 2022,
Vol. 31
2022,
Vol. 31



鱼类作为水生生物多样性的组成部分,对环境的变化有着重要的指示作用。鱼类群落对自然栖息地具有不同的生态、形态和行为适应性,是评价水生态系统的有效指标[1-2]。全球气候的变化和人类活动不同程度地影响着河流和湖泊系统中生物的栖息地,如湖泊面积萎缩、河流断流或干涸、人工筑坝和水库修建等导致水文连通性的改变,可能会对现有生物造成更大的生存压力,威胁到现有生物种群的长期生存能力[3-4]。淡水鱼类多样性正迅速成为全球生物多样性的组成部分,更好地了解鱼类群落如何在空间和时间上对环境因素的反应,对于制定有效的保护策略非常重要[4]。
白洋淀位于河北省雄安新区境内,是华北平原最大的内陆淡水湖泊,对华北地区生态环境有着重大影响,素有“华北之肾”之称。白洋淀鱼类是华北地区物种多样性的重要组成部分。关于白洋淀鱼类资源调查研究始于20世纪30年代,1960年郑葆珊等[5]所著《白洋淀鱼类》一书较为详尽地记录了白洋淀54种鱼类,此时白洋淀鱼类的种类和数量极为丰富。但自20世纪中期以后,白洋淀上游修建水利工程、水量明显减少、淀区面积萎缩、水体污染事件频频发生、生态环境破坏严重[6],水量、水质、水温等因素的改变对生境中的生物尤其是鱼类影响较大,致使鱼类资源数量、区系组成发生重大变化。如王所安等[7]在1975—1976年间继白洋淀上游修库拦洪后的调查显示,白洋淀减少了沿海溯河入淀和上游河流产卵洄游入淀的鱼类,并且鱼产量明显下降。为恢复白洋淀生态功能,政府开展了“引黄济淀”、“南水北调”等一系列调水补水工程,白洋淀生态环境得到了改善,但白洋淀鱼类资源恢复并不乐观,对白洋淀鱼类资源定期性的调查显示[8-12],白洋淀鱼类物种数和个体数仍维持在较低水平,物种数不到40种。然而,在2019—2020年的调查中,闫丽等[13]记录了白洋淀鱼类44种,显示白洋淀鱼类资源得到了一定的恢复,但与最初的54种相比,仍缺少沿海溯河入淀和上游河流产卵洄游性鱼类。
于2019—2020年开展对白洋淀鱼类资源的调查,旨在了解淀区内现有的鱼类群落结构特征,分析鱼类群落结构与环境因子的关系,为今后白洋淀鱼类资源的管理、保护和生态修复提供科学依据,也为白洋淀鱼类多样性研究积累基础资料。
1 材料与方法 1.1 样品采集2019年5月、8月、10月和2020年8月、10月(2020年5月由于疫情影响未调查)共5次分3个季度对白洋淀淀区30个位点进行鱼类资源调查(图 1)。每个位点均设置2条单层丝网和2条地笼。丝网规格为1 m×30 m和1.5 m×45 m,网目分别为3 cm和7 cm;地笼规格为35 cm×30 cm和25 cm×20 cm,网目为1 cm。丝网和地笼设置时间约为14 h,即下午17:00~19:00下网至次日5:00~7:00起网。依据最新鱼类学专著或研究论文对采集的渔获物进行分类鉴定及生态类型分类[5, 14-18]。对现场采集到新鲜样本进行体长、体质量等常规生物学处理,体质量精确到g,体长精确到mm,并统计每个位点捕获鱼类的物种数和个体数。对于现场无法鉴定的物种经10%的甲醛溶液固定,并带回实验室进一步分类鉴定。

|
图 1 白洋淀位点示意图 Fig. 1 Collection sites in Baiyangdian Lake |
对30个位点使用水质测定仪测定水温(T)、溶解氧(DO)、生化需氧量(BOD5)、pH、透明度(SD)、叶绿素a(Chl.a)、总磷(TP)、总氮(TN)等环境因子,使用水深测定仪现场测定水深。
1.2 数据处理与分析通过Excel 2010对每个位点每网的物种数(S)、个体数(N)、体质量(W)等原始数据进行统计。利用Primer 5.0软件计算Margalef种类丰富度指数(D)[19]、Shannon-Wiener多样性指数(H′)[20]和Pielou均匀度指数(J′)[21],利用Excel 2010计算Pinkas相对重要性指数[22],丰度生物量比较曲线(ABC曲线)[23],公式如下:
 (1)
(1)
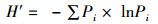 (2)
(2)
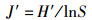 (3)
(3)
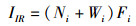 (4)
(4)
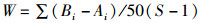 (5)
(5)
式中:S为某个位点渔获物的种类数,N为某个位点渔获物总的个体数,Pi为某个位点第i种鱼类生物的个体数占总个体数的比例,Ni为渔获物第i种鱼的尾数占总尾数的百分数,%;Wi为第i种鱼的生物量占总生物量的百分数,%;Fi为第i种鱼在所有采样次数中出现的频率,定义IIR < 1为稀有种;1≤IIR < 10为一般种;10≤IIR < 100为常见种;100≤IIR < 500为主要种;IIR≥500为优势种;Ai为某物种相对应的丰度累积百分比,%;Bi为某物种相对应的生物量累积百分比,%;W值作为ABC曲线定量的判断标准。
为满足数据的正态性和方差齐性,对鱼类丰度数据分析时,对原始数据进行log2(x+1)对数转换,然后运用SPSS 22.0进行单因素方差分析(One-way ANOVA)检验不同季节及位点的鱼类多样性差异。如鱼类多样性显著性差异,进一步进行SNK(Student-Newman-Keuls)多重比较分析不同季节间及位点的鱼类多样性变化[24]。
在Primer 5.0软件中,以每个季节鱼类物种数量为原始矩阵,进行log2(x+1)转换后,运用单因素分析(ANOSIM)检验不同季节对鱼类种群结构组成的影响[25],根据Global R值来确定鱼类种群的离散程度:当R≤0.25时,鱼类群落重叠无分离现象;当0.25 < R≤0.5,群落存在一定程度的重叠但仍可分离;0.5 < R≤0.75,群落存在一定程度重叠但分离明显;R>0.75,群落完全分离。运用非参数多变量排序(non-metric multidimensional scaling,NMS)构建鱼类群落结构的二维双标图,运用相似性百分比分析(similarity percentages,SIMPER)[26]确定维持不同季节间结构相似性的关键物种。
利用Canoco 5.0[27]软件对2019年3个季度鱼类物种组成和环境因子进行去趋势对应分析(detrended correspondence analysis,DCA),根据分析结果中各排序轴的大小选择线性模型(redundancy analysis,RDA)或单峰模型(canonical correspondence analysis,CCA)分析鱼类物种和环境因子的相关性。如果排序轴小于3,选择RDA分析;介于3和4之间,RDA分析和CCA分析均可;排序轴大于4,选择CCA分析。在RDA分析时,由于原始鱼类物种数据存在许多0值,首先对原始数据进行Hellinger转化,用转化后的鱼类物种数据作为响应变量,原始环境因子数据作为解释变量,对鱼类数据和环境因子进行RDA分析。
2 结果 2.1 鱼类组成、生态类型及优势种2019—2020年在白洋淀共捕获鱼类30种,隶属4目12科(表 1)。其中,鲤形目最多,为2科17种,占56.67%;其次为鲈形目,为6科9种,占30.00%;鲇形目为2科2种,占6.67%;合鳃鱼目2科2种,占6.67%。

|
表 1 白洋淀鱼类组成、各月份相对重要指数(IRI)及生态类型 Tab.1 Fish composition, relative importance index (IRI) of each month and ecological types in Baiyangdian Lake |
鱼类生态类型划分(表 1):按生活习性划分,定居型鱼类(S)13种,占43.33%;河流型鱼类(R)13种,占43.33%;河湖洄游性鱼类(RL)4种占13.33%。按食性划分,肉食性鱼类(Car)11种,占36.67%;植食性鱼类(Her)2种,占6.67%;杂食性鱼类(Omn)最多,为17种,占56.67%。按栖息水层划分,中上层鱼类(U)6种,占20.00%;中下层鱼类(Lo)11种,占36.67%;底栖鱼类(De)最多,有13种,占43.33%。按产卵类型划分,产黏性卵鱼类(V)10种,占33.33%;产沉性卵鱼类(D)有7种,占23.33%;产漂流性卵鱼类(Dr)有10种,占33.33%;喜贝性卵鱼类(Sp)有3种,占10.00%。
根据相对重要指数IIR≥500为优势种,白洋淀鱼类优势种共9种(表 1)。5次调查中,各月优势种不同,其中,2019年5月优势种有鲫(Carassius auratus)、红鳍鲌(Chanodichthys erythropterus)、麦穗鱼(Pseudorasbora parva)、子陵吻虾虎鱼(Rhinogobiu giurinus);2019年8月白洋淀优势种有鲫、

单因素方差分析结果显示(表 2),白洋淀鱼类在不同位点间鱼类的个体数(N)、物种数(S)、Margalef种类丰富度指数(D)、Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J′)均无显著差异(P>0.05);不同位点的鱼类个体数、物种数、丰富度指数、多样性指数、均匀度指数呈显著的季节差异(P < 0.05),进一步使用SNK多重比较分析,结果显示除均匀度指数外,个体数、物种数、丰富度指数、多样性指数均为2019年5月最高(图 2)。2019年5月,多样性指数各位点为0.82~2.03(平均为1.63);丰富度指数各位点为1.03~2.82(平均为2.03);均匀度指数各位点为0.34~0.84(平均为0.67)。

|
表 2 基于单因素方差分析检验(F值)白洋淀鱼类多样性指数不同位点及季节的变化 Tab.2 Changes of fish diversity index at different loci and in different seasons in Baiyangdian Lake on one-way ANOVA test (F value) |

|
图 2 不同季节间白洋淀鱼类多样性指数差异 Fig. 2 Differences of fish diversity index in different seasons in Baiyangdian Lake |
对白洋淀鱼类群落进行非参数多变量(NMS)排序分析和单因子相似性分析(ANOSIM)结果显示(图 3),不同季节间鱼类群落组成既重叠又呈现一定的分离状态(Global R=0.524)。相似性百分比分析SIMPER显示(表 3),2019年5月各位点鱼类平均相似性为61.31%,主要贡献种为鲫、麦穗鱼、子陵吻虾虎鱼、小黄黝鱼、红鳍鲌,累积贡献率为79.50%;2019年8月各位点鱼类平均相似性为65.62%,主要贡献种为鲫、红鳍鲌、

|
图 3 白洋淀鱼类群落不同季节分布NMS图 Fig. 3 Non-metric multi-dimensional scaling (NMS) analysis of fish community in Baiyangdian Lake in different seasons |

|
表 3 白洋淀不同季节的主要贡献种相似性百分比分析(贡献度大于5%) Tab.3 The main contribution species of Baiyangdian Lake in different seasons and similarity percentage analysis (contribution rate greater than 5%) |
ABC曲线分析结果显示(图 4),2019年5月、8月、10月鱼类丰度优势度在生物量优势度之下,W值分别为0.092、0.032、0.105,鱼类群落受轻度干扰的状态;2020年8月、10月鱼类丰度优势度在生物量优势度之上,W值分别为-0.039、-0.022,鱼类群落受中度干扰状态。

|
图 4 白洋淀鱼类群落不同季节ABC曲线 Fig. 4 Abundance-biomass comparison curves of fish community in Baiyangdian Lake in different seasons |
DCA排序结果显示4个排序轴长度均小于3(表 4),其中第一轴长为2.36,物种累积解释变量为18.83%,物种和环境累积解释变量为18.17%,物种和环境与排序轴相关性最高为75.49%。白洋淀鱼类群落与环境因子分析适用于线性模型RDA分析。

|
表 4 白洋淀鱼类群落DCA分析 Tab.4 Detrended correspondence analysis of fish community in Baiyangdian Lake |
RDA分析中第一轴和第二轴为主成分轴(表 5),第一轴解释变化率为22.01%,第一轴和第二轴累积解释变化率为25.71%,物种和环境与前两轴相关性分别为68.94%和51.75%。白洋淀鱼类群落与环境因子RDA排序图结果显示(图 5),在第一轴上(Axis 1),对鱼类群落组成影响最大的环境变量为叶绿素a和溶解氧,蒙特卡洛检验叶绿素a与第一轴的正相关性较高(F=7.8,P=0.002),以鲫、鲇(Silurus asotus)、麦穗鱼、棒花鱼(Abbottina rivularis)等具有较强耐受性鱼类为主;在第二轴上(Axis 2),水温对鱼类群落组成影响较大,蒙特卡洛检验呈正相关性(F=12.5,P=0.002),以

|
表 5 白洋淀鱼类群落RDA分析 Tab.5 Redundancy analysis of fish community in Baiyangdian Lake |

|
Tem.水温;Dep.水深;DO.溶解氧;Chl.a.叶绿素a;SD.透明度;BOD5.生化需氧量;TN.总氮;TP.总磷;sp1.鲫;sp2.鲤;sp3.鲢;sp4.鳙;sp5.;sp6.似;sp7.红鳍鲌;sp8.团头鲂;sp9.兴凯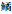 ;sp10.中华鳑鲏;sp11.高体鳑鲏;sp12.草鱼;sp13.麦穗鱼;sp14.黑鳍鳈;sp15.棒花鱼;sp16.泥鳅;sp17.大鳞副泥鳅;sp18.刺鳅;sp19.黄鳝;sp20.子陵吻虾虎鱼;sp21.林氏吻虾虎鱼;sp22.波氏吻虾虎鱼;sp23.福岛虾虎鱼;sp24.小黄黝鱼;sp25.圆尾斗鱼;sp26.鳜;sp27.奥利亚罗非鱼;sp28.乌鳢;sp29.黄颡鱼;sp30.鲇。 Tem.Temperature; Dep.Depth; DO.Dissolved Oxygen; Chl.a.Chlorophyll a; SD.Spectral distribution; BOD5.Biochemical Oxygen Demand; TN.Total Nitrogen; TP.Total Phosphorus; sp1.Carassius auratus; sp2.Cyprinus carpio; sp3.Hypophthalmichthys molitrix; sp4.Hypophthalmichthys nobilis; sp5.Hemiculter leucisculus; sp6.Toxabramis swinhonis; sp7.Chanodichthys erythropterus; sp8.Megalobrama amblycephala; sp9.Acheilognathus chankaensis; sp10.Rhodeus sinensis; sp11.Rhodeus ocellatus; sp12.Ctenopharyngodon idella; sp13.Pseudorasbora parva; sp14.Sarcocheilichthys nigripinnis; sp15.Abbottina rivularis; sp16.Misgurnus anguillicaudatus; sp17.Misgurnus dabryanus; sp18.Sinobdella sinensis; sp19.Monopterus albus; sp20.Rhinogobius giurinus; sp21.Rhinogobius lindbergi; sp22.Rhinogobius cliffordpopei; sp23.Rhinogobius fukushimai; sp24.Micropercops swinhonis; sp25.Macropodus ocellatus; sp26.Siniperca chuatsi; sp27.Oreochromis aureus; sp28.Channa argus; sp29.Tachysurus fulvidraco; sp30.Silurus asotus.
图 5 白洋淀鱼类群落与环境因子RDA分析
Fig. 5 Redundancy analysis of fish community and environmental factors in Baiyangdian Lake ;sp10.中华鳑鲏;sp11.高体鳑鲏;sp12.草鱼;sp13.麦穗鱼;sp14.黑鳍鳈;sp15.棒花鱼;sp16.泥鳅;sp17.大鳞副泥鳅;sp18.刺鳅;sp19.黄鳝;sp20.子陵吻虾虎鱼;sp21.林氏吻虾虎鱼;sp22.波氏吻虾虎鱼;sp23.福岛虾虎鱼;sp24.小黄黝鱼;sp25.圆尾斗鱼;sp26.鳜;sp27.奥利亚罗非鱼;sp28.乌鳢;sp29.黄颡鱼;sp30.鲇。 Tem.Temperature; Dep.Depth; DO.Dissolved Oxygen; Chl.a.Chlorophyll a; SD.Spectral distribution; BOD5.Biochemical Oxygen Demand; TN.Total Nitrogen; TP.Total Phosphorus; sp1.Carassius auratus; sp2.Cyprinus carpio; sp3.Hypophthalmichthys molitrix; sp4.Hypophthalmichthys nobilis; sp5.Hemiculter leucisculus; sp6.Toxabramis swinhonis; sp7.Chanodichthys erythropterus; sp8.Megalobrama amblycephala; sp9.Acheilognathus chankaensis; sp10.Rhodeus sinensis; sp11.Rhodeus ocellatus; sp12.Ctenopharyngodon idella; sp13.Pseudorasbora parva; sp14.Sarcocheilichthys nigripinnis; sp15.Abbottina rivularis; sp16.Misgurnus anguillicaudatus; sp17.Misgurnus dabryanus; sp18.Sinobdella sinensis; sp19.Monopterus albus; sp20.Rhinogobius giurinus; sp21.Rhinogobius lindbergi; sp22.Rhinogobius cliffordpopei; sp23.Rhinogobius fukushimai; sp24.Micropercops swinhonis; sp25.Macropodus ocellatus; sp26.Siniperca chuatsi; sp27.Oreochromis aureus; sp28.Channa argus; sp29.Tachysurus fulvidraco; sp30.Silurus asotus.
图 5 白洋淀鱼类群落与环境因子RDA分析
Fig. 5 Redundancy analysis of fish community and environmental factors in Baiyangdian Lake
|
白洋淀位于古北区的华北亚区[28],兴起于第三纪,成于第四纪[29]。白洋淀鱼类特点是以江河平原鱼类为主,因其地理位置临近渤海湾,因而有部分海产鱼类可溯河进入淀区。白洋淀鱼类代表性的资料《白洋淀鱼类》记录了54种鱼类,隶属11目17科50属[5]。国内其他调查资料显示,水库、大坝等涉水工程的修建,导致洄游通道阻塞,是造成洄游性鱼类资源衰退甚至灭绝的主要原因[30]。20世纪60年代在入淀河系上游相继建库修坝,白洋淀鱼类减少了沿海溯河入淀和上游河流产卵洄游入淀的种类,如日本鳗鲡(Anguilla japonica)、龟

|
表 附表 白洋淀鱼类资源历史变化 Tab.附表 Historical changes of fish resources in Baiyangdian Lake |

2019—2020年白洋淀鱼类优势种有鲫、红鳍鲌、麦穗鱼,这与1976年的调查结果一致[7],与20世纪60年代初相比,白洋淀鱼类以耐低氧的种类为主。除上述优势种外,经济价值低的小型鱼类,如
多样性指数(H′)和均匀度指数(J′)是群落结构稳定的重要评价指标,当物种丰富度越高,个体数分布越均匀,群落结构越稳定,多样性指数和均匀度指数则较大,反之,物种受到环境的胁迫,多样性指数和均匀度指数则低[36-37]。本次研究中,白洋淀鱼类个体数、物种数、丰富度指数、多样性指数和均匀度指数均为2019年5月最高,这与其他研究结果类似[27, 37],通常春节鱼类多样性指数较高,可能原因是春节为鱼类繁殖季节,水位也较其他季节低,鱼类多在浅水区活动,容易被地笼和刺网等捕捞渔具捕获。白洋淀鱼类多样性指数(平均为1.63)与北方其他水域接近,如济南市黄河流域多样性指数为1.64~1.90[38];与南方水域相比,白洋淀鱼类资源多样性水平明显较低,如东太湖多样性指数为2.14~3.26[27]。原因可能是北方地区鱼类物种多样性原本就低于南方地区[39],在一定程度上北方地区鱼类群落结构较简单,更容易受环境胁迫的影响。
ABC曲线最早被应用于对海洋大型底栖动物群落结构的研究,随后被越来越广泛地用于评价水体中游泳动物或无脊椎动物群落受污染、干扰的程度[23]。一般认为生物量曲线始终在丰度曲线之上,群落趋向稳定的状态,一旦两曲线相交或重叠,说明群落收到干扰,稳定性下降[23]。白洋淀鱼类群落ABC曲线的结果与优势种、鱼类多样性和相似性百分比SIMPER结果大体一致,物种数、丰富度指数、多样性指数2019年高于2020年。2019年5月、8月、10月鲫、红鳍鲌等体型较大的鱼类相对重要指数、百分比贡献度较高,鱼类生物量曲线中生物量优势度略高于丰度优势度;而2020年8月、10月体型较小的麦穗鱼相对重要指数、百分比贡献度较高,鱼类生物量曲线中丰度优势度略高于生物量优势度,这表明白洋淀鱼类资源的捕捞压力过大,体型较大的类群或个体数量减少速度大于体型小的鱼类。
3.2 白洋淀鱼类群落结构与环境因子关系通常鱼类多样性与栖息生境异质性相关,栖息地异质性高,鱼类多样性就高[40]。白洋淀各位点鱼类群落的物种组成、个体数、丰富度指数、多样性指数、均匀度指数均不存在显著差异,这在一定程度上反映白洋淀各区域鱼类群落结构处在一个相对稳定的状态。在不同季节间,白洋淀鱼类的多样性指数、丰富度指数以及均匀度指数均存在差异,这表明白洋淀鱼类群落主要受非生物环境因子影响。相似性分析显示,白洋淀鱼类2019年5月与2020年8月差异性最大,2019年8月和2020年10月差异性最小。鲫、红鳍鲌、麦穗鱼相似性贡献度最高,达60%左右,而分歧种为团头鲂、鳜、黑鳍鳈(Sarcocheilichthys nigripinnis)、刺鳅(Sinobdella aculeatus),表明白洋淀以耐污的小型鱼类为主。
已有研究表明,水温、水深、透明度、溶解氧、pH、叶绿素a、总磷、总氮等环境因子对湖泊鱼类群落结构分布均有一定的影响[40-43]。通常认为水草覆盖度高,透明度高,与之相反总磷、总氮和叶绿素a低的湖泊鱼类多样性和生物量高[41-44]。本研究中,叶绿素a、溶解氧和水温是影响白洋淀鱼类群落结构的主要环境因子。其中以叶绿素a为主,白洋淀鱼类中鲫、麦穗鱼、鲇、黄颡鱼(Tachysurus fulvidraco)等与之相关性强,叶绿素a与浮游植物密度相关,这也符合白洋淀以耐污的杂食性鱼类为主的特点。溶解氧是许多鱼类分布的限制因子,大部分鱼类尤其是肉食性耗氧高的种类喜生活在高溶氧区域[45-46],白洋淀鱼类中鳑鲏鱼类和虾虎鱼类与溶解氧相关性强,该类群通常在较为清澈的水草中分布。在一定范围内,水温升高,鱼类的摄食量和活跃性增强,白洋淀鱼类中
通过对白洋淀鱼类资源调查及其与环境因子关系分析,调查中共采集到4目12科30种鱼类,优势种为鲫、
| [1] |
KARR J R. Assessment of biotic integrity using fish communities[J]. Fisheries, 1981, 6(6): 21-27. DOI:10.1577/1548-8446(1981)006<0021:AOBIUF>2.0.CO;2 |
| [2] |
MCCORMICK F H, PECK D V, LARSEN D P. Comparison of geographic classification schemes for Mid-Atlantic stream fish assemblages[J]. Journal of the North American Benthological Society, 2000, 19(3): 385-404. DOI:10.2307/1468102 |
| [3] |
JACKSON D A, PERES-NETO P R, OLDEN J D. What controls who is where in freshwater fish communities - the roles of biotic, abiotic, and spatial factors[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2001, 58(1): 157-170. |
| [4] |
PEASE A A, TAYLOR J M, WINEMILLER K O, et al. Multiscale environmental influences on fish assemblage structure in central Texas streams[J]. Transactions of the American Fisheries Society, 2011, 140(5): 1409-1427. DOI:10.1080/00028487.2011.623994 |
| [5] |
郑葆珊, 范勤德, 戴定远. 白洋淀鱼类[M]. 石家庄: 河北人民出版社, 1960. ZHENG B S, FAN Q D, DAI D Y. Fish of Baiyangdian lake[M]. Shijiazhuang: Hebei People's Publishing House, 1960. |
| [6] |
王玉蛟, 李桃. 浅析白洋淀生态意义及环境保护与治理[J]. 河北地质, 2017(2): 31-33. WANG Y J, LI T. Analysis of Baiyangdian ecological significance, environmental protection and governance[J]. Hebei Geology, 2017(2): 31-33. |
| [7] |
王所安, 顾景龄. 白洋淀环境变化对鱼类组成和生态的影响[J]. 动物学杂志, 1981(4): 8-11. WANG S A, GU J L. Effects of environmental changes on fish composition and ecology in Baiyangdian Lake[J]. Chinese Journal of Zoology, 1981(4): 8-11. DOI:10.13859/j.cjz.1981.04.004 |
| [8] |
曹玉萍. 白洋淀重新蓄水后鱼类资源状况初报[J]. 淡水渔业, 1991(5): 20-22. CAO Y P. Preliminary report on fish resources after re-impoundment of Baiyangdian Lake[J]. Freshwater Fisheries, 1991(5): 20-22. |
| [9] |
韩希福, 王所安, 曹玉萍, 等. 白洋淀重新蓄水后鱼类组成的生态学分析[J]. 河北渔业, 1991(6): 8-11. HAN X F, WANG S A, CAO Y P, et al. Ecological analysis of fish composition after re-impounding in Baiyangdian Lake[J]. Hebei Fisheries, 1991(6): 8-11. |
| [10] |
曹玉萍, 王伟, 张永兵. 白洋淀鱼类组成现状[J]. 动物学杂志, 2003, 38(3): 65-69. CAO Y P, WANG W, ZHANG Y B. Present situation of fish stocks in Baiyangdian lake[J]. Chinese Journal of Zoology, 2003, 38(3): 65-69. DOI:10.13859/j.cjz.2003.03.029 |
| [11] |
赵春龙, 肖国华, 罗念涛, 等. 白洋淀鱼类组成现状分析[J]. 河北渔业, 2007(11): 49-50. ZHAO C L, XIAO G H, LUO N T, et al. Analysis on current fish composition in Baiyangdian Lake[J]. Hebei Fisheries, 2007(11): 49-50. |
| [12] |
谢松, 贺华东. "引黄济淀"后河北白洋淀鱼类资源组成现状分析[J]. 科技信息, 2010(9): 17. XIE S, HE H D. Present condition analysis of Hebei Baiyangdian lake fish stocks after diversion from Yellow River to the lake[J]. Science & Technology Information, 2010(9): 17. |
| [13] |
闫丽, 邓鹏春, 慕建东, 等. 白洋淀衡水湖鱼类多样性区系比较研究[J]. 水产研究, 2021, 8(2): 53-60. YAN L, DENG P C, MU J D, et al. Comparative study on fish diversity fauna in Hengshui Lake of Baiyangdian Lake[J]. Open Journal of Fisheries Research, 2021, 8(2): 53-60. |
| [14] |
王所安, 王志敏, 李国良, 等. 河北动物志: 鱼类[M]. 石家庄: 河北科学技术出版社, 2001. WANG S A, WANG Z M, LI G L, et al. The fauna of Hebei, China: Pisces[M]. Shijiazhuang: Hebei Science & Technology Press, 2001. |
| [15] |
朱松泉. 中国淡水鱼类检索[M]. 南京: 江苏科学技术出版社, 1995. ZHU S Q. The synopsis of freshwater fishes of China[M]. Nanjing: Jiangsu Science and Technology Publishing House, 1995. |
| [16] |
陈咏霞, 管敏. 河北省淡水虾虎鱼类形态特征和分类地位的研究[J]. 凯里学院学报, 2011, 29(6): 60-64. CHEN Y X, GUAN M. Study on morphological character and taxonomic status of the freshwater Gobioidei fishes of Hebei Province, China[J]. Journal of Kaili University, 2011, 29(6): 60-64. |
| [17] |
陈宜瑜. 中国动物志硬骨鱼纲鲤形目(中卷)[M]. 北京: 科学出版社, 1998. CHEN Y Y. Fauna sinica: Osteichthyes: Cypriniformes (volume Ⅱ)[M]. Beijing: Science Press, 1998. |
| [18] |
乐佩琦. 中国动物志: 硬骨鱼纲鲤形目(下卷)[M]. 北京: 科学出版社, 2000. YUE P Q. Fauna sinica: Osteichthyes: Cypriniformes (volume Ⅲ)[M]. Beijing: Science Press, 2000. |
| [19] |
MARGALEF R. Information theory in ecology[J]. General Systems, 1957, 3(1): 36-71. |
| [20] |
SHANNON C E. A mathematical theory of communication[J]. The Bell System Technical Journal, 1948, 27(3): 379-423. |
| [21] |
PIELOU E C. The measurement of diversity in different types of biological collections[J]. Journal of Theoretical Biology, 1966, 13: 131-144. |
| [22] |
PINKAS L, OLIPHANT M S, IVERSON I L K. Food habits of albacore, Bluefin tuna, and bonito in California waters[R]. Long Beach: Department of Fish and Game, 1971: 1-105.
|
| [23] |
YEMANE D, FIELD J G, LESLIE R W. Exploring the effects of fishing on fish assemblages using Abundance Biomass Comparison (ABC) curves[J]. ICES Journal of Marine Science, 2005, 62(3): 374-379. |
| [24] |
张东, 宛凤英, 储玲, 等. 青弋江鱼类分类群和功能群的α和β多样性纵向梯度格局[J]. 生物多样性, 2018, 26(1): 1-13. ZHANG D, WAN F Y, CHU L, et al. Longitudinal patterns in α and β diversity of the taxonomic and functional organizations of stream fish assemblages in the Qingyi River[J]. Biodiversity Science, 2018, 26(1): 1-13. |
| [25] |
CLARKE K R, WARWICK R M. Change in marine communities: an approach to statistical analysis and interpretation[M]. Plymouth: PRIMER-E Laboratory, 1994.
|
| [26] |
CLARKE K R. Non-parametric multivariate analyses of changes in community structure[J]. Austral Ecology, 1993, 18(1): 117-143. |
| [27] |
刘燕山, 谷先坤, 唐晟凯, 等. 东太湖鱼类群落结构特征及其与环境因子的关系[J]. 生态学报, 2021, 41(2): 769-780. LIU Y S, GU X K, TANG S K, et al. Fish community structure and its relationship with main environmental variables in the East Taihu Lake[J]. Acta Ecologica Sinica, 2021, 41(2): 769-780. |
| [28] |
李思忠. 中国淡水鱼类的分布区划[M]. 北京: 科学出版社, 1981. LI S Z. Distribution division of freshwater fishes in China[M]. Beijing: Science Press, 1981. |
| [29] |
常利伟. 白洋淀湖群的演变研究[D]. 长春: 东北师范大学, 2014. CHANG L W. The evolution of Baiyangdian Lake group research[D]. Changchun: Northeast Normal University, 2014. |
| [30] |
刘鹏飞, 张婉平, 徐东坡, 等. 钱塘江西湖段鱼类群落结构特征[J]. 上海海洋大学学报, 2021, 30(3): 525-535. LIU P F, ZHANG W P, XU D P, et al. Current situation of fish community structure in West Lake section of Qiantang River[J]. Journal of Shanghai Ocean University, 2021, 30(3): 525-535. |
| [31] |
张春光, 赵亚辉. 北京及其邻近地区的鱼类: 物种多样性、资源评价和原色图谱[M]. 北京: 科学出版社, 2013. ZHANG C G, ZHAO Y H. Fishes in Beijing and adjacent areas, China[M]. Beijing: Science Press, 2013. |
| [32] |
韩占江. 加大增殖放流力度推动白洋淀渔业可持续发展[J]. 河北渔业, 2008(10): 48, 50. HAN Z J. Improve sustainable development of fisheries in Baiyangdian Lake by strengthen artificial realeasing[J]. Hebei Fisheries, 2008(10): 48, 50. |
| [33] |
韩婵, 高春霞, 田思泉, 等. 淀山湖鱼类群落结构多样性的年际变化[J]. 上海海洋大学学报, 2014, 23(3): 403-410. HAN C, GAO C X, TIAN S Q, et al. Analysis of annual variations for fish community structure in Dianshan Lake[J]. Journal of Shanghai Ocean University, 2014, 23(3): 403-410. |
| [34] |
徐东坡, 凡迎春, 周彦锋, 等. 太湖鳑鲏亚科鱼类群落结构及其时空变动[J]. 上海海洋大学学报, 2018, 27(1): 115-125. XU D P, FAN Y C, ZHOU Y F, et al. Spatial and temporal variations of Acheilognathinae in Lake Taihu[J]. Journal of Shanghai Ocean University, 2018, 27(1): 115-125. |
| [35] |
田佳丽, 王银平, 李佩杰, 等. 长江近口段近岸段鱼类群落多样性现状[J]. 上海海洋大学学报, 2021, 30(2): 320-330. TIAN J L, WANG Y P, LI P J, et al. Species diversity of fish in the section near the Yangtze River estuary[J]. Journal of Shanghai Ocean University, 2021, 30(2): 320-330. |
| [36] |
徐开达, 张洪亮, 谢汉阳, 等. 中街山列岛水域甲壳类资源及其群落多样性[J]. 海洋渔业, 2012, 34(3): 308-315. XU K D, ZHANG H L, XIE H Y, et al. Resource density and community diversity of crustaceans in the waters of Zhongjieshan Islands[J]. Marine Fisheries, 2012, 34(3): 308-315. |
| [37] |
张静, 陈永俊, 张然, 等. 2008年夏季东山湾游泳动物种类的组成和多样性[J]. 应用海洋学学报, 2013, 32(2): 222-230. ZHANG J, CHEN Y J, ZHANG R, et al. Nekton species composition and biodiversity in Dongshan Bay during 2008 Summer[J]. Journal of Applied Oceanography, 2013, 32(2): 222-230. |
| [38] |
商书芹, 王帅帅, 殷旭旺. 济南市黄河流域鱼类群落结构及多样性研究[C]//2020(第八届)中国水生态大会论文集. 郑州: 北京沃特咨询有限公司, 2020: 1-16. SHANG S Q, WANG S S, YIN X W. Study on fish community structure and species diversity in Yellow River Basin of Jinan[C]//2020 (8th) China Water Ecology Conference. Zhengshou: Beijing Water Consulting Co., Ltd., 2020: 1-16. |
| [39] |
隋晓云. 中国淡水鱼类的分布格局研究[D]. 北京: 中国科学院水生生物研究所, 2010. SUI X Y. Studying on the distribution pattern of Chinese freshwater fishes[D] Beijing: Institute of Hydrobiology, Chinese Academy of Sciences, 2010. |
| [40] |
KEAST A. Trophic and spatial interrelationships in the fish species of an Ontario temperate lake[J]. Environmental Biology of Fishes, 1978, 3(1): 7-31. |
| [41] |
XIE S G, CUI Y B, LI Z J. Small fish communities in two regions of the Liangzi Lake, China, with or without submersed macrophytes[J]. Journal of Applied Ichthyology, 2010, 17(2): 89-92. |
| [42] |
YE S W, LI Z J, LEK-ANG S, et al. Community structure of small fishes in a shallow macrophytic lake(Niushan Lake) along the middle reach of the Yangtze River, China[J]. Aquatic Living Resources, 2006, 19(4): 349-359. |
| [43] |
程琳. 长江流域湖泊鱼类群落与主要环境因子关系及其预测模型[D]. 北京: 中国科学院水生生物研究所, 2011. CHENG L. Studying on the distribution pattern of Chinese freshwater fishes[D]. Beijing: Institute of Hydrobiology, Chinese Academy of Sciences, 2011. |
| [44] |
李经纬, 徐东坡, 李巍, 等. 滆湖鱼类群落时空分布及其与环境因子的关系[J]. 水产学报, 2022, 46(4): 546-556. LI J W, XU D P, LI W, et al. Spatial and temporal distribution of fish assemblages and its relationship with environmental factors in Lake Gehu[J]. Journal of Fisheries of China, 2022, 46(4): 546-556. |
| [45] |
胡翠林, 张洪亮, 张亚洲, 等. 带鱼保护区春秋季鱼类群落特征及与环境因子的关系[J]. 水产学报, 2018, 42(5): 694-703. HU C L, ZHANG H L, ZHANG Y Z, et al. Fish community structure and its relationship with environmental factors in the Nature Reserve of Trichiurus japonicus[J]. Journal of Fisheries of China, 2018, 42(5): 694-703. |
| [46] |
MAES J, STEVENS M, BREINE J. Modelling the migration opportunities of diadromous fish species along a gradient of dissolved oxygen concentration in a European tidal watershed[J]. Estuarine, Coastal and Shelf Science, 2007, 75(1/2): 151-162. |