 2022,
Vol. 31
2022,
Vol. 31

2. 国家远洋渔业工程技术研究中心,上海 201306;
3. 大洋渔业资源可持续开发教育部重点实验室,上海 201306;
4. 远洋渔业协同创新中心,上海 201306;
5. 上海海洋大学 图书馆,上海 201306
柔鱼(Ommastrephes bartramii)作为一种具有巨大开发潜力的重要经济头足类,广泛分布在太平洋海域[1]。柔鱼分为冬春生群(1—5月)和秋生群(9—2月)2个繁殖群体[2],其中冬春生的西部种群是我国鱿钓渔船自1993年以来的主要捕捞对象,年产量最高可达十几万吨[3-4]。柔鱼是短生命周期种类,其生活史过程与栖息地的海洋环境有重要关联[5-7],海洋环境因子的时空分布与变化显著影响着柔鱼资源的分布范围和资源丰度[8]。因此,准确了解海洋环境因子和柔鱼资源时空分布之间的关系,对于进一步解释柔鱼潜在渔场及适宜栖息地的位置,以及提高对其资源量的预测技术具有重要意义。崔雪森等[9]基于贝叶斯方法,结合海洋环境因子对西北太平洋柔鱼中心渔场位置进行了预报并得到较好的结果。余为等[10]利用序列分析中的交相关方法分析了Niño 3.4区海表面温度距平值(sea surface temperature anomaly, SSTA)与产卵场和育肥场的海表面温度(sea surface temperature, SST)、叶绿素a质量浓度(chlorophyll-a mass concentration,Chl.a)的滞后关系,并基于此结果建立柔鱼资源丰度预测模型。CHEN等[11]利用遥感环境数据对西北太平洋柔鱼栖息地进行了研究,用不同的环境变量组合方法构建栖息地指数模型(habitat suitability index,HSI),结果表明, 结合SST、海表面高度距平值(sea surface height anomaly,SSHA)和Chl.a等3个环境因子的算术平均法模型能很好地预测柔鱼的适宜生境。TIAN等[12]利用SST、35 m水层水温、317 m水层水温、海洋表面盐度、SSH等环境因子,证实了基于捕捞努力量的HSI模型能较好地定义柔鱼的最适栖息地。沈新强等[13]利用范例推理方法实现了中心渔场智能化预报,张月霞等[14]在传统范例推理的基础上,考虑了上级结果的相似性评价对次级检索结果的影响权重,并加入Chl.a及Chl.a梯度等特征因子,提高了中心渔场智能预报系统的准确度。袁红春等[15]和汪金涛等[16]基于传统的环境因子,利用BP神经网络模型对西北太平洋柔鱼资源量和中心渔场位置进行了准确的预报。
目前已有不少学者[4, 11, 17-22]开展了太平洋柔鱼资源丰度与海洋环境因子之间关系的模型构建研究,这些模型在构建时主要考虑了SST、Chl.a和SSH等传统的海洋环境因子。SST是影响柔鱼生命史及空间分布的关键因子[3, 23],对太平洋柔鱼渔场的时空分布影响显著[24-26]。西北太平洋产卵场和育肥场的Chl.a与柔鱼资源丰度关系密切,可作为鱿鱼中心渔场分析中的一个重要参考指标[10]。SSH作为冷暖水的指标,当SSH高于平均海面高度时,海流的辐合或涌升使得该海域营养盐丰富[27-28],有利于鱼类的生长、发育及繁殖,进而丰富了渔场资源[29],因此SSH亦与西北太平洋柔鱼的分布密切相关。此外,海水质量变化、地转流等海洋物理环境因子对海洋环境变化也具有重要影响,但在构建柔鱼资源的时空分布与海洋要素的关系时很少被考虑。本文融合多种卫星传感器的海洋观测数据,综合考虑SST、Chl.a、SSHA、海水质量变化和地转流与西北太平洋柔鱼资源丰度之间的关联,并基于卫星遥感观测与实际的捕捞数据进行预测模型的构建和评估,提高西北太平洋柔鱼资源分布的预测精度,为增加柔鱼捕捞产量、提高捕捞效率提供指导。
1 材料与方法 1.1 渔业生产数据本文的柔鱼生产统计数据来自上海海洋大学中国远洋渔业数据中心,选取了包含2004—2017年5—12月的西北太平洋海域(150°E~175°E和35°N~50°N)的柔鱼历史渔业捕捞数据。这些数据主要统计了捕捞日期(年、月、日)、捕捞位置(经度、纬度)、日产量(t)和捕捞努力量(d),数据的空间分辨率为1°×1°。中国鱿钓船在这一海域的产量占据了北太平洋柔鱼总产量的80%以上,其中95%以上为柔鱼冬春生西部群体,且无副渔获物。
单位捕捞努力量渔获量(catch per unit effort, CPUE)常被假设与资源量成正比[30],因此亦被用来表征资源丰度。使用名义CPUE作为定义柔鱼丰度的指标因子,通过对逐日捕捞数据进行月平均后,采用如下公式计算2004—2017年5—12月的逐月平均、空间分辨率为1°×1°的柔鱼CPUE:
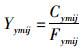 (1)
(1)
式中:Yymij为月平均名义CPUE,t/d;Cymij为一个渔区内所有渔船总产量,t;Fymij为总作业次数,即统计一个渔区内所有渔船总作业天数,d;i为经度;j为纬度;m为月份;y为年份。在本文后续的分析中,将使用实际捕捞数据计算得到的名义CPUE作为真值,用于模型模拟和预测精度的评估。
1.2 卫星遥感数据研究所使用的SST和Chl.a数据均来自于中分辨率成像光谱仪MODIS传感器(http://oceandata.sci.gsfc.nasa.gov/)。MODIS是由美国宇航局研制的大型空间传感器,搭载于Terra和Aqua上。MODIS数据有36个波段,每日或每两日可获得一次全球观测数据。为了保证研究区域内数据的完整性,我们选取了Terra和Aqua卫星平台上的月平均SST和Chl.a数据产品,空间分辨率为9 km。在实际操作中,我们将Terra和Aqua得到的数据产品进行平均后,然后将平均后的SST和Chl.a进行重采样处理,获得2004—2017年5—12月的空间分辨率为1°×1°的月平均SST和Chl.a。
为了获取SSHA数据,选用法国航天局AVISO中心提供的海面高异常格网数据(https://www.aviso.altimetry.fr/data.html),该数据融合了Topex/Poseidon、ERS-1/2、Jason-1、Envisat和Jason-2多颗卫星的测高资料,选取20年(1993—2013)的平均海面高模型作为参考基准。该数据与单颗测高卫星的数据相比,不仅极大地提高了海平面变化的空间分辨率,而且能够提供更长时间序列的海面高异常数据。对于不同的测高卫星的数据融合,采用相同观测时间段的观测资料,计算出不同的测高卫星的系统性偏差。该数据经过了多种必要的地球物理校正,包括电离层延迟校正、对流层干湿分量校正、固体潮和海潮校正、海洋负荷潮汐校正、极潮校正、电磁偏差校正、仪器校正和反变气压计校正等。
为了获取海水质量变化,选取美国德克萨斯大学空间研究中心(CSR)发布的GRACE卫星的Level-2 RL05版本的时变重力场月平均数据(http://www2.csr.utexas.edu/grace/RL05.html)进行后续处理,该版本数据提供平均每个月的地球重力场模型的球谐系数。由于GRACE重力场模型系数只展开至有限阶数,表面密度变化也只能计算到截断阶次,因此在计算地球质量变化时将不可避免会产生截断误差。为了最大程度地降低GRACE噪音基础上尽量保留原有数据的信号,以保证其空间分辨率,本文中选取了500 km为平滑半径,然后利用高斯核函数对GRACE重力位系数进行了平滑滤波处理。
在GRACE数据处理中还需对低阶重力场系数进行处理。利用海洋模型和GRACE数据解算得到的一阶重力场系数来代替GRACE重力场系数中的一阶项,并利用卫星激光测距(SLR)得到的二阶项来代替GRACE重力位系数中的二阶项。由于GRACE在数据处理中扣除了大气和海洋高频非潮汐变化影响,在利用GRACE Level-2数据研究海水质量变化时,还应恢复该影响。具体的数据处理步骤详见参考文献[31-32],经过上述处理之后,基于GRACE数据得到了空间分辨率为1°×1°,时间跨度为2004年1月至2017年12月的月平均全球海水质量变化,然后从中截取出研究海域内的海水质量变化结果。
此外,在研究地转流时,需要首先确定海面地形。海面地形(ζ)是海面高度(h)与大地水准面高度(N)差,可表示为
 (2)
(2)
式中:海面高度h可以由卫星测高数据获得,卫星测高数据主要采用法国国家空间中心AVISO数据中心提供的融合多颗卫星高度计(ERS-1/2、Topex/Posedion、ENVISAT和Jason-1/Jason-2)的网格化海面高度数据,其覆盖区域从66°S~66°N,空间分辨率为1°×1°[33];大地水准面高度N则由高精度的卫星重力场模型EGM2008获得,EGM2008重力场模型是由美国国家地理空间情报局利用地面重力数据、GRACE卫星数据等重力场信息完成的超高阶地球重力场模型,模型阶次完全至2159(球谐系数的阶扩展至2190,次为2159)[34]。然后将海面地形ζ代入如下公式即可获得地转流:
 (3)
(3)
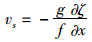 (4)
(4)
式中:f为科里奥利参数;g为重力加速度;us,vs分别表示地转流的东西分量和南北分量。将地转流的东西分量和南北分量进行合成后,即可进一步获取地转流的大小。经过上述处理之后,可以得到空间分辨率为1°×1°,时间跨度为2004年1月至2017年12月的月平均地转流东西分量和南北分量,然后从中获得需要的西北太平洋区域的地转流大小。
1.3 BP神经网络模型BP神经网络属于前向网络,可包含多个隐含层,具备处理线性不可分问题的能力。BP神经网络具备以下特点[35]:网络由多层构成,层与层之间全连接,同一层之间的神经元无连接。多层的网络设计,使BP网络能够从输入中挖掘更多的信息,完成更复杂的任务。采用反向传播算法进行学习。在BP网络中,数据从输入层经隐含层逐层向后传播,训练网络权值时,则沿着减少误差的方向,从输出层经过中间各层向前修正网络的连接权值。随着学习的不断进行,最终的误差越来越小。
BP神经网络的输入层为上述海洋环境因子,经神经网络隐含层传递函数处理后,由输出层输出CPUE;由于输出层输出的CPUE与实际的CPUE存在偏差,神经网络通过评估偏差大小,如果偏差过大,则将偏差反向传输至输入层,并通过迭代更新并调整输入层与隐含层之间的连接权值,直到输出层输出的CPUE与实际CPUE之间的偏差小于指定的阈值或达到设置的迭代次数,即可认为BP神经网络的训练学习达到了所需精度要求,训练得到的神经网络模型即可以用于CPUE的预测。
隐含层节点数对BP神经网络的性能有很大影响,较多的隐含层节点数可以带来更好的性能,但可能导致训练时间过长。采用经验公式设置隐含层节点M:
 (5)
(5)
式中:n为输入层神经元个数;[x]表示不小于x的最小整数。
在进行BP神经网络的实例构建中,选用的训练函数为Levenberg-Marquardt(LM)反向传播算法。该算法类似牛顿法,是为了在修正速率时避免计算Hessian矩阵而设计的[36]。当误差性能函数e具有平方和误差的形式时,Hessian矩阵可近似表示为
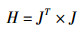 (6)
(6)
梯度可以表示为
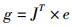 (7)
(7)
式中:J为包含误差性能函数对网络权值一阶导数的雅可比矩阵。LM算法根据下式修正网络权值:
 (8)
(8)
当μ=0时,LM算法退化为牛顿法;当μ很大时,上式相当于步长较小的梯度下降法。柔鱼雅可比矩阵的计算比Hessian矩阵易于计算,因此速度非常快。
BP神经网络的构建是在Matlab 2018b的环境下进行并完成的。所设置的BP神经网络的其他主要参数包括:学习速率0.01,训练精度0.001,最大训练次数为1 000次,隐含层和输出层的传递函数均为线性函数purelin。
2 结果为了客观评价基于BP神经网络进行西北太平洋柔鱼资源丰度模拟和预测的精度,将2004—2017年5—12月时空同步的SST、Chl.a、SSHA、海水质量变化、地转流和CPUE月平均结果在时间域上进行分段处理,选取的T1时间段和T2时间段分别为2004—2012年5—12月和2013—2017年5—12月。T1时间段的海洋环境和CPUE数据主要用于建立西北太平洋的海洋环境因子与柔鱼CPUE之间的BP神经网络模型,并进行内符合精度评价,而T2时间段的海洋环境数据则主要用于作为建立好的BP神经网络模型的输入参数,然后将输出结果与实际的CPUE数据进行校验,通过进行外符合精度评定来分析所建立的BP神经网络模型进行柔鱼CPUE预测的精度。
此外,为了证实引入海水质量变化和地转流等海洋环境因子的必要性,在进行BP神经网络建模时,设计了2种不同海洋环境因子的组合方案作为模型输入层,并进行比较分析。方案Ⅰ主要使用SST、Chl.a和SSHA等传统的海洋环境因子作为BP神经网络模型的输入参数,而方案Ⅱ则在上述传统海洋环境因子的基础上加入海水质量变化和地转流作为BP神经网络模型的输入参数。
2.1 BP神经网络模型对西北太平洋柔鱼资源丰度的精度评估图 1给出了在T1时间段,基于方案Ⅰ和方案Ⅱ的海洋环境因子作为输入参数,并结合鱿钓船实测CPUE进行BP神经网络建模后,BP神经网络模型在选取的某一格点(43°N、155°E)处模拟和预测的CPUE与名义CPUE的比较结果。为了验证BP神经网络模型输出CPUE的精度,图 1还给出了该格点的CPUE模拟结果与实测值的标准差(STD)和均方根误差(RMSE),分别来表征BP神经网络模拟和预测的CPUE与实测名义CPUE的离散与精确程度。

|
图 1 北纬43°、东经155°处鱿钓船实测西北太平洋柔鱼CPUE与BP神经网络模型模拟和预测的CPUE结果之间的比较 Fig. 1 Comparison of the CPUE of O.bartramii at the grid (43° N, 155° E) in the northwest Pacific from squid jigging vessel with BP neural network modelled and predicted CPUE |
从图 1可以看出,在T1时间段利用方案Ⅰ和方案Ⅱ的海洋环境因子作为BP神经网络模型的输入,并分别建立模型后,模拟结果与实测的名义CPUE均具有较好的一致性。此外,2种方案模拟2004—2012年5—12月的西北太平洋柔鱼CPUE的STD和RMSE均小于1.80 t/d,且方案Ⅱ对西北太平洋柔鱼CPUE的模拟较方案Ⅰ的可靠性和精度都略高。这说明在利用BP神经网络模型建立西北太平洋海洋环境因子与柔鱼CPUE之间的关系时,在输入传统海洋环境变量的基础上,额外加入海水质量变化和地转流,不仅是可行的,还可以进一步提高所建模型对柔鱼CPUE的模拟能力。
为了验证BP神经网络模型对柔鱼CPUE模拟的能力,图 2以2012年7月为例,分析在不同的模型输入方案下的柔鱼CPUE的空间分布情况。结果显示,方案Ⅱ模拟的2012年7月的西北太平洋柔鱼CPUE的空间分布与实测的CPUE结果更为接近。以名义CPUE为参考,方案Ⅰ模拟的2012年7月的西北太平洋柔鱼CPUE的平均偏差和均方根误差分别为0.62和0.88 t/d,而方案Ⅱ模拟结果的平均偏差和均方根误差分别为0.26和0.55 t/d。因此,方案Ⅱ的模拟结果在2012年7月要优于方案Ⅰ。

|
图 2 2012年7月鱿钓船实测西北太平洋柔鱼CPUE的空间分布以及不同输入方案下的BP神经网络模型在该月份模拟的CPUE空间分布 Fig. 2 Spatial distributions of the CPUE of O. bartramii in July 2012 in the northwest Pacific from squid jigging vessel as well as BP neural network model from different schemes |
以鱿钓船实测CPUE为参考,利用方案Ⅰ和方案Ⅱ对T1时间段内逐月的西北太平洋柔鱼CPUE模拟结果的标准差和均方根误差如图 3所示。整体上,与方案Ⅰ相比,方案Ⅱ在较低纬度地区的160°E~175°E范围内的模拟CPUE与实测结果更为接近。方案Ⅰ和方案Ⅱ模拟结果的平均STD分别为0.95和0.74 t/d,说明方案Ⅱ模拟的西北太平洋柔鱼CPUE与实测值的离散程度要低于方案Ⅰ。此外,方案Ⅰ和方案Ⅱ的平均RMSE分别为0.94和0.73 t/d,也进一步说明了方案Ⅱ模拟西北太平洋柔鱼CPUE的精度整体上要高于方案Ⅰ。

|
图 3 不同参数输入方案构建的BP神经网络模型模拟的2004—2012年5—12月的西北太平洋柔鱼CPUE的误差分布 Fig. 3 Spatial distribution of RMSE of BP neural network modelled CPUE of O. bartramii from May to December during 2004 to 2012 |
将基于T1时间段数据建立的BP神经网络模型,应用于T2时间段的海洋环境因子时,即可实现对2013—2017年5—12月的柔鱼CPUE的预测。图 1基于BP神经网络模型预测的43°N、155°E处的柔鱼CPUE的结果显示,引入海水质量变化和地转流参与BP神经网络模型构建后,其预测的柔鱼CPUE的STD和RMSE均比传统构建方法低。方案Ⅰ和方案Ⅱ预测的43°N、155°E处的柔鱼CPUE的STD分别为1.39和0.62 t/d,RMSE分别为1.36和0.67 t/d。因此,方案Ⅱ比方案Ⅰ在43°N、155°E处预测的柔鱼CPUE的精度得到了显著提高。
图 4选取了2013年11月作为分析,给出了鱿钓船实测CPUE的空间分布,以及基于方案Ⅰ和方案Ⅱ采用BP神经网络预测的柔鱼CPUE的空间分布情况。整体上,方案Ⅰ预测的2013年11月的西北太平洋柔鱼CPUE的平均偏差和均方根误差分别为-0.44和1.30 t/d,而方案Ⅱ预测结果的平均偏差和均方根误差分别为0.05和1.21 t/d。方案Ⅱ在2013年11月的预测结果比方案Ⅰ的均方根误差分别提高了约7%。

|
图 4 2013年11月鱿钓船实测西北太平洋柔鱼CPUE的空间分布以及不同输入方案下的BP神经网络模型在该月份预测的CPUE空间分布 Fig. 4 Spatial distributions of the CPUE of O. bartramiiin November 2013 in the northwest Pacific from squid jigging vessel as well as BP neural network model from different schemes |
从图 5可以明显地看到,基于方案Ⅰ预测的柔鱼CPUE存在非常显著的异常结果,而方案Ⅱ的预测结果的精度在整个研究区域内较为一致。方案Ⅰ和方案Ⅱ预测的CPUE结果的平均STD分别为1.98和1.36 t/d,平均RMSE分别为2.11和1.55 t/d。因此,与方案Ⅰ相比,在方案Ⅱ中额外引入海水质量变化和地转流后,能有效减弱方案Ⅰ预测CPUE的误差异常偏大值,方案Ⅱ比方案Ⅰ预测的西北太平洋柔鱼CPUE的STD减少了31%,RMSE减少了26%。

|
图 5 不同参数输入方案构建的BP神经网络模型预测的2013—2017年5—12月的西北太平洋柔鱼CPUE的误差分布 Fig. 5 Spatial distribution of RMSE of BP neural network predicted CPUE of O. bartramii from May to December during 2013 to 2017 |
西北太平洋柔鱼渔场时空分布与所处的海洋环境密切相关。在西北太平洋,黑潮暖流和亲潮寒流形成了广泛的交汇区,这为柔鱼的生长和繁育提供了良好的生物和非生物环境条件。在过去的研究[8-10, 29]中,西北太平洋的SST、Chl.a、SSHA等海洋环境因子均被认为可用于寻找和开发柔鱼渔场。因此,更好地建立海洋环境因子与柔鱼资源的时空分布之间的联系,对进一步了解预测柔鱼的资源变动,认识柔鱼栖息地与海洋环境及气候变化的关系,以及促进柔鱼资源的可持续利用,均具有重要的意义。但是,以往在建立柔鱼资源丰度的分布与海洋环境因子之间的关系模型时,往往仅考虑了SST、Chl.a和SSHA等传统的海洋环境信息,并未考虑海水质量变化、地转流等其他海洋条件对柔鱼资源丰度的影响。然而,海水质量变化和地转流与海水温度、海洋中的营养盐类等海洋环境密切相关,从而对柔鱼的栖息环境条件有重要影响,使得忽略这些因素可能在一定程度上影响模型的预测精度。
本文在传统海洋环境因子的基础上,基于卫星遥感观测额外引入了海水质量变化和地转流等海洋环境信息,利用BP神经网络模型建立了西北太平洋柔鱼资源丰度与多海洋环境因子之间的关系,并对所建模型进行西北太平洋柔鱼CPUE模拟和预测的精度进行了评估。结果发现,基于2004—2012年5—12月的海洋环境因子与实测CPUE进行BP神经网络模型构建时,在SST、Chl.a和SSHA等传统海洋环境因子的基础上,引入海水质量变化和地转流,能有效提高BP神经网络模型对西北太平洋柔鱼CPUE的模拟能力,这可能与海水质量变化和地转流的时间分布更能反映柔鱼栖息地环境特征有关。与传统方案相比,改进方案模拟西北太平洋柔鱼CPUE的STD减少了22%,且RMSE亦减少了22%。
此外,基于已建BP神经网络模型对2013—2017年5—12月进行西北太平洋柔鱼CPUE的预测时,发现传统方案易导致局部海域的预测结果存在明显的异常偏差,而改进方案的CPUE预测结果的精度在整个研究区域内较为一致。整体上来看,额外引入海水质量变化和地转流建立的BP神经网络模型与仅使用传统海洋环境因子相比,所预测的西北太平洋柔鱼CPUE的STD减少了31%,RMSE减少了26%。与现有的预测模型仅考虑传统海洋环境因子相比,本文提出在传统海洋环境因子的基础上,额外引入海水质量变化和地转流,使得综合使用的海洋环境因子能更好地表征海洋环境特征和柔鱼的栖息环境。因此,本文提出的综合多源卫星遥感观测的西北太平洋柔鱼CPUE预测方法可更好地模拟柔鱼的时空分布,并有效提高西北太平洋柔鱼资源的预测能力。
| [1] |
ROPER C F E, SWEENEY M J, NAUEN C E. Cephalopods of the world: an annotated and illustrated catalogue of species of interest to fisheries[M]. Rome: FAO Species Catalogue for Fishery Purposes, 1984: 1-277.
|
| [2] |
YATSU A, MIDORIKAWA S, SHIMADA T, et al. Age and growth of the neon flying squid, Ommastrephesbartrami, in the north Pacific ocean[J]. Fisheries Research, 1997, 29(3): 257-270. DOI:10.1016/S0165-7836(96)00541-3 |
| [3] |
CHEN X J, CHEN Y, TIAN S Q, et al. An assessment of the west winter-spring cohort of neon flying squid (Ommastrephes bartramii) in the northwest Pacific Ocean[J]. Fisheries Research, 2008, 92(2/3): 221-230. |
| [4] |
CHEN X J, ZHAO X H, CHEN Y. Influence of El Niño/La Niña on the western winter-spring cohort of neon flying squid (Ommastrephes bartramii) in the northwestern Pacific Ocean[J]. ICES Journal of Marine Science, 2007, 64(6): 1152-1160. DOI:10.1093/icesjms/fsm103 |
| [5] |
ICHII T, MAHAPATRA K, SAKAI M, et al. Changes in abundance of the neon flying squid Ommastrephes bartramii in relation to climate change in the central north Pacific Ocean[J]. Marine Ecology Progress Series, 2011, 441: 151-164. DOI:10.3354/meps09365 |
| [6] |
ALABIA I D, SAITOH S I, HIRAWAKE T, et al. Elucidating the potential squid habitat responses in the central North Pacific to the recent ENSO flavors[J]. Hydrobiologia, 2016, 772(1): 215-227. DOI:10.1007/s10750-016-2662-5 |
| [7] |
IGARASHI H, ICHII T, SAKAI M, et al. Possible link between interannual variation of neon flying squid (Ommastrephes bartramii) abundance in the north Pacific and the climate phase shift in 1998/1999[J]. Progress in Oceanography, 2017, 150: 20-34. DOI:10.1016/j.pocean.2015.03.008 |
| [8] |
YU W, CHEN X J, YI Q, et al. Variability of suitable habitat of western winter-spring cohort for neon flying squid in the northwest Pacific under anomalous environments[J]. PLoS One, 2015, 10(4): e0122997. DOI:10.1371/journal.pone.0122997 |
| [9] |
崔雪森, 唐峰华, 张衡, 等. 基于朴素贝叶斯的西北太平洋柔鱼渔场预报模型的建立[J]. 中国海洋大学学报, 2015, 45(2): 37-43. CUI X S, TANG F H, ZHANG H, et al. The establishment of northwest Pacific Ommastrephes bartramii fishing ground forecasting model based on naive bayesmethod[J]. Periodical of Ocean University of China, 2015, 45(2): 37-43. |
| [10] |
余为, 陈新军. 西北太平洋柔鱼栖息地环境因子分析及其对资源丰度的影响[J]. 生态学报, 2015, 35(15): 5032-5039. YU W, CHEN X J. Analysis of environmental conditions and their influence on the abundance of neon flying squid in the northwest Pacific Ocean[J]. Acta Ecologica Sinica, 2015, 35(15): 5032-5039. |
| [11] |
CHEN X J, TIAN S Q, CHEN Y, et al. A modeling approach to identify optimal habitat and suitable fishing grounds for neon flying squid (Ommastrephes bartramii) in the northwest Pacific Ocean[J]. Fishery Bulletin, 2010, 108(1): 1-14. |
| [12] |
TIAN S Q, CHEN X J, CHEN Y, et al. Evaluating habitat suitability indices derived from CPUE and fishing effort data for Ommatrephes bratramii in the northwestern Pacific Ocean[J]. Fisheries Research, 2009, 95(2/3): 181-188. |
| [13] |
沈新强, 樊伟, 韩士鑫, 等. 中心渔场智能预报系统的设计与实现[J]. 中国水产科学, 2000, 7(2): 69-72. SHEN X Q, FAN W, HAN S X, et al. Design and realization of intelligent predicting system of central fishing ground[J]. Journal of Fishery Sciences of China, 2000, 7(2): 69-72. DOI:10.3321/j.issn:1005-8737.2000.02.015 |
| [14] |
张月霞, 丘仲锋, 伍玉梅, 等. 基于案例推理的东海区鲐鱼中心渔场预报[J]. 海洋科学, 2009, 33(6): 8-11. ZHANG Y X, QIU Z F, WU Y M, et al. Predicting central fishing ground of Scomber japonica in East China Sea based on case-based reasoning[J]. Marine Sciences, 2009, 33(6): 8-11. |
| [15] |
袁红春, 顾怡婷, 汪金涛, 等. 西北太平洋柔鱼中长期预测方法研究[J]. 海洋科学, 2013, 37(10): 65-70. YUAN H C, GU Y T, WANG J T, et al. Study on medium to long term forecasting of squid in the northwestern Pacific[J]. Marine Sciences, 2013, 37(10): 65-70. |
| [16] |
汪金涛, 陈新军, 雷林, 等. 基于频度统计和神经网络的北太平洋柔鱼渔场预报模型比较[J]. 广东海洋大学学报, 2014, 34(3): 82-87. WANG J T, CHEN X J, LEI L, et al. Comparisons between two forecasting models of fishing ground based on frequency statistics and neural network for Ommastrephes bartramiiin the North Pacific Ocean[J]. Journal of Guangdong Ocean University, 2014, 34(3): 82-87. DOI:10.3969/j.issn.1673-9159.2014.03.015 |
| [17] |
YU W, CHEN X J, YI Q. Fishing ground distribution of Neon Flying Squid(Ommastrephes bartramii) in relation to oceanographic conditions in the Northwest Pacific Ocean[J]. Journal of Ocean University of China, 2017, 16(6): 1157-1166. DOI:10.1007/s11802-017-3354-0 |
| [18] |
NISHIKAWA H, IGARASHI H, ISHIKAWA Y, et al. Impact of paralarvae and juveniles feeding environment on the neon flying squid (Ommastrephes bartramii) winter-spring cohort stock[J]. Fisheries Oceanography, 2014, 23(4): 289-303. DOI:10.1111/fog.12064 |
| [19] |
YATSU A, WATANABE T, MORI J, et al. Interannual variability in stock abundance of the neon flying squid, Ommastrephes bartramii, in the north Pacific Ocean during 1979-1998: impact of driftnet fishing and oceanographic conditions[J]. Fisheries Oceanography, 2000, 9(2): 163-170. DOI:10.1046/j.1365-2419.2000.00130.x |
| [20] |
ALABIA I D, SAITOH S I, MUGO R, et al. Seasonal potential fishing ground prediction of neon flying squid (Ommastrephes bartramii) in the western and central north Pacific[J]. Fisheries Oceanography, 2015, 24(2): 190-203. DOI:10.1111/fog.12102 |
| [21] |
YU W, CHEN X J, YI Q, et al. Spatio-temporal distributions and habitat hotspots of the winter-spring cohort of neon flying squid Ommastrephes bartramii in relation to oceanographic conditions in the northwest Pacific Ocean[J]. Fisheries Research, 2016, 175: 103-115. DOI:10.1016/j.fishres.2015.11.026 |
| [22] |
YU W, WEN J, ZHANG Z, et al. Spatio-temporal variations in the potential habitat of a pelagic commercial squid[J]. Journal of Marine Systems, 2020, 206: 103339. DOI:10.1016/j.jmarsys.2020.103339 |
| [23] |
陈新军, 田思泉, 许柳雄. 西北太平洋海域柔鱼产卵场和作业渔场的水温年间比较及其与资源丰度的关系[J]. 上海水产大学学报, 2005, 14(2): 168-175. CHEN X J, TIAN S Q, XU L X. Analysis on changes of surface water temperature in the spawning and feeding ground of Ommastrephes bartrami and its relationship with abundance index in the northwestern Pacific Ocean[J]. Journal of Shanghai Fisheries University, 2005, 14(2): 168-175. |
| [24] |
SAKURAI Y, KIYOFUJI H, SAITOH S, et al. Changes in inferred spawning areas of Todarodes pacificus (Cephalopoda: Ommastrephidae) due to changing environmental conditions[J]. ICES Journal of Marine Science, 2000, 57(1): 24-30. DOI:10.1006/jmsc.2000.0667 |
| [25] |
CAO J, CHEN X J, CHEN Y. Influence of surface oceanographic variability on abundance of the western winter-spring cohort of neon flying squid Ommastrephes bartramii in the NW Pacific Ocean[J]. Marine Ecology Progress Series, 2009, 381: 119-127. DOI:10.3354/meps07969 |
| [26] |
BOWER J R, ICHII T. The red flying squid (Ommastrephes bartramii): a review of recent research and the fishery in Japan[J]. Fisheries Research, 2005, 76(1): 39-55. DOI:10.1016/j.fishres.2005.05.009 |
| [27] |
POLOVINA J J, KLEIBER P, KOBAYASHI D R. Application of TOPEX-POSEIDON satellite altimetry to simulate transport dynamics of larvae of spiny lobster, Panulirus marginatus, in the northwestern Hawaiian Islands, 1993-1996[J]. Fishery Bulletin, 1999, 97(1): 132-143. |
| [28] |
CHEN X J, LI G, FENG B, et al. Habitat suitability index of Chub mackerel (Scomber japonicus) from July to September in the East China Sea[J]. Journal of Oceanography, 2009, 65(1): 93-102. DOI:10.1007/s10872-009-0009-9 |
| [29] |
张炜, 张健. 西南大西洋阿根廷滑柔鱼渔场与主要海洋环境因子关系探讨[J]. 上海水产大学学报, 2008, 17(4): 471-475. ZHANG W, ZHANG J. A discussion about relationship between the distribution of production of Illex argentinus and marine environmental factors in the southwest Atlantic Ocean[J]. Journal of Shanghai Fisheries University, 2008, 17(4): 471-475. |
| [30] |
MACCALL A D. Quantitative fish dynamics[J]. Journal of the American Statistical Association, 2001, 96(454): 781. |
| [31] |
WAHR J, MOLENAAR M, BRYAN F. Timevariability of the Earth's gravity field: Hydrological and oceanic effects and their possible detection using GRACE[J]. Journal of Geophysical Research: Solid Earth, 1998, 103(B12): 30205-30229. DOI:10.1029/98JB02844 |
| [32] |
CHAMBERS D P, BONIN J A. Evaluation of Release-05 GRACE time-variable gravity coefficients over the ocean[J]. Ocean Science, 2012, 8(5): 859-868. DOI:10.5194/os-8-859-2012 |
| [33] |
DUCET N, LE TRAON P Y, REVERDIN G. Global high-resolution mapping of ocean circulation from TOPEX/Poseidon and ERS-1 and -2[J]. Journal of Geophysical Research: Oceans, 2000, 105(C8): 19477-19498. DOI:10.1029/2000JC900063 |
| [34] |
PAVLIS N K, HOLMES S A, KENYON S C, et al. An earth gravitational model todegree 2160: EGM2008[C]//Proceedings of EGU General Assembly 2008. Vienna: National Geospatial-Intelligence Agency, 2008.
|
| [35] |
ABE J M. Paraconsistent artificial neural networks: an introduction[M]//NEGOITA M G, HOWLETT R J, JAIN L C. Knowledge-Based Intelligent Information and Engineering Systems. Berlin: Springer, 2004: 942-948.
|
| [36] |
SANGWAN P, DESHWAL D, KUMAR D, et al. Isolated word language identification system with hybrid features from a deep belief network[J]. International Journal of Communication Systems, 2020, e4418. DOI:10.1002/dac.4418 |
 ,
CHEN Fanglin1,
CHEN Xinjun1,2,3,4,
YU Wei1,2,3,4,
FENG Guiping1,2,3,4,
LI Yangdong1,2,3,4,
ZENG Wei5
,
CHEN Fanglin1,
CHEN Xinjun1,2,3,4,
YU Wei1,2,3,4,
FENG Guiping1,2,3,4,
LI Yangdong1,2,3,4,
ZENG Wei5
2. National Engineering Research Center for Oceanic Fisheries, Shanghai 201306, China;
3. Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai 201306, China;
4. Collaborative Innovation Center for Distant-water Fisheries, Shanghai 201306, China;
5. Library, Shanghai Ocean University, Shanghai 201306, China