 2022,
Vol. 31
2022,
Vol. 31

2. 中国水产科学研究院长江水产研究所 农业农村部淡水鱼类种质资源与生物技术重点实验室, 湖北 武汉 430223;
3. 巢湖管理局渔政管理总站, 安徽 合肥 238001
 (Paracanthobrama guichenoti)、鳜(Siniperca chuatsi)等22种鱼类均为偶见种。不同样点的指示物种差异明显,季节间和样点间鱼类群落组成均有一定程度的分离,样点间鱼类物种组成结构较季节间更加离散。鱼类多样性较低,各样点的Shannon-Wiener多样性指数(H')、Simpson多样性指数(D)、Margalef丰富度指数(d)、Pielou均匀度指数(J)的变化幅度分别为0.56~1.09、0.25~0.57、1.45~3.82、0.19~0.32,2018年夏季各项多样性指数高于其他季节,杭埠河口、东湖浅水区域高于其它样点。研究结果补充了巢湖鱼类群落的基础数据,为巢湖鱼类资源保护与管理提供科学依据。
(Paracanthobrama guichenoti)、鳜(Siniperca chuatsi)等22种鱼类均为偶见种。不同样点的指示物种差异明显,季节间和样点间鱼类群落组成均有一定程度的分离,样点间鱼类物种组成结构较季节间更加离散。鱼类多样性较低,各样点的Shannon-Wiener多样性指数(H')、Simpson多样性指数(D)、Margalef丰富度指数(d)、Pielou均匀度指数(J)的变化幅度分别为0.56~1.09、0.25~0.57、1.45~3.82、0.19~0.32,2018年夏季各项多样性指数高于其他季节,杭埠河口、东湖浅水区域高于其它样点。研究结果补充了巢湖鱼类群落的基础数据,为巢湖鱼类资源保护与管理提供科学依据。湖泊是陆地水圈的重要组成部分,在大自然水分循环、能量流动、气候调节、生物多样性维持等方面发挥重要作用[1]。我国大多数湖泊生物资源未得到合理的开发利用,近年来水体富营养化、生物多样性下降、生态功能衰退等问题日益严重[2]。鱼类是水生态系统的重要组成部分,对维系水生态系统结构与功能稳定具有重要作用,是关注度最高的水生生物类群之一[3-4]。
巢湖位于长江下游北岸,水域面积约770 km2,是我国“五大淡水湖”之一,也是“引江济淮”工程中连接长江和淮河的重要枢纽和合肥市重要的饮用水源地[5]。巢湖流域面积约13 500 km2,经裕溪河连接长江,主要入湖河流有杭埠河、南淝河、柘皋河、白石天河、马尾河、派河等河流[6]。1960年和1966年分别建成的巢湖闸、裕溪闸阻断了巢湖与长江的自然连通,巢湖成为人工控制水位的半封闭湖泊。巢湖渔业资源丰富,历史共记录鱼类94种[7]。自20世纪80年代以来,随着人口的增长和经济的发展,外源污染物大量输入巢湖,导致巢湖水域水体富营养化问题逐渐加剧,蓝藻“水华”频发,水体生态功能衰退,加之捕捞强度过大、水利工程修建、水生植物衰退和江湖连通受阻,巢湖中优质渔业资源不断减少,鱼类群落结构发生了显著的变化[8-10]。
自20世纪60年代开始,学者对巢湖鱼类进行了多次调查[7, 10],最近一次调查为过龙根等[8]于2002—2004年开展。此前对巢湖鱼类群落的研究主要集中在鱼类种类名录和渔业捕捞占比统计,缺乏鱼类群落结构和多样性的研究。基于2017—2018年对巢湖不同生境类型鱼类的调查,分析了巢湖鱼类群落结构和多样性空间分布特征。随着“引江济淮”工程的推进,巢湖即将恢复与长江的连通,并与淮河连通;此外,巢湖于2020年1月1日起实施为期10年的全湖禁捕。本研究通过分析巢湖鱼类群落的时空差异与生物多样性变化,为巢湖的鱼类现状提供本底资料,为今后巢湖的生态调水、渔业资源管理与保护提供科学依据。
1 材料与方法 1.1 样本采集巢湖的主要入湖河流中,杭埠河发源于大别山区,水质好,水量大,是巢湖最大入湖河流和最重要的清水来源;南淝河流经合肥市区,水质较差,是巢湖重要污染负荷来源;柘皋河周边土地利用类型以农田为主,水质轻度污染。[11]2017和2018年,在杭埠河口、南淝河口、柘皋河口和巢湖东部、西部和中部的敞水区和岸边浅水区,共设置9个样点进行鱼类调查(图 1)。采样时间为每年7、10、12月,由于1—6月为巢湖禁渔期,未能开展春季采样。每个样点使用20个定置(串联)倒须笼壶(俗称地笼网)(截面30 cm × 30 cm,网目2 a = 1 cm,长10 m)、1 200 m多网目组合三重刺网(网目2 a=2 cm、2 a=4 cm、2 a=6 cm、2 a=10 cm刺网各300 m,网高2.5 m)和拖网(网口直径4 m,网目2 a = 1.2 cm)采集鱼类,地笼提前两天放置,刺网于采样前一天下午放置,采样当天上午收刺网和地笼,下午用拖网采样,拖网使用渔船拖行约1 000 m,拖网速度约为5 km/h。对采集到的渔获物参照《中国动物志: 硬骨鱼纲鲤形目(中卷)》[12]和《中国淡水鱼类检索》[13]进行分类、计数并称体质量(精确到0.1 g),参照《江苏鱼类志》[14]和《浙江动物志淡水鱼类》[15]进行生态类型划分,专著中未记录的鱼类查阅相关文献确定。此外,调查渔民捕捞渔获物,以补充鱼类名录。

|
NF. 南淝河口;HB. 杭埠河口;ZG. 柘皋河口;E1. 东部敞水区;E2. 东部浅水区;M1. 中部敞水区;M2. 中部浅水区;W1. 西部敞水区;W2西部浅水区。 NF. estuary of Nanfei River; HB. estuary of Hangbu River; ZG. estuary of Zhegao River; E1. eastern open water area; E2. eastern shallow water area; M1. middle open water area; M2. middle shallow water area; W1. western open water area; W2. western shallow water area. 图 1 巢湖鱼类调查样点 Fig. 1 Fish sampling sites in the Chaohu Lake |
使用Shannon-Wiener多样性指数(H′)、Simpson多样性指数(D)、Margalef丰富度指数(d)和Pielou均匀度指数(J)表征鱼类多样性变化结构[16],计算公式如下:
Shannon-Wiener多样性指数(H′)
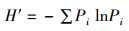 (1)
(1)
Simpson多样性指数(D)
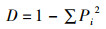 (2)
(2)
Margalef丰富度指数(d)
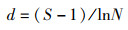 (3)
(3)
Pielou均匀度指数(J)
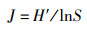 (4)
(4)
式中:Pi为第i种鱼类个体数占总个体数的比例;N为总个体数;S为样点总鱼类种数。
鱼类重要性指数由相对重要指数(Index of Relative Importance,IRI)来确定:
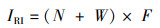 (5)
(5)
式中:N为第i种鱼的尾数占总尾数的百分比;W为第i种鱼的质量占总质量的百分比;F为第i种鱼出现次数占总调查样点的百分比。IRI≥1 000为优势种,100≤IRI < 1 000为重要种,10≤IRI < 100为常见种,1≤IRI < 10为一般种,IRI < 1为偶见种。[17]
使用指示物种分析(indicator species analysis,ISA)检验物种在生态区域内的亲和力关系[18];通过位点×物种矩阵进行非度量多维标尺(nonmetric multidimensional scaling,NMDS)排序,评估鱼类物种群落结构的不同时间和空间格局下的差异[19];采用Bray-Curtis相异距离构建物种在样点之间的距离矩阵, 采用置换多元方差分析(permutational multivariate ANOVA,PERMANOVA)检验鱼类群落结构在时空上的差异性和离散性,根据线性模型直接划分距离矩阵中的变化模式,计算基于距离矩阵的多元模型中每一项的Fisher’s F-ratio值,并使用置换检验计算显著性P值[20]。以上数据分析均在R语言中的vegan软件包中进行。
2 结果与分析 2.1 鱼类物种组成结构共采集到鱼类50种,渔民渔获物中补充采集到吻

|
表 1 2017—2018年巢湖调查渔获物种类组成 Tab.1 Species composition of fish collected from the Chaohu Lake from 2017 to 2018 |
物种多度方面:刀鲚个体数占比最高为66.5%;其次是太湖新银鱼(Neosalanx taihuensis),占30.34%。在9个样点6次调查过程中,刀鲚、太湖新银鱼、鲫(Carassius auratus)、鲢(Hypophthalmichthys molitrix)、鳙(Aristichthys nobilis)和翘嘴鲌(Culter alburnus)出现频率均为100%,鲤(Cyprinus carpio)、蒙古鲌(Culter mongolicus)等17种鱼类的出现频率超过50%,而青鱼(Mylopharyngodon piceus)、草鱼(Ctenopharyngodon idella)等12种鱼类的出现频率不到10%。重要性方面:刀鲚、太湖新银鱼、鲢、鳙的相对重要性指数分别为8 885.42、4 011.02、3 217.67、1 042.94,为优势种;鲤、鲫、翘嘴鲌、蒙古鲌为重要种;鳊(Parabramis pekinensis)、团头鲂(Megalobrama amblycephala)、须鳗虾虎鱼(Taenioides cirratus)、大银鱼(Protosalanx hyalocranius)等12种鱼为常见种;间下

物种指示值分析结果显示,在南淝河口样点,须鳗虾虎鱼、圆尾斗鱼(Macropodus ocellatus)为主要指示物种,指示值分别为0.479、0.208;杭埠河口样点中的主要指示物种为须鳗虾虎鱼、大鳍

|
表 2 各采样点主要指示鱼类的物种指示值 Tab.2 Indicator value of main indicative fish species in each sampling site |
使用非度量多维标尺排序方法处理不同样点物种多度数据矩阵,分析结果显示,季节间和区域间鱼类群落组成均有一定程度的分离。在时间尺度下,冬季12月采样点渔获物组成与7月和10月物种组成结构之间存在一定的分离,夏季7月和秋季10月物种组成结构差异性不明显(表 3)。多元方差置换检验分析PERMANOVA结果显示,不同采样时间物种组成结构的差异性达到显著性水平(F=1.93,P=0.016), 其中,差异性主要表现在2017年12月物种组成结构与2018年各个采样季节之间(图 2)。

|
表 3 时间尺度下鱼类物种组成的置换方差检验 Tab.3 Permutation test on fish species compositions on time cale |

|
点表示单个样点单次采样物种组成结果,单次采样不同样点之间用椭圆圈框表示样点间物种组成的离散程度,黑色点和圈表示标注时间不同样点的采样结果。 The point shows the species composition of a single sampling site at a sampling time. The points of different sampling sites at a sampling time are framed by an ellipse to indicate the dispersion degree of species composition among sampling sites. The species composition of different sampling sites at each specific sampling time is marked with black points and ellipse. 图 2 鱼类群落时间变化的NMDS排序图 Fig. 2 NMDS ordination of temporal variation of fish community |
物种组成结构在不同采样点之间存在明显的差异(图 3)。南淝河口、杭埠河口、东湖1号、东湖2号和西湖1号样点物种组成呈现单独聚集状态,鱼类物种组成结构特异性较强;柘皋河口和西湖2号样点与其他样点鱼类物种组成呈现显著的交叉,物种组成结构相似;中湖样点在空间排序中处于中间位置,说明物种组成结构相对泛化,与其他样点之间存在明显交叉。不同样点间鱼类物种组成结构差异性较明显。

|
点表示单个样点单次采样物种组成结果,单次采样不同样点之间用椭圆圈框表示样点间物种组成的离散程度,黑色点和圈表示标注时间不同样点的采样结果。 The point shows the species composition of a single sampling site at a sampling time. The points of different sampling sites at a sampling time are framed by an ellipse to indicate the dispersion degree of species composition among sampling sites. The species composition of different sampling sites at each specific sampling time is marked with black points and ellipse. 图 3 鱼类群落空间变化的NDMS排序图 Fig. 3 NMDS ordination of spatial variation of fish community |
各样点在各季节的Shannon-Wiener多样性指数(H′)的变动范围为0.56~1.09,Simpson多样性指数(D)的变动范围为0.25~0.57,Margalef丰富度指数(d)变化范围为1.45~3.82,Pielou均匀度指数(J)变化范围为0.19~0.32, 见图 4,季节和区域对各项多样性指数均有影响。

|
图 4 不同时间和样点的鱼类生物多样性指数 Fig. 4 Biodiversity indices of fish community at different time and sites |
在时间尺度下,2018年夏季各项多样性指数最高,H′、D、d和J的各样点均值分别为0.92、0.53、2.88和0.28。Simpson多样性指数和Pielou均匀度指数在季节间没有显著差异,Shannon-Wiener多样性指数的季节间的差异性主要表现在2017年7月与2018年12月之间,Margelef多样性指数季节间的差异性主要表现在2017年7月与2018年3次调查之间以及2017年10月与2018年3次调查之间。
空间格局下,Shannon-Wiener多样性指数在杭埠河口和东湖较高,区域间差异主要是东湖、杭埠河口与西湖、中湖样点之间;Simpson多样性指数在杭埠河口、西湖2号和中湖1号样点较高,南淝河口最低;Margelef丰富度指数东湖2号点最高, 区域间差异性主要是杭埠河口、东湖1号、2号点与南淝河口、西湖1号、2号点之间;Peilou均匀度指数样点间差异性较小。物种多样性指数在区域间的差异性大于季节间的差异性。
3 讨论 3.1 巢湖鱼类组成变化从20世纪60年代开始,学者多次对巢湖鱼类进行了调查。王岐山[10]在1959至1963年连续对巢湖鱼类进行调查,共记录鱼类85种;1973年组织的巢湖鱼类资源调查,共采集到鱼类62种[21];1980年组织开展的巢湖鱼类调查,共记录78种[7]。以上每次调查的鱼类组成都略有不同,共记录鱼类94种。过龙根等[8]2002—2004年开展的巢湖鱼类资源调查,仅记录54种。本次调查使用了刺网、地笼、拖网等多种网具,调查区域涵盖湖区主要生境类型,仅记录52种,且22种为偶见种,与2002—2004年的调查结果较为吻合。与20世纪调查结果相比,本次调查到的鱼类物种数大幅度减少,物种组成发生显著变化。共采集到5种洄游性鱼类,历史记录存在的暗纹东方鲀(Takifugu obscurus)、胭脂鱼(Myxocyprinus asiaticus)、中华鲟(Acipenser sinensis)等洄游性鱼类未采集到,其中,鲢、鳙、日本鳗鲡近年来均有巢湖管理局组织的人工增殖放流,很可能为人工增殖放流种群,青鱼和草鱼可能由附近养殖场逃逸或民众放生,巢湖中洄游性鱼类自然种群可能已基本消失。此外,赤眼鳟(Squaliobarbus curriculus)、马口鱼(Opsariichthys bidens)等喜清澈溪流的鱼类和
外来鱼类入侵可能会成为巢湖鱼类新的危机。本次调查新记录了外来种须鳗

巢湖鱼类群落结构在不同水域和季节间均表现出一定变化。非度量多维标尺排序分析结果显示,在不同采样点及不同采样时间鱼类物种组成结构均出现明显的分离。研究[24, 27]发现,湖区捕捞作业强度对鱼类群落结构的变化具有明显的影响作用。对于巢湖而言,鱼类区系空间结构的差异与人为捕捞强度和自然环境空间差异性具有紧密的关系。一方面,捕捞期内对全湖的捕捞强度并不均匀,对部分鱼类聚集区域会增大捕捞强度,例如:捕捞刀鲚和太湖新银鱼时,近岸浅水区域和河口区域刀鲚和太湖新银鱼相对丰富,渔民的捕捞强度高,对湖心敞水区捕捞力度相对较低;捕鱼时渔船的轰鸣和网具的搅动也会促进不同区域间鱼类的交换,从而降低区域间鱼类群落结构的差异。另一方面,湖区支流水环境的变化将通过影响湖区水质进一步改变湖区鱼类群落结构[24],在巢湖中存在类似的现象。巢湖流域独特的自然环境特征将整合集水区分为不同的功能区划[6],不同功能区划内土地利用类型及自然景观环境的差异性导致入湖支流水环境的变化,从而改变了巢湖不同区域间水体环境的差异性,进而影响鱼类群落的组成结构。
鱼类群落结构时间格局下的差异性受群落内部调节和外部环境因素的共同影响。首先,不同季节间鱼类繁殖群体的变化、鱼类在湖区内的迁移活动等均会导致不同鱼类群落结构的季节性差异[28-29];其次,捕捞、水文因子、饵料生物等外部环境的变化也是导致鱼类群落结构季节性差异的重要原因。全面禁捕前,巢湖实行分阶段开捕,6月底—12月的捕捞期细分为虾类捕捞期(6月底—7月,网具为地笼网和虾笼网)、小型鱼类捕捞期(8月中旬—9月,网具为拖网和推网)、大中型鱼类捕捞期(10月中旬—12月,捕捞工具为刺网)等不同时期,渔民分阶段对鱼(虾)类进行选择性捕捞,例如小型鱼类捕捞期,主要捕捞对象为刀鲚、太湖新银鱼等小型鱼类,根据我们的调查,许多渔船一晚捕捞的刀鲚和太湖新银鱼超过3 000 kg,刀鲚、太湖新银鱼等小型鱼类的资源量在这一时期急剧下降,而在大中型鱼类捕捞期,只允许刺网捕捞,鲢、鳙、鲤、鲌类等体型较大鱼类的资源量会在这一时期急剧下降,捕捞会增强巢湖季节间鱼类群落结构的差异性。另一方面,鱼类物种组成结构差异与环境因子的季节性变动具有十分重要的联系[19, 24],不同采样季节水温、水体理化和饵料生物资源均存在差异,从而导致不同季节鱼类群落在物种组成结构上的分离,与鄱阳湖、太湖等水域研究[22, 30]的结果一致。
3.3 鱼类多样性变化调查表明,巢湖鱼类多样性较低,优势鱼类为刀鲚和太湖新银鱼两种小型鱼类,渔获物数量占比达96.84%,表明巢湖鱼类低龄化、小型化趋势严重。各样点不同时期的Shannon-Wiener多样性指数(0.56~1.09)、丰富度指数(0.19~0.32)、均匀度指数(1.45~3.82)均较低,多样性指数低于MAGURRAN[31]提出的多样性一般范围(1.5~3.5),低于长江中下游的洞庭湖[16]、洪泽湖[24],与太湖[23]较为接近。江湖阻隔、过度捕捞是巢湖鱼类多样性低的重要原因。与此同时,巢湖鱼类生物多样性指数的季节性差异与鱼类群落自身生活史特征和栖息环境的差异性有关。首先,对于泛滥平原湖泊,在春夏洪泛季节,江河中的鱼类随索饵和生殖洄游进入湖泊是其鱼类多样性和资源量的重要补充,河流的季节性水位波动在江湖连通的支持下驱动了通江湖泊鱼类群落的季节波动[32-33]。其次,鱼类物种多样性同水生植物的多样性呈正相关,水生植物丰富的区域,空间异质性及饵料生物的多样性高,鱼类多样性高[34-36]。巢湖地处长江中下游的泛滥平原,巢湖闸和裕溪闸的建立使巢湖鱼类洄游通道受阻,虽然修建有鱼道,但过鱼效果不明显,鱼类多样性未得到有效补充。建闸后,巢湖水位大幅提高,加上沿岸堤防工程的建设,巢湖挺水植物严重衰退,鱼类栖息的空间异质性及饵料生物的多样性降低,部分依靠水生植物产卵的鱼类繁殖场所大幅减少[5, 8]。巢湖周边有约1 800户渔民,6月底—12月为捕捞期,捕捞期内湖面上通常是千帆竞行,刺网、地笼、拖网、推网等网具均会使用,捕捞压力大,生命周期较长、营养级较高、综合繁殖力相对较低的大型鱼类受捕捞的影响会明显大于营养层次较低、综合繁殖力较高的小型鱼类,从而导致鱼类种群小型化加剧[4-37]。
巢湖从2020年1月1日开始实施为期10年的全面禁渔,捕捞压力骤然消失,位于食物链上游的鲌类、鳜等掠食性鱼类较少,刀鲚、太湖新银鱼等繁殖力强、生命周期短的小型鱼类优势地位可能会提高,增加鱼类群落的不稳定性,需要对巢湖进行持续监测。随着引江济淮工程的实施,巢湖即将恢复与长江的连通,并与淮河连通,需加强“引江济淮”生态调度研究,充分发挥江湖连通的生态作用,避免调水引起更多生物入侵。
| [1] |
马荣华, 杨桂山, 段洪涛, 等. 中国湖泊的数量、面积与空间分布[J]. 中国科学: 地球科学, 2011, 41(3): 394-401. MA R H, YANG G S, DUAN H T, et al. China's lakes at present: number, area and spatial distribution[J]. Science China Earth Sciences, 2011, 41(3): 394-401. |
| [2] |
王洪铸, 刘学勤, 王海军. 长江河流-泛滥平原生态系统面临的威胁与整体保护对策[J]. 水生生物学报, 2019, 43(s1): 157-182. WANG H Z, LIU X Q, WANG H J. The Yangtze river-floodplain ecosystem: multiple threats and holistic conservation[J]. Acta Hydrobiologica Sinica, 2019, 43(s1): 157-182. DOI:10.7541/2019.178 |
| [3] |
LÉVÊQUE C, OBERDORFF T, PAUGY D, et al. Global diversity of fish (Pisces) in freshwater[J]. Hydrobiologia, 2008, 595(1): 545-567. DOI:10.1007/s10750-007-9034-0 |
| [4] |
WOODCOCK P, O'LEARY B C, KAISER M J, et al. Your evidence or mine? Systematic evaluation of reviews of marine protected area effectiveness[J]. Fish and Fisheries, 2017, 18(4): 668-681. DOI:10.1111/faf.12196 |
| [5] |
郝贝贝, 吴昊平, 刘文治, 等. 巢湖湖滨带植被特征及其退化原因分析研究[J]. 环境科学与管理, 2013, 38(6): 59-65. HAO B B, WU H P, LIU W Z, et al. Vegetation features and degradation causes in Chaohu lakeshore zone[J]. Environmental Science and Management, 2013, 38(6): 59-65. DOI:10.3969/j.issn.1673-1212.2013.06.013 |
| [6] |
钱红, 严云志, 储玲, 等. 巢湖流域河流鱼类群落的时空分布[J]. 长江流域资源与环境, 2016, 25(2): 257-264. QIAN H, YAN Y Z, CHU L, et al. Spatial and temporal patterns of fish assemblages in the rivers of Chaohu basin[J]. Resources and Environment in the Yangtze Basin, 2016, 25(2): 257-264. |
| [7] |
安徽省人民政府水产局区划办公室. 安徽省长江水系重点水域渔业资源调查报告汇编[M]. 合肥: 安徽省人民政府水产局区划办公室, 1985: 1-35. Division Office of Fisheries Bureau, People's Government of Anhui Province. Compilation of investigation reports on fishery resources in key water systems of the Yangtze River in Anhui Province[M]. Hefei: Division Office of Fisheries Bureau, People's Government of Anhui Province, 1985: 1-35. |
| [8] |
过龙根, 谢平, 倪乐意, 等. 巢湖渔业资源现状及其对水体富营养化的响应研究[J]. 水生生物学报, 2007, 31(5): 700-705. GUO L G, XIE P, NI L Y, et al. The status of fishery resources of Lake Chaohu and its response to eutrophication[J]. Acta Hydrobiologica Sinica, 2007, 31(5): 700-705. DOI:10.3321/j.issn:1000-3207.2007.05.015 |
| [9] |
汪顺丽, 张宏才, 魏荷芬, 等. 巢湖东半湖富营养化现状的研究[J]. 生物学杂志, 2016, 33(2): 13-17. WANG S L, ZHANG H C, WEI H F, et al. Eutrophication current situation and research of the east-half of Chaohu Lake[J]. Journal of Biology, 2016, 33(2): 13-17. DOI:10.3969/j.issn.2095-1736.2016.02.013 |
| [10] |
王岐山. 巢湖鱼类区系研究[J]. 安徽大学学报(自然科学版), 1987, 11(2): 70-78. WANG Q S. Study on ichthyologlcal fauna of Chao Lake[J]. Journal of Anhui University (Natural Science), 1987, 11(2): 70-78. |
| [11] |
王书航, 姜霞, 金相灿. 巢湖入湖河流分类及污染特征分析[J]. 环境科学, 2011, 32(10): 2834-2839. WANG S H, JIANG X, JIN X C. Classification and pollution characteristic analysis for inflow rivers of Chaohu Lake[J]. Environmental Science, 2011, 32(10): 2834-2839. DOI:10.13227/j.hjkx.2011.10.024 |
| [12] |
陈宜瑜. 中国动物志硬骨鱼纲鲤形目(中卷)[M]. 北京: 科学出版社, 1998. CHEN Y Y. Fauna sinica, osterichthyes, cypriniformes Ⅱ[M]. Beijing: Science Press, 1998. |
| [13] |
朱松泉. 中国淡水鱼类检索[M]. 南京: 江苏科学技术出版社, 1995. ZHU S Q. The synopsis of freshwater fishes of China[M]. Nanjing: Jiangsu Science and Technology Publishing House, 1995. |
| [14] |
倪勇, 伍汉霖. 江苏鱼类志[M]. 北京: 中国农业出版社, 2006. NI Y, WU H L. Fishes of Jiangsu province[M]. Beijing: China Agriculture Press, 2006. |
| [15] |
浙江动物志编辑委员会. 浙江动物志: 淡水鱼类[M]. 杭州: 浙江科学技术出版社, 1991. Zhejiang Zoological Editorial Committee. Fauna of Zhejiang freshwater fishes[M]. Hangzhou: Zhejiang Science and Technology Publishing House, 1991. |
| [16] |
蒋忠冠, 曹亮, 张鹗. 洞庭湖鱼类的群落结构及其时空动态[J]. 水生生物学报, 2019, 43(s1): 42-48. JIANG Z G, CAO L, ZHANG E. Spatio-temporal variations of fish assemblages in the Dongting Lake[J]. Acta Hydrobiologica Sinica, 2019, 43(s1): 42-48. DOI:10.7541/2019.165 |
| [17] |
王雪辉, 杜飞雁, 邱永松, 等. 1980—2007年大亚湾鱼类物种多样性、区系特征和数量变化[J]. 应用生态学报, 2010, 21(9): 2403-2410. WANG X H, DU F Y, QIU Y S, et al. Variations of fish species diversity, faunal assemblage, and abundances in Daya Bay in 1980-2007[J]. Chinese Journal of Applied Ecology, 2010, 21(9): 2403-2410. |
| [18] |
PEASE A A, TAYLOR J M, WINEMILLER K O, et al. Multiscale environmental influences on fish assemblage structure in central Texas streams[J]. Transactions of the American Fisheries Society, 2011, 140(5): 1409-1427. DOI:10.1080/00028487.2011.623994 |
| [19] |
郝雅宾, 刘金殿, 郭爱环, 等. 千岛湖国家级水产种质资源保护区(梓桐核心区)鱼类群落结构[J]. 上海海洋大学学报, 2019, 28(4): 587-596. HAO Y B, LIU J D, GUO A H, et al. Fish community structure in Zitong core area in Qiandao Lake national aquatic genetic resources conservation area[J]. Journal of Shanghai Ocean University, 2019, 28(4): 587-596. |
| [20] |
ANDERSON M J. A new method for non-parametric multivariate analysis of variance[J]. Austral Ecology, 2001, 26(1): 32-46. |
| [21] |
安徽省革命委员会科技局, 安徽省革命委员会农林局. 安徽长江主要经济鱼类资源调查报告汇编[M]. 合肥: 安徽省科技局, 安徽省农林局, 1974. Science and Technology Bureau of Anhui Revolutionary Committee, Agriculture and Forestry Bureau of Anhui Revolutionary Committee. Compilation of investigation reports on fishery resources of the Yangtze River in Anhui Province[M]. Hefei: Anhui Science and Technology Bureau, Anhui agriculture and Forestry Bureau, 1974. |
| [22] |
杨少荣, 黎明政, 朱其广, 等. 鄱阳湖鱼类群落结构及其时空动态[J]. 长江流域资源与环境, 2015, 24(1): 54-64. YANG S R, LI M Z, ZHU Q G, et al. Spatial and temporal variations of fish assemblages in Poyanghu Lake[J]. Resources and Environment in the Yangtze Basin, 2015, 24(1): 54-64. |
| [23] |
毛志刚, 谷孝鸿, 曾庆飞, 等. 太湖鱼类群落结构及多样性[J]. 生态学杂志, 2011, 30(12): 2836-2842. MAO Z G, GU X H, ZENG Q F, et al. Community structure and diversity of fish in Lake Taihu[J]. Chinese Journal of Ecology, 2011, 30(12): 2836-2842. |
| [24] |
毛志刚, 谷孝鸿, 龚志军, 等. 洪泽湖鱼类群落结构及其资源变化[J]. 湖泊科学, 2019, 31(4): 1109-1119. MAO Z G, GU X H, GONG Z J, et al. The structure of fish community and changes of fishery resources in Lake Hongze[J]. Journal of Lake Sciences, 2019, 31(4): 1109-1119. |
| [25] |
中国科学院中国动物志编辑委员会, 伍汉霖, 钟俊生. 中国动物志, 硬骨鱼纲, 鲈形目(五), 虾虎鱼亚目[M]. 北京: 科学出版社, 2008. Editorial Committee of Zoology of China, Chinese Academy of Sciences, WU H L, ZHONG J S. Fauna sinica, ostichthyes, perciformes (Ⅴ), gobioidei[M]. Beijing: Science Press, 2008. |
| [26] |
LIANG Y Y, FANG T, LI J, et al. Age, growth and reproductive traits of invasive goby Taenioides cirratus in the Chaohu Lake, China[J]. Journal of Applied Ichthyology, 2020, 36(2): 219-226. |
| [27] |
刘恩生. 太湖鱼类群落变化规律、机制及其对环境影响分析[J]. 水生态学杂志, 2009, 2(4): 8-14. LIU E S. Changes of fish community, the mechanism of changes, and the effects of the changes on environment in Taihu Lake[J]. Journal of Hydroecology, 2009, 2(4): 8-14. |
| [28] |
HE D K, KANG Z J, TAO J, et al. Hydrologic connectivity driven natural stream fish assemblages in mountain streams in the Yangtze River basin: implications for stream fish conservation in monsoonal East Asia[J]. Hydrobiologia, 2017, 785(1): 185-206. |
| [29] |
MULLEN J A, BRAMBLETT R G, GUY C S, et al. Determinants of fish assemblage structure in Northwestern Great Plains streams[J]. Transactions of the American Fisheries Society, 2011, 140(2): 271-281. |
| [30] |
李其芳, 严云志, 储玲, 等. 太湖流域河流鱼类群落的时空分布[J]. 湖泊科学, 2016, 28(6): 1371-1380. LI Q F, YAN Y Z, CHU L, et al. Spatial and temporal patterns of stream fish assemblages within Taihu basin[J]. Journal of Lake Sciences, 2016, 28(6): 1371-1380. |
| [31] |
MAGURRAN A E. Ecological diversity and its measurement[M]. Princeton: Princeton University Press, 1988.
|
| [32] |
FERNANDES I M, HENRIQUES-SILVA R, PENHA J, et al. Spatiotemporal dynamics in a seasonal metacommunity structure is predictable: the case of floodplain-fish communities[J]. Ecography, 2014, 37(5): 464-475. |
| [33] |
RÖPKE C P, AMADIO S A, WINEMILLER K O, et al. Seasonal dynamics of the fish assemblage in a floodplain lake at the confluence of the Negro and Amazon Rivers[J]. Journal of Fish Biology, 2016, 89(1): 194-212. |
| [34] |
DIBBLE E D, PELICICE F M. Influence of aquatic plant-specific habitat on an assemblage of small neotropical floodplain fishes[J]. Ecology of Freshwater Fish, 2010, 19(3): 381-389. |
| [35] |
DUFRÊNE M, LEGENDRE P. Species assemblages and indicator species: the need for a flexible asymmetrical approach[J]. Ecological Monographs, 1997, 67(3): 345-366. |
| [36] |
田佳丽, 王银平, 李佩杰, 等. 长江近口段近岸段鱼类群落多样性现状[J]. 上海海洋大学学报, 2021, 30(2): 320-330. TIAN J L, WANG Y P, LI P J, et al. Species diversity of fish in the section near the Yangtze River estuary[J]. Journal of Shanghai Ocean University, 2021, 30(2): 320-330. |
| [37] |
PINSKY M L, PALUMBI S R. Meta-analysis reveals lower genetic diversity in overfished populations[J]. Molecular Ecology, 2014, 23(1): 29-39. |
 ,
LU Wenxuan1,
YANG Kun1,
LI Jing1,
ZHAO Xiuxia1,
FANG Ting1,
GAO Na1,
CHEN Kang2
,
LU Wenxuan1,
YANG Kun1,
LI Jing1,
ZHAO Xiuxia1,
FANG Ting1,
GAO Na1,
CHEN Kang2 ,
YIN Feng3
,
YIN Feng3
2. Key Laboratory of Freshwater Biodiversity Conservation, Ministry of Agriculture and Rural Affairs, Yangtze River Fisheries Research Institute, Chinese Academy of Fishery Sciences, Wuhan 430223, Hubei, China;
3. Fishery Management Station, Chaohu Administration, Hefei 238001, Anhui, China