 2022,
Vol. 31
2022,
Vol. 31



2. 大洋渔业资源可持续开发教育部重点实验室, 上海 201306;
3. 中国水产科学研究院南海水产研究所, 广东 广州 510300
近年来,基于生态系统的渔业管理(ecosystem-based fisheries management,EBFM)已成为国际渔业科学领域研究的一个新趋势[1]。EBFM的前提是认识生态系统结构和功能,而食物网是研究生态系统结构和功能的基础[2-3]。食物网特征研究是通过物种之间的营养相互作用来描述和量化生态系统结构和复杂性的一种方法[4],对于理解生态系统至关重要。研究生态系统动力学需要定量描述食物网结构[5]。生态网络分析(ecological network analysis,ENA)可以量化生态系统内物种的相互作用[6],同时可以分析这种相互作用如何间接影响整个生态系统[7],因此已被用于海洋生态系统及其食物网结构的定量研究[8-10]。
热带太平洋东部海域具有较高的生物多样性和丰富的鱼类资源[11],也是世界金枪鱼和类金枪鱼的重要栖息海域。因金枪鱼和类金枪鱼具有很高的营养价值和商业价值,吸引了世界各国前往热带太平洋东部海域进行金枪鱼延绳钓作业。热带太平洋的金枪鱼渔业已有70多年历史,随着金枪鱼资源的不断开发,金枪鱼渔业资源的可持续利用引起了有关区域管理和国际组织的高度关注和重视[12]。VASSLIDES等[13]研究发现,长期的渔业捕捞会影响海洋生态系统的平衡。随着渔业发展,顶级捕食物种的丰度大幅下降,可能导致大型捕食动物甚至整个生态系统的营养关系发生较大变化[14-16]。
本文根据我国金枪鱼渔业科学观察员随生产航次观测的渔获物及其胃含物资料,结合文献记载资料,以商业性渔业长期捕捞的经济鱼种和兼捕物种为主要对象,构建热带东太平洋中上层食物网,分析食物网网络结构和复杂性指数,以期为热带太平洋东部海域食物网功能的深入研究及基于生态系统的渔业管理提供参考。
1 材料与方法 1.1 样品采集摄食数据主要来源于我国金枪鱼科学观察员随生产渔船采集的渔获物胃样品,同时通过查阅相关历史文献[17]、世界鱼类数据库网站(https://www.fishbase.de/)对部分物种的摄食数据进行了补充。生产渔船共计6个航次,时间为2017年6月—2019年2月,6个航次作业的区域为14°03′N~30°22′S和179°36′E~114°29′W,详见表 1。渔船为超低温金枪鱼延绳钓渔船,观察员对钓获的渔获物测定体长和体质量等生物学信息后,进行现场解剖,取出整胃置于10%的甲醛溶液中并冷冻保存。共计从全部渔获物中随机采集280个胃样品(表 2),带回实验室进行胃含物分析。

|
表 1 观察员航次采样信息 Tab.1 Sampling information of the observer trips |

|
表 2 观察员采集的胃样品种类及数量 Tab.2 Species and number of stomach samples collected by observers |
胃含物样品解冻后进行生物学测定,记录长度(精确至0.1 mm)和质量(精确至0.01 g)等信息。参照图鉴[18]对胃含物样品中形态较完整的个体,在体视显微镜和解剖镜下直接进行物种鉴定。对肉眼难以鉴别的物种,取其肌肉放入装有体积分数为95%的乙醇溶液的离心管中冷冻保存,或取其耳石放入装有蒸馏水的离心管中,于4 ℃冰箱中保存,后续进行DNA分析和耳石形态分析来鉴定种类。尽量将胃含物种类鉴定到最低分类单元。少数胃含物样品消化较完全,难以鉴定,则归纳为未知物种。
1.3 网络指数分析为构建尽可能详细的食物网,在实验数据之外,还查阅了相关历史文献[17]以及世界鱼类数据库网站(https://www.fishbase.de/)进行数据补充。将整理获得的数据,构建成由S行和S列组成的矩阵,其中S是研究中所涉及的物种与类群的总数,物种的顺序在行和列之间是相同的。在食物网矩阵中第i行和第j列的交叉点上的值为0或1,1表示捕食者j捕食了第i种猎物,0则表示i和j不存在摄食关系[19]。使用NetDraw软件从构造的矩阵中绘制食物网图。
运用12种常见的网络指数来描述食物网结构特征和复杂性[19-22],包括:物种与类群总数量(species, S)、摄食关系形成的连接数(links, L)、连接密度(linkage density, D)、连通度(connectance, Cl)、顶级物种百分比(top species, T)、中间物种百分比(intermediate species, I)、基础物种百分比(basal species, B)、杂食物种百分比(omnivorous, O)、连接复杂性指数(connection complexity index, CSC),计算公式为
 (1)
(1)
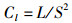 (2)
(2)
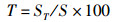 (3)
(3)
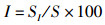 (4)
(4)
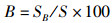 (5)
(5)
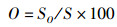 (6)
(6)
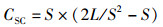 (7)
(7)
式中:D为连接密度;Cl为连通度;S为物种与类群总数量;L为连接数;T、I、B、O分别为食物网中顶级物种、中间物种、基础物种、杂食物种的百分比,%;ST、SI、SB、SO分别为顶级物种、中间物种、基础物种和杂食物种的数量;CSC为连接复杂性指数。
利用WILLIAMS等[23]的短加权营养级公式计算了物种的营养级(trophic level, TTL)。短加权营养级定义为最短营养级和猎物平均营养级的平均值。食物网中一个捕食者的最短营养级等于从这个捕食者到任何基础种的最短链长加1[23]。猎物平均营养级等于所有捕食者所捕食的猎物的平均营养级再加1,计算公式为
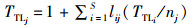 (8)
(8)
式中:TTLj、TTLi分别为j物种和i物种的营养级;S是食物网中物种与类群的总数;对于列j和行i,lij是S行和S列的连接矩阵,如果物种j捕食物种i,lij为1,如果物种j不捕食物种i,lij为0;nj为j种摄食被捕食物种的数量。因此,短加权营养级产生一个营养级的最小估计值,并假设基础物种的营养级为1.0[23]。在本研究中我们将食物网的平均营养级(MT)看作是所有物种营养级的平均值。
特征路径长度(characteristic path length, P),所有物种对之间的平均最短路径长度,公式如下:
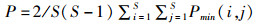 (9)
(9)
式中: ∑i=1S∑j=1SPmin(i, j)表示任意物种对之间的最短路径长度;2/S(S-1)表示节点数。
聚类系数(clustering coefficient,CC),连接到相同物种和彼此相互连接的物种对的平均数量,以此来描述食物网中物种节点的聚集程度。
2 结果胃含物样品中共采集热带太平洋东部海域中上层食物网中50个物种和33个类群(表 3)。食物网结构如图 1,图中不同物种节点的大小代表每个物种的连接数(包括捕食与被捕食),箭头方向代表从被捕食者到捕食者。连接数(L)超过10的物种(类群)有33种,占总物种数的40%(表 3)。L较高的物种(类群)为黄鳍金枪鱼(Thunnus albacares, 54)、大眼金枪鱼(Thunnus obesus, 53)、头足类(Cephalopoda, 184)和甲壳类(Crustacea, 54),其中黄鳍金枪鱼和大眼金枪鱼为太平洋延绳钓渔业中的重要经济物种,在食物网中主要扮演捕食者角色;头足类和甲壳类为主要经济物种的重要摄食对象。L最低的为白边真鲨(Carcharhinus albimarginatus,3),这可能与白边真鲨的主要摄食数据缺失有关。该食物网在83个物种与类群之间共有542条连接数,物种的连接密度为6.53,连通度为0.08(表 4)。

|
表 3 热带太平洋东部海域中上层食物网物种及其连接数 Tab.3 Species and trophic links of pelagic food web in the tropical eastern Pacific Ocean |

|
节点大小代表物种间的连接数量,箭头方向代表物种间的捕食关系。 Size of nodes represents the number of links between species and the direction of arrows represents the feeding relationship from the prey to the predator. 图 1 热带东太平洋海域中上层食物网结构(编号1~83见表 3) Fig. 1 Trophic structure of pelagic food web in the tropical eastern Pacific Ocean(See ID 1-83 in Tab. 3) |

|
表 4 热带东太平洋海域中上层食物网结构指数 Tab.4 Structural indices of pelagic food web in the tropical eastern Pacific Ocean |
顶级物种为只捕食猎物的物种,即营养链只有连入的物种,其占总物种数的10.85%;基础物种为仅作为被捕食者的物种,即营养链只有连出的物种,其占总物种数的6.02%;中间物种为既是捕食者又是被捕食者的物种,即营养链同时有连入和连出的物种,其占总物种数的83.13%(表 4)。中间物种数量远超过顶级物种和基础物种,即大多数物种既是捕食者又是被捕食者。杂食性物种的数量为总数量的92.77%,处于较高水平。食物网的平均营养级(MT)为2.4,特征路径长度为1.85,聚类系数为0.242(表 4)。
3 讨论 3.1 食物网稳定性分析本食物网结构与其他海洋食物网结构相比存在一定差异(表 5)。与极地海洋食物网(波特湾、北极、南极食物网)相比,本食物网与其他非极地海洋食物网一样,基础物种分类到种的比例较低。因为基础物种鉴定较为困难,这在其他食物网研究中也较为常见[19]。在本食物网中,中间物种和杂食性物种的比例高于其他海洋食物网(表 5)。在食物网中,中间物种和物种杂食性水平通常具有相关性,因为一般情况下,大多数既作为捕食者又作为猎物的中间物种会跨越多个营养级进行摄食,属于杂食性物种[19]。NORKKO等[24]的研究表明,物种的杂食性对食物网的稳定性具有积极作用,因为它可能对受到时间和空间限制且不可预测的食物来源做出更迅速、更灵活的响应。因此,杂食性指数越高,食物网越稳定[24]。本食物网杂食性指数为92.77%,与其他的海洋食物网相比处于较高位置(表 5),这说明该食物网具有较高的稳定性。

|
表 5 热带东太平洋中上层食物网与其他水域食物网指数的对比 Tab.5 Comparison of food web indices of the tropical eastern Pacific Ocean with other aquatic food webs |
DUNNE等[20]研究表明,海洋食物网的特征路径长度的正常范围为1.3~3.7。本食物网的特征路径长度为1.85,说明该食物网的连接路线长度处于正常范围内,食物网可在一定程度上抵御外界干扰。但是,需要注意的是,该食物网的特征路径长度与稳健食物网[25]相比偏低,这说明如果该生态系统受到外界持续剧烈的干扰,例如系统内的主要经济鱼种遭受过度捕捞,食物网中的其他物种也会因此受到负面影响[26],进而影响生态系统的维稳性。
3.2 食物网营养级分析本食物网的平均营养级(2.4)相对较低,最大营养级为4.15,为虎鲸(Orcinus orca),低于大多数其他的食物网[20, 34],这表明虎鲸和底部物种之间的营养链较短,可能原因是本研究数据中部分小型鱼类的饵料数据不够完善。另外,本食物网中,重要经济鱼种,例如黄鳍金枪鱼、大眼金枪鱼、鲣(Katsuwonus pelamis)的营养级分别为3.16、3.08、2.99,在另一关于热带东太平洋食物网的研究中[35],3个物种的营养级分别为4.57、4.53、4.57。本研究中重要经济鱼种的营养级较低的原因,一方面可能是本研究的食物网缺失部分初级生产者信息;另一方面本研究未考虑不同生活史阶段鱼类的摄食习性变化,即饵料生物的比例变化对食物网营养结构的影响。由于热带东太平洋金枪鱼延绳钓渔业中涉及的渔获物体型较大,重要经济鱼种在相对资源量统计中并未区分个体大小[36],从而可能产生误差。CORTÉS[37]认为还需要应用综合指数,如相对重要性指数(IRI)、物种生物量等对营养级进行修正计算。但事实上,相对于近海,大洋性生态系统范围广,取样困难,基于渔业生产的取样仍是目前国际上常用的方法。另外,对食物中的饵料进行分类已非常困难,要获得饵料质量的难度更大。大多数胃含物分析结果只是反映了物种最后一次进食的状况,饵料在胃内能保持完整的个体较少,将所有的饵料残肢都换算成个体的更正质量难度也较大[38]。此外,本样品采集未考虑季节变化,未来需要尽可能实现该海域一年四季的采样调查,以研究摄食的季节变化。
3.3 食物网复杂性分析食物网的复杂性主要通过连接密度(D)和连通度(Cl)来分析[19],D和Cl的值越大,食物网复杂性越高[39]。在本食物网中,D和Cl的值分别为6.53和0.08。食物网的连接密度和连通度过高和过低都是非正常现象[20],通过对比其他水域食物网(表 5)以及其他类型食物网[20]发现,本食物网指标的数值处于中间位置。生态网络中D和Cl值的高低,对深入探究生态系统稳定性至关重要。现实中, 热带东太平洋生态系统内的物种数量(S)通常大于实验确定的值,未来需要对热带东太平洋食物网的复杂性开展长期的跟踪调查,提高对摄食种类的鉴定准确性,以期深入研究热带东太平洋食物网的复杂性及其动态变化。
本食物网的连接复杂性指数(CSC)为13.22(表 4),与其他水域食物网相比(表 5),处于中间位置。该食物网的连接复杂性指数表明物种之间的连接较为紧密,可维持食物网结构的稳定性。种间相互作用可用聚类系数来阐释,如果聚类系数较低,则说明食物网中大部分物种间的联系是相似的,即没有较强的种间相互作用[19]。本食物网的聚类系数为0.242,与其他水域食物网相比(表 5),处在中间水平。聚类系数表明本食物网中的物种在一定程度上能够互相联系,形成几个连接性较密切的子网络结构。其中,连接性较高的物种是黄鳍金枪鱼、大眼金枪鱼、帆蜥鱼(Alepisaurus ferox)、剑鱼(Xiphias gladius)和鲯鳅(Coryphaena hippurus), 见表 3。这些杂食性物种在本食物网中具有广泛的生态位,可以跨多个营养级进行摄食,并存在同类相食现象,对食物网中其他物种的种间相互作用有较大影响。同时有研究[40]证明,这些物种是热带太平洋东部海域生态系统的关键物种。如果这些物种被过度捕捞,导致生态系统生物多样性丧失等扰动,可能对整个食物网产生至关重要的影响[19]。
3.4 食物网研究的局限性本研究中的样品主要通过科学观察员获得,样品采集存在一定局限性,一方面延绳钓渔船的主捕对象为金枪鱼与类金枪鱼等高经济价值鱼种,渔具对不同鱼种的选择性不同,一定程度上限制了调查取样的随机性、全面性;另一方面,延绳钓渔船主要捕捞栖息于50~300 m水深的高营养级物种,导致研究对热带东太平洋海域内低营养级的物种涉及较少,且在实验过程中部分饵料物种因消化较完全无法鉴定至最低分类单元,未知鱼类和头足类比例略高,这可能会低估S与L的值,使网络指数的计算产生一定误差。因此,本研究中的食物网只代表热带东太平洋中上层食物网的信息,是基于热带东太平洋食物网的基础和试探性研究,以期为热带东太平洋整个生态系统研究提供参考。
综合食物网结构和复杂性的指数分析表明,热带东太平洋海域中上层食物网结构稳定性较高,复杂性处于正常水平,对人为扰动以及环境变化存在一定的抵御性,从而保证生态系统功能的正常运行。后续需要不断开展取样和调查研究,进一步完善物种捕食数据,以构建更接近现实的食物网。通过对热带东太平洋海域中上层食物网结构与复杂性的研究,有助于进一步加深我们对其生态系统的认识,能够为生态系统功能的深入研究以及渔业资源的科学管理提供参考。
| [1] |
金显仕, 窦硕增, 单秀娟, 等. 我国近海渔业资源可持续产出基础研究的热点问题[J]. 渔业科学进展, 2015, 36(1): 124-131. JIN X S, DOU S Z, SHAN X J, et al. Hot spots of frontiers in the research of sustainable yield of Chinese in shore fishery[J]. Progress in Fishery Sciences, 2015, 36(1): 124-131. |
| [2] |
朱江峰, 戴小杰, 王学昉, 等. 海洋食物网拓扑学方法研究进展[J]. 渔业科学进展, 2016, 37(2): 153-159. ZHU J F, DAI X J, WANG X F, et al. A review of methodology in Marie food-web topology[J]. Progress in Fishery Sciences, 2016, 37(2): 153-159. |
| [3] |
YENJ D L, CABRAL R B, CANTOR M, et al. Linking structure and function in food webs: maximization of different ecological functions generates distinct food web structures[J]. Journal of Animal Ecology, 2016, 85(2): 537-547. DOI:10.1111/1365-2656.12484 |
| [4] |
BASCOMPTE J. Disentangling the web of life[J]. Science, 2009, 325(5939): 416-419. DOI:10.1126/science.1170749 |
| [5] |
ALLESINA S, ALONSO D, PASCUAL M. A general model for food webstructure[J]. Science, 2008, 320(5876): 658-661. DOI:10.1126/science.1156269 |
| [6] |
FATH B D, SCHARLER U M, ULANOWICZ R E, et al. Ecological network analysis: network construction[J]. Ecological Modelling, 2007, 208(1): 49-55. DOI:10.1016/j.ecolmodel.2007.04.029 |
| [7] |
BROSE U, DUNNE J A. Modelling the dynamics of complex food webs[M]//VERHOEF H A, MORIN P J. Community Ecology: Processes, Models, and Applications. Oxford: Oxford University Press, 2009: 37-44.
|
| [8] |
BAIRD D, ASMUS H, ASMUS R. Trophic dynamics of eight intertidal communities of the Sylt-Rømø Bight ecosystem, northern Wadden Sea[J]. Marine Ecology Progress Series, 2007, 351: 25-41. DOI:10.3354/meps07137 |
| [9] |
ULANOWICZ R E. Quantitative methods for ecological network analysis and its application to coastal ecosystems[J]. Treatise on Estuarine and Coastal Science, 2011, 9: 35-57. |
| [10] |
HEYMANS J J, COLL M, LIBRALATO S, et al. Global patterns in ecological indicators of marine food webs: a modelling approach[J]. PLoS One, 2014, 9(4): e95845. DOI:10.1371/journal.pone.0095845 |
| [11] |
朱江峰, 王嗣鑫. 热带太平洋中南部上层鱼类的种间关系研究[J]. 海洋湖沼通报, 2015(2): 9-15. ZHU J F, WANG S X. Interspecific association for pelagic species in the central and southern area of Tropical Pacific Ocean[J]. Transactions of Oceanology and Limnology, 2015(2): 9-15. DOI:10.13984/j.cnki.cn37-1141.2015.02.002 |
| [12] |
许柳雄, 戴小杰. 国际金枪鱼渔业管理趋势及对我国发展金枪鱼渔业的影响[J]. 海洋渔业, 2003, 25(3): 109-111, 119. XU L X, DAI X J. Management trends of international tuna fisheries and the impact on the development of tuna fisheries in China[J]. Marine Fisheries, 2003, 25(3): 109-111, 119. DOI:10.3969/j.issn.1004-2490.2003.03.001 |
| [13] |
VASSLIDES J M, JENSEN O P. Quantitative vs. semiquantitative ecosystem models: comparing alternate representations of an estuarine ecosystem[J]. Journal of Coastal Research, 2017, 78(s1): 287-296. |
| [14] |
PAULY D, CHRISTENSEN V, DALSGAARD J, et al. Fishing down marine food webs[J]. Science, 1998, 279(5352): 860-863. DOI:10.1126/science.279.5352.860 |
| [15] |
COX S P, ESSINGTON T E, KITCHELL J F, et al. Reconstructing ecosystem dynamics in the central Pacific Ocean, 1952-1998: Ⅱ. A preliminary assessment of the trophic impacts of fishing and effects on tuna dynamics[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2002, 59(11): 1736-1747. DOI:10.1139/f02-138 |
| [16] |
POLOVINA J J, ABECASSIS M, HOWELL E A, et al. Increases in the relative abundance of mid-trophic level fishes concurrent with declinesin apex predators in the subtropical North Pacific, 1996—2006[J]. Fishery Bulletin, 2009, 107(4): 523-531. |
| [17] |
刘攀, 戴小杰, 王杰, 等. 热带西太平洋帆蜥鱼胃含物组成分析[J]. 南方水产科学, 2019, 15(1): 20-30. LIU P, DAI X J, WANG J, et al. Composition of stomach contents of longnose lancet fish(Alepisaurus ferox) in western tropical Pacific Ocean[J]. South China Fisheries Science, 2019, 15(1): 20-30. |
| [18] |
陈大刚, 张美昭. 中国海洋鱼类[M]. 中国海洋大学出版社, 2016. CHEN D G, ZHANG M Z. Marine fishes of China[M]. China Ocean University Press, 2016. |
| [19] |
MARINA T I, SALINAS V, CORDONE G, et al. The food web of potter cove (Antarctica): complexity, structure and function[J]. Estuarine, Coastal and Shelf Science, 2018, 200: 141-151. DOI:10.1016/j.ecss.2017.10.015 |
| [20] |
DUNNE J A, WILLIAMS R J, MARTINEZ N D. Food-web structure and network theory: the role of connectance and size[J]. Proceedings of the National Academyof Sciences of the United States of America, 2002, 99(20): 12917-12922. DOI:10.1073/pnas.192407699 |
| [21] |
ALBERT R, BARABÁSIA L. Statistical mechanics of complex networks[J]. Reviews of Modern Physics, 2002, 74(1): 47-97. DOI:10.1103/RevModPhys.74.47 |
| [22] |
MONTOYA J M, SOLÉR V. Small world patterns in food webs[J]. Journal of Theoretical Biology, 2002, 214(3): 405-412. DOI:10.1006/jtbi.2001.2460 |
| [23] |
WILLIAMS R J, MARTINEZ N D. Limits to trophic levels and omnivory in complex food webs: theory and data[J]. The American Naturalist, 2004, 163(3): 458-468. DOI:10.1086/381964 |
| [24] |
NORKKO A, THRUSH S F, CUMMINGS V J, et al. Trophic structure of coastal Antarctic food webs associated with changes in sea ice and food supply[J]. Ecology, 2007, 88(11): 2810-2820. DOI:10.1890/06-1396.1 |
| [25] |
徐从军, 刘阳, 程远, 等. 基于拓扑网络研究海州湾食物网结构与复杂性[J]. 海洋学报, 2020, 42(4): 47-54. XU C J, LIU Y, CHENG Y, et al. Structure and complexity of Haizhou Bay food web based on topological network analysis[J]. Acta Oceanologica Sinica, 2020, 42(4): 47-54. DOI:10.3969/j.issn.0253-4193.2020.04.006 |
| [26] |
DUNNE J A, WILLIAMS R J, MARTINEZ N D. Network structure and biodiversity loss in food webs: robustness increases with connectance[J]. Ecology Letters, 2002, 5(4): 558-567. DOI:10.1046/j.1461-0248.2002.00354.x |
| [27] |
BODINI A, BELLINGERI M, ALLESINA S, et al. Using food web dominator trees to catch secondary extinctions in action[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2009, 364(1524): 1725-1731. DOI:10.1098/rstb.2008.0278 |
| [28] |
DESANTANA C N, ROZENFELD A F, MARQUET P A, et al. Topological properties of polar food webs[J]. Marine Ecology Progress Series, 2013, 474: 15-26. DOI:10.3354/meps10073 |
| [29] |
OPITZ S. Trophic interactions in Caribbean coral reefs[J]. Manila: International Center for Living Aquatic Resources Management, 1996, 341. |
| [30] |
YODZIS P. Local tropho dynamics and the interaction of marine mammals and fisheries in the Benguela ecosystem[J]. Journal of Animal Ecology, 1998, 67(4): 635-658. DOI:10.1046/j.1365-2656.1998.00224.x |
| [31] |
POLIS G A. Complex trophic interactions in deserts: an empirical critique of food-web theory[J]. The American Naturalist, 1991, 138(1): 123-155. DOI:10.1086/285208 |
| [32] |
CLOSS G P, LAKE P S. Spatial and temporal variation in the structure of an intermittent-stream food web[J]. Ecological Monographs, 1994, 64(1): 1-21. DOI:10.2307/2937053 |
| [33] |
MARTINEZ N D. Artifacts or attributes? Effects of resolution on the little rock lake food web[J]. Ecological Monographs, 1991, 61(4): 367-392. DOI:10.2307/2937047 |
| [34] |
DUNNE J A, WILLIAMS R J, MARTINEZ N D. Network structure and robustnessof marine food webs[J]. Marine Ecology Progress Series, 2004, 273: 291-302. DOI:10.3354/meps273291 |
| [35] |
HETHERINGTONE D, OLSONR J, DRAZENJ C, et al. Spatial food-web structure in the eastern tropical Pacific Ocean basedon compound-specific nitrogen isotope analysis of amino acids[J]. Limnology and Oceanography, 2017, 62(2): 541-560. DOI:10.1002/lno.10443 |
| [36] |
冯慧丽, 朱江峰, 陈彦. 基于Ecopath的热带东太平洋生态系统模型构建及其比较[J]. 上海海洋大学学报, 2019, 28(6): 921-932. FENG H L, ZHU J F, CHEN Y. Construction and historical comparison of ecosystem structure of the eastern tropical Pacific Ocean based on Ecopath model[J]. Journal of Shanghai Ocean University, 2019, 28(6): 921-932. |
| [37] |
CORTÉS E. Standardized diet compositions and trophic levels of sharks[J]. ICES Journal of Marine Science, 1999, 56(5): 707-717. DOI:10.1006/jmsc.1999.0489 |
| [38] |
张波, 唐启升. 渤、黄、东海高营养层次重要生物资源种类的营养级研究[J]. 海洋科学进展, 2004, 22(4): 393-404. ZHANG B, TANG Q S. Study on trophic level of important resources species at high troph levels in the Bohai Sea, Yellow Sea and East China Sea[J]. Advances in Marine Science, 2004, 22(4): 393-404. DOI:10.3969/j.issn.1671-6647.2004.04.001 |
| [39] |
MONTOYA J M, SOLÉR V. Topological properties of food webs: from real data to community assembly models[J]. Oikos, 2003, 102(3): 614-622. DOI:10.1034/j.1600-0706.2003.12031.x |
| [40] |
LIN Q Q, ZHU J F. Topology-based analysis of pelagic food web structure in the central and eastern tropical Pacific Ocean based on longline observer data[J]. Acta Oceanologica Sinica, 2020, 39(6): 1-9. DOI:10.1007/s13131-020-1592-2 |
2. Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai 201306, China;
3. South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510300, Guangdong, China