay[J]. Journal of Fishery Sciences of China, 2015, 22(3): 517-527. |
| [12] |
刘西方. 海州湾两种高营养级鱼类摄食生态及其食物关系研究[D]. 青岛: 中国海洋大学, 2015. LIU X F. Study on feeding ecology and food relations of two high trophic level fishes in Haizhou Bay[D]. Qingdao: Ocean University of China, 2015. |
| [13] |
刘鸿雁, 孙彤彤, 曾晓起, 等. 崂山湾人工鱼礁区星康吉鳗摄食生态及食物网结构[J]. 应用生态学报, 2018, 29(4): 1339-1351. LIU H Y, SUN T T, ZENG X Q, et al. Feeding ecology of Conger myriaster and structure of the food webs in artificial reef zone, Laoshan Bay, China[J]. Chinese Journal of Applied Ecology, 2018, 29(4): 1339-1351. |
| [14] |
李明坤, 张崇良, 李敏, 等. 山东南部近海秋、冬季星康吉鳗分布与环境因子的关系[J]. 中国水产科学, 2018, 25(5): 1115-1122. LI M K, ZHANG C L, LI M, et al. Relationship between the spatiotemporal distribution of Conger myriaster and environmental factors in the southern waters of the Shandong Peninsula during autumn and winter[J]. Journal of Fishery Sciences of China, 2018, 25(5): 1115-1122. |
| [15] |
李雪童, 王琨, 徐宾铎, 等. 山东近海鱼类群落种类组成与空间结构的周年变化[J]. 水产学报, 2021, 45(4): 552-562. LI X T, WANG K, XU B D, et al. Annual variation of species composition and spatial structure of fish community in Shandong offshore[J]. Journal of fisheries of China, 2021, 45(4): 552-562. |
| [16] |
MU X X, ZHANG C L, XU B D, et al. Varying growth rates of a marine eel, the whitespotted conger (Conger myriaster), are explained by the interaction between seasonal temperature and prey availability[J]. Marine Biology, 2022, 169(1): 6. DOI:10.1007/s00227-021-03976-y |
| [17] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/T 12763.6-2007海洋调查规范第6部分: 海洋生物调查[S]. 北京: 中国标准出版社, 2008: 6-17. General Administration of Quality Supervision, Inspection and Quarantine of the People's Republic of China, Standardization Administration of the People's Republic of China. GB/T 12763.6-2007 Specifications for oceanographic survey-Part 6: Marine biological survey[S]. Beijing: Standards Press of China, 2008: 6-17. |
| [18] |
KEYS A B. The weight-length relation in fishes[J]. Proceedings of the National Academy of Sciences of the United States of America, 1928, 14(12): 922-925. DOI:10.1073/pnas.14.12.922 |
| [19] |
戴强, 戴建洪, 李成, 等. 关于肥满度指数的讨论[J]. 应用与环境生物学报, 2006, 12(5): 715-718. DAI Q, DAI J H, LI C, et al. Discussion on relative fatness[J]. Chinese Journal of Applied and Environmental Biology, 2006, 12(5): 715-718. DOI:10.3321/j.issn:1006-687X.2006.05.026 |
| [20] |
BATES D, MÄCHLER M, BOLKER B, et al. Fitting linear mixed-effects models using lme4[J]. Journal of Statistical Software, 2015, 67(1): 1-48. |
| [21] |
BURNHAM K P, ANDERSON D R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach[M]. 2nd ed. New York: Springer, 2002: 484-488.
|
| [22] |
TANAKA K, CHEN Y. Spatiotemporal variability of suitable habitat for American lobster (Homarus americanus) in Long Island Sound[J]. Journal of Shellfish Research, 2015, 34(2): 531-543. DOI:10.2983/035.034.0238 |
| [23] |
麻秋云, 牟秀霞, 任一平, 等. 东、黄海星康吉鳗生长、死亡和单位补充量渔获量[J]. 水产学报, 2018, 42(6): 881-888. MA Q Y, MU X X, REN Y P, et al. The growth, mortality and yield per recruitment of white-spotted conger (Conger myriaster) in the Yellow Sea and the East China Sea[J]. Journal of Fisheries of China, 2018, 42(6): 881-888. |
| [24] |
陈大刚. 黄渤海渔业生态学[M]. 北京: 海洋出版社, 1991: 478-480. CHEN D G. Fisheries ecology of Yellow Sea and Bohai Sea[M]. Beijing: China Ocean Press, 1991: 478-480. |
| [25] |
LI M, ZHANG C L, XU B D, et al. Evaluating the approaches of habitat suitability modelling for whitespotted conger (Conger myriaster)[J]. Fisheries Research, 2017, 195: 230-237. DOI:10.1016/j.fishres.2017.07.024 |
| [26] |
GONZÃLEZ A, LONG J M, GOSCH N J C, et al. Spatial and temporal variation in length-weight relationships of age-0 Scaphirhynchus sturgeon in the Lower Missouri River[J]. The American Midland Naturalist, 2021, 186(1): 106-121. |
| [27] |
FROESE R, THORSON J T, REYES R B JR. A Bayesian approach for estimating length-weight relationships in fishes[J]. Journal of Applied Ichthyology, 2014, 30(1): 78-85. DOI:10.1111/jai.12299 |
| [28] |
RICKER W E. Methods for assessment of fish production in fresh waters[M]. Oxford: Blackwell Scientific Press, 1968: 93-123.
|
2. Field Observation and Research Station of Haizhou Bay Fishery Ecosystem, Ministry of Education, Qingdao 266003, Shandong, China;
3. Laboratoray for Marine Fishery Science and Food Production Processes, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, Shandong, China;
4. Fishery Administration Supervision and Management Station of Rizhao Shanhaitian Tourist Resort, Rizhao 276800, Shandong, China