 2022,
Vol. 31
2022,
Vol. 31



2. 上海海洋大学 海洋牧场工程技术研究中心, 上海 201306;
3. 国家海洋局东海预报中心, 上海 200136
枸杞岛位于长江入海口浊度锋面东侧,受长江冲淡水扩展的影响,周边水体具有高营养盐、低浊度的特征,十分利于浮游动植物和大型附着藻类的生长及繁殖[1]。正是优异的自然条件使得贻贝养殖业成为枸杞岛的支柱性产业之一[2]。枸杞岛贻贝养殖方式以浮筏吊养为主,即在浅海水面利用浮子和绳索组成浮筏以缆绳固定于海底,并用吊绳将附着贻贝的养成绳和海面浮筏连接在一起[3]。一般地,养成绳长约2.3~2.8 m,吊绳长约1.5~1.7 m,浮子直径约0.35 m,高度约0.47 m,整个养殖设施及养殖对象约据海面4.0 m的深度。连片的养殖筏架形成了复杂的三维生境,为海洋生物提供庇护和栖息场所[4-5]。泡沫、绳索等疏松多孔的材质构成的养殖浮筏是大型附生海藻的良好附着基,贻贝排泄物释放的营养盐为大型附生海藻的生长提供了直接的营养来源[6-7]。相比天然海藻场,贻贝养殖场内附生海藻具有分布广、生长快、生物量大的特点[8-9]。附生海藻有助于改善养殖海域的水质环境,其脱落的有机碎屑可作为贻贝的部分饵料来源,也有助于增加海底生物群落的多样性[10-12]。由于浮筏及贝串在海表产生的阻流效应改变了海域的水动力条件,使得从养殖筏架上脱落的海藻碎屑的输运和沉降有异于天然海域。为了探究养殖海域附生海藻腐烂脱落后的碎屑在海表阻流效应下的运动情况,本文建立考虑养殖设施阻力的三维海洋数值模型模拟养殖海域的水动力条件,并采用拉格朗日质点追踪的方法探究脱落的海藻碎屑的输运范围及沉降情况。
1 材料与方法 1.1 研究区域模型计算域包含长江口,杭州湾及邻近海域(图 1),重点加密枸杞岛附近海域。模型水平方向采用Arakawa C网格,网格数为336×256,最高水平分辨率为70 m;垂向采用sigma坐标,分为31层,重点加密0~3.8 m贝串所在的深度,垂向最大分辨率为水深的0.01倍。模型采用潮位边界,由16个分潮驱动(M2、S2、N2、K2、K1、O1、P1、Q1、MU2、NU2、T2、L2、2N2、J1、M1、OO1),振幅和迟角数据由TPXO7.1数据集内插得到,该数据集在沿海区域分辨率最大可达1/30°[13]。

|
图 1 模型计算区域 Fig. 1 Model domain |
模型验证采用养殖区内部及外围、嵊山北部海域的观测资料(图 2)。潮位验证采用2020年7月19日10点—7月29日8点(S1)、2018年11月5日9点—11日12点(S2,S3)的实测数据和绿华山验潮站2020年7月18日—2020年7月31日的潮位数据。S3站点位于养殖场外,流场观测时间为2018年11月5日9点—11月6日12点(大潮期间)。在枸杞岛西北岸养殖区内部布设观测站点(S4~S6)用来验证模型流速模拟结果的准确性,S4站点观测处于大潮期间(2020年7月21日9点—7月25日10点),S5、S6站点观测处于小潮期间(2020年7月25日15点—7月28日15点)。

|
粉红色星号表示两块最大养殖区的中心位置;黑色圆圈表示以红色点为中心的5 km区域。 Pink asterisk is the center of the two largest mussel farm; The black circle represents the 5 km area centered on the red dot. 图 2 水深与站点分布 Fig. 2 Depth and observation sites |
养殖贝串约占据海表 0~4.0 m的深度,占到养殖区水深的1/7~1/3,对养殖海域表层一定深度海水的潮流运动产生阻碍作用。传统的对数法近似认为贝串对表层水流的阻力类似于潮流底边界的摩擦阻力且符合对数曲线的形式[14-15],因此对数法不适合本研究。本文采用的动量汇法考虑了养殖设施阻力作用使得表层水体损失了一部分动量,计算出这部分损失的动量并添加到模型的动量方程中,以实现养殖贝串阻流效应的模拟。动量汇法根据贝串阻流面积、养殖密度、周围水体运动速度等变量计算出损失部分的动量,具体计算公式如下[16-17]:
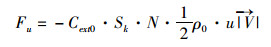 (1)
(1)
 (2)
(2)
式中: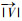

|
表 1 养殖设施组成对应的Sk Tab.1 Sk corresponding to each part of the aquaculture facility |
贻贝养殖海域附生海藻碎屑的运动随着水动力条件的变化而改变,为模拟海藻碎屑的漂移和沉降情况,基于三维水动力模型耦合拉格朗日质点追踪模块对海域海藻碎屑的水平输运轨迹和垂向沉降进行了数值模拟。拉格朗日追踪法计算方程如下[18]:
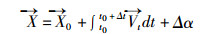 (3)
(3)
式中:


在数值模型中拉格朗日质点随机运动距离采用随机函数算法。质点初始位置(
对建立好的三维水动力模型进行潮位和流速的验证,用于评价模拟与实测结果精度的参数有:技术评分(skill score,SS),相关系数(correlation coefficient,CC),均方根误差(root mean square error,RMSE)。SS划分的评价标准:[0.65, 1)为模型模拟结果优秀;[0.5, 0.65)为优良;[0.2, 0.5)为良好;(0,0.2)为结果差。CC越接近1,RMSE越接近0,表示模拟结果与实测越相近[20-21]。
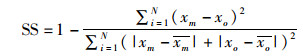 (4)
(4)
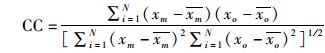 (5)
(5)
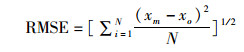 (6)
(6)
式中:xm为模拟结果;xo为实测结果;N为样本量;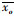
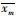
4个站点潮位模拟结果如图 3所示,相位模拟精度高于振幅,各站点评价参数如表 2所示,SS和CC值均大于0.9,RMSE值小于0.2,表明模型潮位模拟较高。

|
图 3 潮位验证结果 Fig. 3 Validation result of elevation |

|
表 2 潮位验证参数 Tab.2 Validation parameters of elevation |
养殖区外部及内部(S3、S4)流速u、v分量验证结果的评价参数如表 3所示,中、下层流速验证结果优于表层,场外验证结果优于场内。篇幅所限,此处仅给出场外(S3)、场内(S4)各1个站点的模拟与实测流速流向的时间过程曲线(图 4,图 5)。最低SS值在0.40以上,平均值0.77,表明本文所建立数值模型的精度优良,可用于后续的碎屑输运动力学研究。

|
表 3 流速验证参数 Tab.3 Validation parameters of current velocity |

|
(a), (b), (c)分别表示表层流速和流向; (d), (e), (f)分别表示中层流速和流向; (g), (h), (i)分别表示底层流速和流向。 (a), (b), (c) represent the velocity and direction of surface layer; (d), (e), (f) represent the velocity and direction of middle layer; (g), (h), (i)represent the velocity and direction of bottom layer. 图 4 S3站点潮流实测与模拟验证结果 Fig. 4 Tidal current velocity validation results of S3 |

|
(a), (b), (c)分别表示表层流速和流向; (d), (e), (f)分别表示中层流速和流向; (g), (h), (i)分别表示底层流速和流向。 (a), (b), (c)represent the velocity and direction ofsurface layer; (d), (e), (f)represent the velocity and direction of middle layer; (g), (h), (i)represent the velocity and direction of bottom layer. 图 5 S4站点潮流实测与模拟验证结果 Fig. 5 Tidal current velocity validation results of S4 |
分别计算了海表有、无阻流效应的2个算法在涨急和落急时刻流场的差异性。海表养殖设施明显降低了养殖海域内的表层流速大小(图 6)。无阻流效应时,枸杞岛西北岸贻贝养殖海域表层(1 m)涨急与落急时刻的流速分别约为1 m/s和0.5 m/s,而有阻流效应时,涨急、落急时的枸杞岛西北岸及北岸养殖区表层(1 m)的流速均约为0.1 m/s,与实测流速接近(图 4, 图 5)。不同于养殖区内部,贝串的阻流效应对枸杞岛西北岸贻贝养殖区外围海域的流速有强化作用(图 6)。

|
图 6 大潮期间涨急、落急流场 Fig. 6 Current field at flood and ebb during the spring |
为了能够确切得知枸杞岛贝串阻流效应对流速的衰减程度,利用有、无阻流效应2个算法的表、中、底流速求出流速相对差,具体计算公式如下:
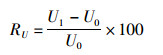 (7)
(7)
式中:U1为有阻流效应时的流速;U0为无阻流效应时的流速;RU为2种条件下的流速相对差, %。利用上式计算出养殖场中心的S4、S5站点不同深度层的RU。结果显示,表层流速平均降低了约80%,中、下层流速分别降低了50%和30%左右。位于表层的养殖设施对垂向整个水体均有阻滞作用,但对表层流速的衰减作用明显大于中下层。
2.3 潮致欧拉余流欧拉余流的计算采用数值模型在58 d(5个主要天文分潮周期的公倍数)内的潮流平均值,得到枸杞岛贻贝养殖海域在海表有、无阻流效应2种情况下的欧拉余流场(图 7),最大潮致欧拉余流可达0.3 m/s。贝串对水流的阻滞作用主要集中表现在削弱了养殖海域内部余流流速。在无阻流效应时枸杞岛附近的海域有很明显的3个潮致余流分支:第1分支位于枸杞岛西北侧养殖海域的内部,由东北方向流向西南,在枸杞岛西侧与第2分支,即由东南流向西北的余流交汇,第3分支位于嵊山岛北侧和东侧,由西向东转而从嵊山岛东侧绕向西南方向,呈顺时针半圆弧型。当施加贝串对水流的阻滞作用时,阻流效应显著削弱了第1分支潮致余流的流速,其大小仅为无阻流效应下的1/30,这是本海域由于养殖设施存在而导致的水动力系统最大的变化,将深刻影响海域的物质输运。养殖设施的存在对枸杞岛西侧第2分支潮致余流略有加强。底层余流的分布态势与表层相同,但量值偏小,仅养殖场内的第1潮致余流分支底层强度大于表层。

|
图 7 表层潮致欧拉余流 Fig. 7 Euler residual flow on the surface layer |
质点分75批释放后,海域质点个数保持不变(图 8),质点总量为61 350个。以质点的输运扩散表示养殖海域附生海藻碎屑脱落后的运动情况。

|
图 8 养殖海域释放质点数量随时间的变化 Fig. 8 Variation of the total amount of released particles with time inside the aquaculture sea area |
附生海藻碎屑从筏架脱落后,在水流带动下向外海扩散。大、小潮的潮动力差异,海表阻流效应及初始沉降速率(w)等均是影响碎屑输运的重要因素。养殖区作为海藻碎屑的源头,其脱落的一小部分碎屑可能直接被养殖贝类滤食并以假粪形式沉降,一部分在水流的带动下输送至养殖区外,还有一部分直接沉降至海底。本文设定当质点所处的深度大于0.98倍的水深时,便认为该碎屑沉落至海底,沉降至海底的碎屑会被底栖生物所摄食或者被微生物分解利用后重新进入生物地球化学循环过程。
当初始沉降速率w为0 m/s时,碎屑受浮力作用很难快速下沉,养殖海域内质点沉底率约1%,大部分质点在水流的带动下输送至养殖区外,致使养殖区外质点沉底率与质点总沉底率变化趋势接近,且随时间呈上升的趋势。在有、无阻流效应两种情况下,质点的沉底率整体变化趋势较为一致,大、小潮期间释放质点的沉底率无明显的差异性(图 9a、图 9 b)。无阻流效应时,当质点的沉底率达到36.79%(e-1)时所需要的时间约为592~600 h(表 4);在有阻流效应的条件下质点达到36.79%的沉底率所需时间略大于无阻流效应。

|
红色线条表示有阻流效应,黑色线条表示无阻流效应; (a), (c), (e), (g): 大潮期间释放; (b), (d), (f), (h): 小潮期间释放。 The red line indicates the blocking effect, the black line indicates the no-blocking effect; (a), (c), (e), (g): released during the spring tides; (b), (d), (f), (h): released during neap tides. 图 9 养殖区内、外大小潮期间质点的沉底率 Fig. 9 Sedimentation rate of seaweed detritus during spring and neap tides, in and outside the culture area |

|
表 4 养殖海域质点达到不同沉底率所耗费的时间 Tab.4 Time for particles to reach different sedimentation rates in the aquaculture sea area |
当初始沉降速率w大于0 m/s时,在前20 h内图质点沉底率的增长最迅速,随后质点沉底率随时间的变化趋于平稳。养殖海域内、外质点总沉底率较为接近,当所有75批质点释放完成后,总沉底率已接近99%(图 9c~图 9h, 表 4)。
有、无阻流效应2种条件下质点均能够实现接近99%的沉底率,所需要的时间约76~82 h(表 4),沉降范围主要集中在距养殖区10 km以内的海域(图 10, 表 5)。有阻流效应的情况下质点完全沉底所耗费的时间略大于无阻流效应情况下质点沉降所用的时间。

|
小圆圈表示距养殖区中心10 km的范围,大圆圈表示距养殖区中心20 km的范围; (a), (e), (i), (m): 大潮释放,无阻流效应;(b), (f), (j), (n): 小潮释放,无阻流效应;(c), (g), (k), (o): 大潮释放,有阻流效应; (d), (h), (l), (p): 小潮释放,有阻流效应。 The small circle indicates the range of 10 km from the center of the culture zone, the large circle indicates the range of 20 km from the center of the culture zone; (a), (e), (i), (m): released during spring tides and without the retarding effect; (b), (f), (j), (n): released during neap tides and without retarding effect; (c), (g), (k), (o): released during spring tides and with retarding effect; (d), (h), (l), (p): released during neap tides and with retarding effect. 图 10 质点沉底时间 Fig. 10 Time of seaweed detritus sinking to the bottom |

|
表 5 养殖海域质点达到不同沉底率时距养殖海域中心的距离 Tab.5 Distance from the center of the aquaculture area when particles reach different settlement rates |
相比大潮,小潮期间水动力条件较弱,质点沉降范围集中在养殖区内部及周边海域(表 5)。海表阻流效应的存在进一步减弱了水体的流动性,缩小了质点的沉降范围,使其更加集中分布在养殖区内部及边缘(表 4)。3种不同质点沉降速度对质点的沉底率、输运范围、沉底所需时长等的差异并不显著(表 4,图 11)。

|
红色表示无阻流效应,蓝色表示有阻流效应。 Red with the retarding effect, blue without the retarding effect. 图 11 M2分潮潮流椭圆 Fig. 11 Tidal ellipse of M2 |
由上述结果可知,当质点具有固定沉降速度时便能在较短时间内实现全部沉降(图 10),然而沉降至海底的碎屑所处的位置有较大差异,主要表现在有、无阻流效应2种情况下和大、小潮期间释放造成的质点空间分布范围的差异性。通过确定达到某一沉底率时质点距养殖区中心的范围可以判断阻流效应和大、小潮对质点水平输送的影响。在无阻流效应时,当质点沉底率达到99%,大、小潮期间释放的质点分布范围距离养殖海域中心分别约为10.5~13.5 km,10.0~11.5 km,高于有阻流效应条件下质点位置距养殖海域中心的距离(表 5)。在养殖设施海表阻流效应的作用下,大潮期间释放质点的平均输送距离较无阻流效应时减少了约1.6 km, 小潮期间释放质点的输送距离减少了约2.8 km。
3 讨论 3.1 贝串阻流效应对潮流的影响涨潮时,来自东海东南部海域的潮流在流经嵊山岛时被分为2股,一股沿嵊山岛北部向西流,在流经枸杞岛北侧养殖海域时由于贻贝串的阻挡作用,大部分海流沿着养殖区外侧流向西北,养殖场过流断面的表层流速和流量大幅减小,从而使得养殖场外围流速加强。另一股潮流沿嵊山南部流向枸杞岛西部,由于养殖贝串的滞流作用,枸杞岛西北岸海域的流速减弱,使得这支海流得以继续向枸杞岛西北侧运动,后与枸杞岛北岸自东向西的涨潮流汇合后流向西北。落急时,同样由于贝串的阻碍作用,枸杞岛西北岸外围海流的过流面减小,致使流速加强。因此,贝串在减弱养殖海域内部流速的同时会增大养殖区外围的流速。
一般地,养殖活动对水流阻碍作用的强弱与养殖对象相关。当贻贝处于生长阶段的初期时,贝串柔性较高、质量较轻,横截阻流面积小;当贻贝长成时贝串粗重、柔性变形弱、不易随水流摆动,形成的阻流效应增大。海带、海藻等一些植物柔性较高,产生的阻流效应较弱,即使在相对封闭的海域环境中养殖其对流速的影响也相对较弱[22]。除养殖对象外,养殖区流速的变化率与养殖区所在的位置也有关。枸杞岛西北岸贻贝养殖区三面靠海,处于半开放海域,自然条件下水交换能力较好,当受到大规模养殖浮筏阻流效应时流速衰减明显;而位于内湾的部分养殖海域自然条件下水动力较弱,养殖活动阻流效应相对不显著[23],但养殖海域的整体阻流效应对局部内湾水交换能力的影响往往更严重。
因此,在制定养殖策略时应该考虑当地自然条件下水动力学特性,并据此调整养殖密度、贝串的长度,以尽可能降低对流场的影响,保证良好的水交换能力。
3.2 水动力条件的强弱对附生海藻碎屑输运的影响海藻碎屑是浅海生态系统中海洋生物重要的食物来源之一,不但能够被底栖次级生产者所利用,而且分解后能释放大量的颗粒有机物和溶解有机物质,促进海区的碳循环,提高初级生产力[24]。因此,海藻碎屑的空间分布对海洋生态系统至关重要。在不考虑海面风应力作用的前提下,潮致余流是影响海水中物质输运的重要动力因子(图 7、图 10),而海藻碎屑输运的主要运动轨迹基本与余流流向一致。此外,枸杞岛附近海域的M2分潮的潮流椭圆的长轴方向也一定程度上指示着海藻碎屑的输运方向(图 11)。水动力条件的强弱对海藻碎屑沉底率高低的影响相对较小,而海藻碎屑有、无初始沉降速率是影响其沉底率高低的关键[25]。
3.3 后续研究展望拉格朗日质点的运动轨迹表示了养殖区海藻碎屑输运过程的空间分布,碎屑被微生物分解后又参与到生态系统中的物质循环,海藻碎屑大范围的输运沉降能够扩大附生海藻的生态辐射范围。本文的不足之处主要在于通过释放质点的方式探究养殖海域碎屑的运动,但未能把海藻碎屑的生源物质含量与质点间建立数量关系。因此,在今后需要结合生物地球化学研究,从固碳效应及其生态辐射强度与范围的角度开展后续研究。
此外,水体中海藻碎屑的输运同时受到潮流、风、斜压效应等动力因素的影响。而本文仅考虑了潮流作用下枸杞岛贻贝养殖海域海表阻流效应对海藻碎屑水平及垂向输运的影响。事实上,枸杞岛附近海域的年平均风速能达到7.2 m/s,年平均≥6级风的大风日数为152.4 d[26]。海表风对海面的摩擦力可能促进或阻碍水体的流动。在夏季枸杞岛附近海域易受热带气旋的影响,年均台风频数约3个[26],台风前后大型藻类的生物量也会发生脱落等受到影响。因此,在后续研究中应考虑附生海藻大量脱落季节,季节风对养殖海域大型海藻输运的影响。
养殖设施的阻流效应除了使养殖区边缘流速增大(图 6、图 7)、使潮流椭圆变形之外(图 11),表层4 m层内的阻流效应也可能加速质点的沉降。质点沉降速度为0时,养殖场内质点沉底率在有阻流效应条件下显著高于无阻流效应(图 9a, 图 9b),质点沉降速度w=0.055 m/s的结果(图 9g, 图 9h)也支持这一观点,但其他2个沉降速度组的结果不明显、甚至相反。后续将对养殖场内不同水层的颗粒物沉降速度开展更深入的现场观测和数值模拟研究。
| [1] |
李训猛, 章守宇, 王凯, 等. 周年温度变化对枸杞岛铜藻生长特性的影响[J]. 海洋与湖沼, 2020, 51(5): 1136-1143. LI X M, ZHANG S Y, WANG K, et al. Effect of annual temperature variation on growth of Sargassum horneri in GouqiIsland[J]. Oceanologiaet Limnologia Sinica, 2020, 51(5): 1136-1143. |
| [2] |
常抗美, 吴剑锋. 厚壳贻贝人工繁殖技术的研究[J]. 南方水产, 2007, 3(3): 26-30. CHANG K M, WU J F. Study on artificial propagation of mussel Mytiluscoruscus[J]. South China Fisheries Science, 2007, 3(3): 26-30. DOI:10.3969/j.issn.2095-0780.2007.03.005 |
| [3] |
徐雯佳. 基于高分卫星影像的秦皇岛近海浮筏养殖分布遥感监测[J]. 河北渔业, 2020(4): 32-34, 63. XU W J. Remote sensing monitoring of floating raft culture distribution in coastal area of Qinhuangdao based on high-resolution satellite images[J]. Hebei Fisheries, 2020(4): 32-34, 63. DOI:10.3969/j.issn.1004-6755.2020.04.011 |
| [4] |
MCKIFDSEY C W, ARCHAMBAULT P, CALLIER M D, et al. Influence of suspended and off-bottom mussel culture on the sea bottom and benthic habitats: A review[J]. Canadian Journal of Zoology, 2013, 89: 622-646. |
| [5] |
刘书荣, 周曦杰, 章守宇, 等. 贻贝筏式养殖区附生大型海藻与两种附着端足目的关系[J]. 生态学杂志, 2018, 37(9): 2737-2744. LIU S R, ZHOU X J, ZHANG S Y, et al. Relationship between epiphytic seaweeds and two seaweed-associated amphipods in mussel raft culture area[J]. Chinese Journal of Ecology, 2018, 37(9): 2737-2744. DOI:10.13292/j.1000-4890.201809.015 |
| [6] |
BRICKHILL M J, LEE S Y, CONNOLLY R M. Fishes associated with artificial reefs: attributing changes to attraction or production using novel approaches[J]. Journal of Fish Biology, 2015, 67(SB): 53-71. |
| [7] |
BOYRA A, SANCHEZ-JEREZ P, TUYA F, et al. Attraction of wild coastal fishes to an Atlantic subtropical cage fish farms, Gran Canaria, Canary islands[J]. Environmental Biology of Fishes, 2004, 70(4): 393-401. DOI:10.1023/B:EBFI.0000035435.51530.c8 |
| [8] |
骆其君. 象山港海域中定点漂浮物的大型海藻演变[J]. 宁波大学学报(理工版), 2009, 22(4): 467-471. LUO Q J. Change of macroalgae in fixed floats in Xiangshanbay[J]. Journal of Ningbo University (NSEE), 2009, 22(4): 467-471. DOI:10.3969/j.issn.1001-5132.2009.04.005 |
| [9] |
章守宇, 孙宏超. 海藻场生态系统及其工程学研究进展[J]. 应用生态学报, 2007, 18(7): 1647-1653. ZHANG S Y, SUN H C. Research progress on seaweed bed ecosystem and its engineering[J]. Chinese Journal of Applied Ecology, 2007, 18(7): 1647-1653. DOI:10.3321/j.issn:1001-9332.2007.07.038 |
| [10] |
徐永健, 钱鲁闽, 韦玮, 等. 两种大型海藻自然环境下的营养动力学研究[J]. 海洋环境科学, 2007, 26(2): 161-165. XU Y J, QIAN L M, WEI W, et al. Studies on nutrient kinetics characteristics of two species seaweeds (Rhodophyta) at outdoor natural conditions[J]. Marine Environmental Science, 2007, 26(2): 161-165. DOI:10.3969/j.issn.1007-6336.2007.02.015 |
| [11] |
李秀辰, 谷晓华, 张国琛, 等. 石莼属海藻的环境增殖及生物质能开发潜力[J]. 水产科学, 2011, 30(12): 789-793. LI X C, GU X H, ZHANG G C, et al. Environment-enhancing and bio-energy producing potential of Ulva sp.[J]. Fisheries Science, 2011, 30(12): 789-793. DOI:10.3969/j.issn.1003-1111.2011.12.015 |
| [12] |
杨宇峰, 宋金明, 林小涛, 等. 大型海藻栽培及其在近海环境的生态作用[J]. 海洋环境科学, 2005, 24(3): 77-80. YANG Y F, SONG J M, LIN X T, et al. Seaweed cultivation and its ecological roles in coastal waters[J]. Marine Environmental Science, 2005, 24(3): 77-80. |
| [13] |
EGBERT G D, EROFEEVA S Y. Efficient inverse modeling of barotropicocean tides[J]. Journal of Atmospheric and Oceanic Technology, 2002, 19(2): 183-204. DOI:10.1175/1520-0426(2002)019<0183:EIMOBO>2.0.CO;2 |
| [14] |
LIN J, LIC Y, ZHANG S Y. Hydrodynamic effect of a large offshore mussel suspended aquaculture farm[J]. Aquaculture, 2016, 451: 147-155. DOI:10.1016/j.aquaculture.2015.08.039 |
| [15] |
SHI J, WEI H, ZHAO L, et al. A physical-biological coupled aquaculture model for a suspended aquaculture area of China[J]. Aquaculture, 2011, 318(3/4): 412-424. |
| [16] |
YANG Z Q, WANG T P, COPPING A E. Modeling tidal stream energy extraction and its effects on transport processes in a tidal channel and bay system using a three-dimensional coastal ocean model[J]. Renewable Energy, 2013, 50: 605-613. DOI:10.1016/j.renene.2012.07.024 |
| [17] |
LI X R, LI M, MCLELLAND S J, et al. Modelling tidal stream turbines in a three-dimensional wave-current fully coupled oceanographic model[J]. Renewable Energy, 2017, 114: 297-307. DOI:10.1016/j.renene.2017.02.033 |
| [18] |
范世平, 王彦芳, 冯民权, 等. 污染物迁移扩散的质点追踪随机模拟[J]. 武汉大学学报(工学版), 2011, 44(1): 32-36. FAN S P, WANG Y F, FENG M Q, et al. Particle tracing random simulation on advection diffusion of pollutants[J]. Engineering Journal of Wuhan University, 2011, 44(1): 32-36. |
| [19] |
章守宇, 向晨, 周曦杰, 等. 枸杞岛海藻场6种大型海藻光合荧光特性比较[J]. 应用生态学报, 2018, 29(10): 3441-3448. ZHANG S Y, XIANG C, ZHOU X J, et al. Photosynthetic fluorescence characteristics of six macroalgae species in seaweed beds of Gouqi Island, Zhejiang, China[J]. Chinese Journal of Applied Ecology, 2018, 29(10): 3441-3448. |
| [20] |
林军, 吴星辰, 杨伟. 潮流作用下人工鱼礁山海域泥沙输运的数值模拟研究[J]. 水产学报, 2020, 44(12): 2087-2099. LIN J, WU X C, YANG W. Numerical modelling of sediment transport in artificial seamount by the action of tidal current[J]. Journal of Fisheries of China, 2020, 44(12): 2087-2099. |
| [21] |
LIN J, LI C Y, BOSWELL K M, et al. Examination of winter circulation in a northern gulf of Mexico estuary[J]. Estuaries and Coasts, 2016, 39(4): 879-899. DOI:10.1007/s12237-015-0048-y |
| [22] |
JACKSON G A, WINANT C D. Effect of a kelp forest on coastal currents[J]. Continental Shelf Research, 1983, 2(1): 75-80. |
| [23] |
史洁, 魏皓. 半封闭高密度筏式养殖海域水动力场的数值模拟[J]. 中国海洋大学学报(自然科学版), 2009, 39(6): 1181-1187. SHI J, WEI H. Simulation of hydrodynamic structures in a semi-enclosed bay with dense raft-culture[J]. Periodical of Ocean University of China, 2009, 39(6): 1181-1187. |
| [24] |
申玉春, 熊邦喜, 叶富良, 等. 虾-鱼-贝-藻生态优化养殖及其水质生物调控技术研究[J]. 生态学杂志, 2005, 24(6): 613-618. SHEN Y C, XIONG B X, YE F L, et al. Prawn-fish-shellfish-algae optimal ecoculture and its bio-manipulation technique of water quality[J]. Chinese Journal of Ecology, 2005, 24(6): 613-618. |
| [25] |
桂福坤, 方帅, 曲晓玉, 等. 舟山近海海域养殖水体悬浮物沉降特性试验研究[J]. 农业工程学报, 2020, 36(10): 206-212. GUI F K, FANG S, QU X Y, et al. Experimental study on settling characteristics of suspended solids in seawater of nearshorein Zhoushan of China[J]. Transactions of the Chinese Society of Agricultural Engineering, 2020, 36(10): 206-212. |
| [26] |
吴祖立, 章守宇. 台风对浙江枸杞岛大型底栖海藻分布的影响分析[J]. 中国农业科技导报, 2019, 21(9): 159-168. WU Z L, ZHANG S Y. Effect of typhoon on the distribution of macroalgae in the seaweed beds of Gouqi Island, Zhejiang province[J]. Journal of Agricultural Science and Technology, 2019, 21(9): 159-168. |
2. Engineering Technology Research Center of Marine Ranching, Shanghai Ocean University, Shanghai 201306, China;
3. East China Sea Forecasting Center, State Oceanic Administration, Shanghai 200136, China