 2022,
Vol. 31
2022,
Vol. 31



太湖流域面积36 895 km2,太湖是流域内最大的湖泊,也是流域洪水和水资源调蓄中心。太湖湖区面积3 192 km2,其中,水面积2 338 km2,岛屿面积89 km2,湖岸山丘地面积765 km2。2008年国务院印发《太湖流域水环境综合治理总体方案》,太湖各项治理工作有序推进。近年来,太湖水生态环境发生了较大变化,主要水质指标总体呈改善趋势[1-2],原来污染严重的西北部水域水质改善效果较为显著,其正从“污水湖”向“自然湖”状态过渡[3]。
鱼类是湖泊生态系统的重要组成部分,对湖泊的稳态转换具有重要影响[4-5]。一直以来,关于太湖鱼类的调查主要集中在种类组成及群落结构,与历史调查结果相比,由于江湖阻隔、围湖造田等因素,近10年太湖鱼类物种数明显减少,其中主要为马口鱼(Opsariichthys bidens)、宽鳍

根据太湖夏季蓝藻水华及水草分布特点[13],将太湖划分为北部湖湾区、贡湖、东部沿岸区、湖心区和西南沿岸区共5个湖区。根据每个湖区的面积大小,共布设10个采样站点,分别为贡湖、梅梁湾、竺山湖、大渚、长兜港、平台山、横山、七都、胥口和漫山(图 1),基本实现太湖不同类型湖区全覆盖。

|
图 1 太湖鱼类群落结构调查站点 Fig. 1 Fish sampling sites in Taihu Lake |
2016—2020年,每年春、夏两季使用多目复合刺网和定置串联笼壶对太湖不同类型湖区10个站点的鱼类群落结构进行调查。各采样站位布设3张多目复合刺网和3条定置串联笼壶。多目复合刺网规格由1.2、2、4、6、8、10、14 cm等7种网目组成,其中1.2、2、4 cm网目刺网长度各15 m,6、8、10、14 cm网目刺网长度各20 m;多目复合刺网总长为125 m、高1.5 m。定置串联笼壶网目为1.6 cm,长、宽、高分别为10 m、40 cm、40 cm。每次于采样前一晚下午18:00左右放网,次日清晨6:00左右收网。同时,综合太湖周边水产品市场、太湖开捕期高踏网走访调查结果,并走访渔政管理部门,定性了解太湖当年出现的其他鱼类。
参考《太湖鱼类志》[14],现场鉴定采集到的鱼类样本,现场未鉴定出种类的样本,固定于4%的甲醛溶液后带回实验室再做详细鉴定。实验室内进行鱼类样本体长(精确至1 mm)、体质量(精确至0.1 g)等常规生物学测量。对于样本数大于30尾的物种,随机抽取30尾,测定其全长、体长、体质量;对于样品数少于30尾的,所有渔获物全部测定。
1.3 数据分析 1.3.1 Shannon-Wiener指数(H′)、Margalef指数(D)和Pielou指数(J)群落多样性除了受取样大小、数量的分布外,主要依赖于群落中种类数多少及个体分布是否均匀。采用Shannon-Wiener指数(H′)、Margalef指数(D)和Pielou指数(J)评价太湖鱼类生物多样性。
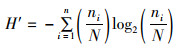 (1)
(1)
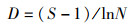 (2)
(2)
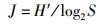 (3)
(3)
式中:S为种类数;ni为第i种的丰度;N为总个体数。
1.3.2 相对重要性指数选用相对重要性指数(IRI)描述群落优势度将IRI>1 000定义为优势种。
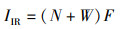 (4)
(4)
式中:IIR为相对重要性指数;N为渔获物中各物种的尾数比例;W为渔获物中各物种的质量比例;F为各物种在各断面所有抽样次数中出现的频率。
1.3.3 鱼类资源量估算体长股分析法(length based cohort analysis, LCA)是鱼类资源量评估高效易行的方法之一,该方法由JONES[15]于1974年提出,是在POPE的年龄结构时代分析模型基础上的发展,即考察资源量变动时,以体长代替年龄,从而有效地利用渔获物体长数据资料[16-17]。基于太湖渔业管理委员会办公室的鱼类资源量年度捕捞量数据及当年野外调查实测数据,采用体长股分析法估算2016—2020年太湖主要经济鱼类资源量。
收集太湖流域管理局太湖水位、排水量数据以及湖区33个站点的逐月水质及蓝藻监测数据用于表征2016年以来太湖水质及蓝藻密度变化。收集太湖渔业管理委员会办公室2016—2020年太湖鱼类人工增殖放流量及捕捞量数据。
除了鱼类物种数统计采用太湖周边水产品市场、高踏网调查结果及渔管会调查信息,其他数据统计均基于10个样点的现场调查成果。采用Pearson相关性系数分析鱼类群落结构多样性指数与太湖水环境指标之间的相关性。数据处理及统计分析采用Excel 2013和SPSS 20.0软件完成。
2 结果与分析 2.1 2016—2020太湖鱼类群落结构变化 2.1.1 2020年群落结构组成2020年春季和夏季调查累计收集渔获物5 461尾,总计质量为122 410.57 g。其中,多目复合刺网渔获物为116 434.45 g(3 760尾),定置串联笼壶渔获物为5 976.12 g(1 701尾)。共发现鱼类56种,隶属9目16科40属,其中鳗鲡(Anguilla japonica)、鲻(Mugil cephalus)、黄鳝(Monopterus albus)、鳜(Siniperca chuatsi)、双带缟虾虎鱼(Tridentiger bifasciatus)、红狼牙虾虎鱼(Odontamblyopus rubicundus)、中华花鲈(Lateolabrax japonicus)、乌鳢(Channa argus)、暗纹东方鲀(Takifugu obscurus)、细鳞斜颌鲴(Xenocypris microlepis)和鳡(Elopichthys bambusa)为太湖周边水产品市场和开捕节后的高踏网调查品种。56种鱼类中,以鲤形目鱼类居多,共34种,占总种类数的60.7%;其次为鲈形目(10种)和鲇形目(4种)鱼类,占比分别为17.6%和7.1%;鳗鲡目、鲱形目、颌针形目、鲻形目、鲀形目和合鳃目等种类仅为1~2种,占总种类数的14.6%。
2020年太湖鱼类全长为35.6~840.0 mm,平均长度为112.1 mm,其中87.9%的鱼类全长在200 mm以下;体质量为0.18~7 850.0 g,平均为25.7 g,其中88.2%的鱼类体质量在50 g以下(图 2)。2020年太湖鱼类群落结构Shannon-Wiener指数、Margalef指数和Pielou指数分别为1.29、2.55和0.45,总体处于较低水平。

|
图 2 2020年太湖鱼类全长和体质量分布直方图 Fig. 2 Histogram of fish total length and body mass of Taihu Lake in 2020 |
2016—2020年,在太湖分别发现鱼类47、56、57、58和56种,鱼类物种数总体稳定,近5年一共调查监测发现鱼类68种(表 1)。对比发现,2016—2020年太湖鱼类种群结构发生了较大变化: 2016年太湖主要优势种为鲤(Cyprinus carpio)、鲢(Hypophthalmichthys molitrix)、鲫(Carassius auratus),相对重要性指数分别为2 832.93、2 430.63和2 237.89;2017年主要优势种转变为刀鲚和鲢,相对重要性指数分别为11 294.56和2 104.24;而2020年太湖主要优势种又发生了变化,转变为刀鲚、鲫、鲤,相对重要性指数分别为9 219.28、1 698.78和1 075.27。同时也可以看出,鲫、鲤等底栖杂食性鱼类是太湖的优势类群。

|
表 1 2016—2020年太湖鱼类种类组成 Tab.1 Fish species of Taihu Lake from 2016 to 2020 |
LCA模型统计结果表明,2016—2019年刀鲚资源量分别为27 660 t、28 580 t、30 100 t和31 822 t,占比分别为35.4%、34.6%、38.2%和38.9%,均处于优势地位,反映出刀鲚对太湖生态环境的高度适应能力。同时,进一步分析发现,2016—2020年太湖洄游性鱼类种类数占比分别为12.8%、12.3%、10.3%、11.9%和16.4%,2016年和2020年的占比明显高于其他年份。
统计结果表明,2016—2020年太湖鱼类群落结构多样性波动较大且总体处于较低水平。Shannon-Wiener指数波动范围为1.12~2.08,Margalef指数波动范围为1.90~3.55,Pielou指数波动范围为0.35~0.52, 见图 3。

|
图 3 2016—2020年太湖鱼类多样性变化 Fig. 3 Fish community diversity indexes of Taihu Lake from 2016 to 2020 |
2016—2020年太湖主要水质指标(表 2)总体保持稳定:高锰酸盐质量浓度在4.24~4.60 mg/L波动,按照地表水环境质量标准(GB 3838—2002)均为Ⅲ类;氨氮均低于0.11 mg/L,为Ⅰ类;总磷质量浓度在0.073~0.087 mg/L波动,为Ⅳ类;总氮质量浓度在1.45~1.96 mg/L波动,为Ⅳ~Ⅴ类;蓝藻密度在8 624×104~11 766×104 L-1之间波动。根据《水华遥感与地面监测评价技术规范》(HJ1098—2020)确定的藻类密度水华程度分级标准,当藻类密度≥5.0×107 L-1时,则认为水体发生中度水华;当藻类密度≥1.0×108 L-1时,则认为水体发生重度水华。可以看出,2016年以来太湖蓝藻水华程度较为严重,在中度到重度水华之间波动。

|
表 2 2016—2020年太湖主要水质指标平均值 Tab.2 Main water environmental factors content in Taihu Lake from 2016 to 2020 |
分析表明,Shannon-Wiener指数、Margalef指数和Pielou指数与太湖高锰酸盐、氨氮、总磷和总氮质量浓度等主要水质指标之间具有一定相关性但相关性均不显著,与蓝藻密度之间负相关但相关性也均不显著,见表 3。2016—2020年太湖年平均水位分别为3.58、3.27、3.29、3.31和3.39 m,水位波动较大且总体处于较高水平。2016年,太湖流域发生特大洪水,年内太湖水位最高涨至4.87 m,超过保证水位(4.65 m)0.22 m,仅低于1999年历史最高水位(4.97 m)0.10 m,为1954年以来第二高水位。2020年,太湖流域发生超标准洪水,年内太湖水位最高涨至4.79 m,与1991年并列历史第3位。相关性分析结果表明,太湖年平均水位与Shannon-Wiener指数、Margalef指数和Pielou指数均正相关,但仅与Pielou指数的相关性达到显著水平(r=0.97,P < 0.01)。

|
表 3 鱼类多样性指数与太湖主要环境因子相关性 Tab.3 Relationship between fish diversity index and main water environment indicators |
2016—2020年共在太湖调查到鱼类68种,每年调查得到的种类数总体稳定。2009—2010年的调查结果显示太湖的鱼类优势种为鲚、间下
连续5年的监测结果表明,太湖鱼类优势种单一化的问题较为突出,且低龄化、低值化现象日益明显,其中仅刀鲚一种鱼类占到85%,重要鱼类生境破坏、湖体富营养化加剧以及不合理的渔业生产方式是导致该现象的主要原因[1, 6]。已有研究[18]表明,翘嘴鲌、蒙古鲌等肉食性鱼类在太湖鱼类的营养生态位中处于较高水平,能够起到抑制刀鲚和小型鱼类种群规模的目的。根据历史资料,20世纪80年代,太湖大型鲌类的资源占比可接近全湖总资源量的13%,截至2019年,该项数据已下降为不足2%。太湖中顶级消费者数量减少,有利于刀鲚等小型次级消费者数量的增加,再加上这些小型鱼类繁殖力强、食性杂、摄食强度高、生长速度快、世代更迭周期短[19-21],会大量侵占其他鱼类的生存空间,从而不利于太湖生态系统的稳定性。
3.2 流域大洪水对太湖鱼类群落结构影响2016年以来不同季度的调查结果表明,春季调查得到的种类数较为稳定,夏季变化较大,且2020年夏季在太湖调查发现暗纹东方鲀为近5年首次。夏季为太湖流域传统汛期,流域降雨多且集中,为降低太湖水位增加调蓄空间以保证流域防洪安全,每年夏季太湖流域管理局会根据流域水雨情及时调整太湖排水调度方案。2016和2020年太湖流域均发生大洪水,这两年太湖的排水量显著高于其他年份,而其中相当一部分水量会通过望虞河排出,进入长江。望虞河全长60.3 km,是直接连接长江与太湖的唯一通道[9]。统计结果显示,2016年和2020年望虞河长江枢纽排水量分别为23.3亿m3和23.7亿m3,是降雨平水年2017年的3倍,排水量的上升显著增加了太湖与长江的江湖连通性,这可能是2016年和2020年太湖的洄游鱼类种类占比高于近5年中其他年份的一个主要原因。与20世纪80年代相比,太湖洄游性鱼类种类明显减少,洄游通道阻隔可能是主要原因之一[6],本次研究结果也印证了这一结论。
3.3 太湖鱼类群落结构对水环境的响应许多研究[21-23]结果均表明,水体富营养化会导致鱼类群落结构退化。本研究发现2016年以来鱼类群落多样性与太湖水环境因子之间的关系并不显著。同时,谷孝鸿等[3]的研究结果表明虽然2016年以来太湖蓝藻水华强度达到历史最高水平,但太湖鱼类捕捞产量也达到1956年以来最大。分析导致太湖鱼类与水环境因子相关性不显著的原因可能有以下两方面。
一是出入太湖的氮磷营养盐收支不均,尤其是2016—2018连续3年出现“收”大于“支”的现象,导致湖体营养盐质量浓度持续增大[24],随之而来的是饵料生物资源不断增加与湖泊渔产潜力的逐年提升,这也使得部分小型鱼类资源可凭借自身生物学特征而获得优先增长的机遇,有研究[23]也证实了太湖近30年来的营养盐质量浓度与刀鲚等小型资源呈正比关系;二是太湖鱼类未达到其生态容量,以滤食性鱼类鲢鳙为例,当前鲢鳙资源量仅为10.21 t/km2,而其生态容量可达到51.6 t/km2,有较大的资源增殖空间,如将刀鲚资源量限制在9.49 t/km2, 鲢鳙生态容量可达206.4 t/km2,且能促使太湖生态系统进一步成熟稳定[25]。综上所述,合理优化太湖鱼类群落结构,限制外源性营养物质输入是基础保障,此外,还需因地制宜根据太湖鱼类资源现状与生态容量,完善增殖放流技术体系,确定放流的种类、规格与数量,定向从湖体中移出目标群体,强化人工调控力度。
3.4 “十年禁渔”背景下对太湖鱼类保护对策建议《中华人民共和国长江保护法》明确规定,在国家规定的期限内,长江干流和重要支流、大型通江湖泊、长江河口规定区域等重点水域全面禁止天然渔业资源的生产性捕捞。太湖全部水域已于2020年10月1日起全部停止捕捞作业,其中3个保护区2.67万hm2水域将常年禁捕,其余18.67万hm2水域将全面退捕。鱼类处在湖泊生态系统中食物链的顶端,其群落结构组成对湖泊稳态起着重要作用[4, 26]。目前,太湖大部分水域处于蓝藻水华主导的藻型湖区[27]。许多研究[28-31]结果表明,实施以鲢鳙增殖放流为主的非经典生物操纵在抑制浅水湖泊蓝藻水华暴发强度、降低水体叶绿素a质量浓度、改善水环境方面取得了积极进展和明显成效,但也有部分研究结果表明鲢鳙鱼控藻成效不明显[32],且鲢鳙鱼粪便中的藻类仍具有光合活性[29],迫切需要密切结合太湖的营养水平、水华强度等要素进一步加强以渔控藻的太湖鲢鳙鱼密度、比例和关键技术研究,以最大程度发挥太湖鲢鳙鱼控藻潜力。同时,由于增殖放流和高强度鱼类捕捞是影响太湖鱼类群落的关键因子,在太湖实施渔业禁捕、退捕的大背景下,迫切需要建立太湖鱼类精细化捕捞方案,合理控制捕捞强度[33],引导太湖鱼类群落结构的恢复,改善太湖水环境。
| [1] |
毛志刚, 谷孝鸿, 曾庆飞, 等. 太湖渔业资源现状(2009—2010年)及与水体富营养化关系浅析[J]. 湖泊科学, 2011, 23(6): 967-973. MAO Z G, GU X H, ZENG Q F, et al. Status and changes of fishery resources(2009-2010) in Lake Taihu and their responses to water eutrophication[J]. Journal of Lake Science, 2011, 23(6): 967-973. |
| [2] |
朱广伟, 秦伯强, 张运林, 等. 2005—2017年北部太湖水体叶绿素a和营养盐变化及影响因素[J]. 湖泊科学, 2018, 30(2): 279-295. ZHU G W, QIN B Q, ZHANG Y L, et al. Variation and driving factors of nutrients and chlorophyll-a concentrations in northern region of Lake Taihu, China, 2005-2017[J]. Journal of Lake Science, 2018, 30(2): 279-295. |
| [3] |
谷孝鸿, 曾庆飞, 毛志刚, 等. 太湖2007—2016十年水环境演变及"以渔改水"策略探讨[J]. 湖泊科学, 2019, 31(2): 305-318. GU X H, ZENG Q F, MAO Z G, et al. Water environment change over the period 2007-2016 and the strategy of fishery improve the water quality of Lake Taihu[J]. Journal of Lake Science, 2019, 31(2): 305-318. |
| [4] |
ANTTILA S, KETOLA M, KUOPPAMÄKI K, et al. Identification of a biomanipulation-driven regime shift in Lake Vesijärvi: implications for lake management[J]. Freshwater Biology, 2013, 58(7): 1494-1502. DOI:10.1111/fwb.12150 |
| [5] |
AUBER A, TRAVERS-Trolet M, VILLANUEVA M C, et al. Regime shift in an exploited fish community related to natural climate oscillations[J]. Plose One, 2015, 10(7): e129883. |
| [6] |
毛志刚, 谷孝鸿, 曾庆飞, 等. 太湖鱼类群落结构及多样性[J]. 生态学杂志, 2011, 30(12): 2836-2842. MAO Z G, GU X H, ZENG Q F, et al. Community structure and diversity of fish in Lake Taihu[J]. Chinese Journal of Ecology, 2011, 30(12): 2836-2842. DOI:10.13292/j.1000-4890.2011.0435 |
| [7] |
徐东坡, 凡迎春, 周彦锋, 等. 太湖鳑鲏亚科鱼类群落结构及其时空变动[J]. 上海海洋大学学报, 2018, 27(1): 115-125. XU D P, FAN Y C, ZHOU Y F, et al. Spatial and temporal variations of Acheilognathinae in Lake Taihu[J]. Journal of Shanghai Ocean University, 2018, 27(1): 115-125. |
| [8] |
徐东坡, 杨彦平, 周彦锋, 等. 太湖水域 亚科鱼类的时空分布[J]. 长江流域资源与环境, 2018, 27(2): 371-379.XU D P, YAND Y P, ZHOU Y F, et al. Spatial-temporal feature of Gobioninae fishes in Lake Taihu[J]. Resources and Environment in the Yangtze Basin, 2018, 27(2): 371-379. |
| [9] |
熊满辉, 任泷, 徐东坡. 太湖及"引江济太"河道鱼类群落结构调查研究[J]. 水生态学杂志, 2020, 1-10. XIONG M H, REN L, XU D P. Community structure and diversity of fish in Taihu Lake and Wangyuehe River[J]. Journal of Hydroecology, 2020, 1-10. |
| [10] |
QIN B, PAERL H W, BROOKES J D, et al. Why Lake Taihu continues to be plagued with cyanobacterial blooms through 10 years (2007-2017) efforts[J]. Science Bulletin, 2019, 64(6): 354-356. DOI:10.1016/j.scib.2019.02.008 |
| [11] |
朱伟, 胡思远, 冯甘雨, 等. 特大洪水对浅水湖泊磷的影响: 以2016年太湖为例[J]. 湖泊科学, 2020, 32(2): 325-336. ZHU W, HU S Y, FENG G Y, et al. Effects of great floods on phosphorus in shallow lakes: a case study of Lake Taihu in 2016[J]. Journal of Lake Science, 2020, 32(2): 325-336. |
| [12] |
何俊, 谷孝鸿, 王小林, 等. 太湖鱼类放流增殖的有效数量和合理结构[J]. 湖泊科学, 2012, 24(1): 104-110. HE J, GU X H, WANG X L, et al. Fish stocking quantities and structures of the fishery resources enhancement in Lake Taihu[J]. Journal of Lake Science, 2012, 24(1): 104-110. |
| [13] |
赵凯, 周彦锋, 蒋兆林, 等. 1960年以来太湖水生植被演变[J]. 湖泊科学, 2017, 29(29): 351-362. ZHAO K, ZHOU Y F, JIANG Z L, et al. Changes of aquatic vegetation in Lake Taihu since 1960[J]. Journal of Lake Science, 2017, 29(2): 351-362. |
| [14] |
倪勇, 朱成德. 太湖鱼类志[M]. 上海科学技术出版社, 2005. NI Y, ZHU C D. Fish dictionary of the Taihu Lake[M]. Shanghai: Science Publishing Company, 2005. |
| [15] |
JONE S. Assessing the long-term effects of changes in fishing effort and mesh size from length composition data[J]. ICESCN, 1974(33): 1-13. |
| [16] |
吴斌, 方春林, 贺刚, 等. FiSATⅡ软件支持下的体长股分析法探讨[J]. 南方水产科学, 2013, 9(4): 94-98. WU B, FANG C L, HE G, et al. FiSAT Ⅱ software supported length based cohort analysis[J]. South China Fisheries Sciences, 2013, 9(4): 94-98. |
| [17] |
虞功亮, 刘军, 许蕴玕, 等. 葛洲坝下游江段中华鲟产卵场食卵鱼类资源量估算[J]. 水生生物学报, 2002, 26(6): 591-599. YU G L, LIU J, XU Y X, et al. Estimation on abundance of egg-predatory fishes in the spawning ground of Chinese sturgeon below the Gezhouba Dam[J]. Acta Hydrobiologica Sinica, 2002, 26(6): 591-599. |
| [18] |
刘恩生, 吴林坤, 曹萍, 等. 太湖鲌鱼数量变化规律及生态效应分析[J]. 水利渔业, 2007(3): 70-73. LIU E S, WU L K, CAO P, et al. Population dynamics and ecological responses of fishes in Culter in Taihu Lake[J]. Reservoir Fisheries, 2007(3): 70-73. |
| [19] |
于谨磊, 何虎, 李宽意, 等. 太湖贡湖湾鲚(Coilia ectenes taihuensis Yen et Lin)食物组成的季节变化[J]. 湖泊科学, 2012, 24(5): 765-770. YU J L, HE H, LI K Y, et al. Seasonal variations in the diets of Coilia ectenes taihuensis Yen et Lin in Gonghu Bay of Lake Taihu[J]. Journal of Lake Science, 2012, 24(5): 765-770. |
| [20] |
王银平, 谷孝鸿, 曾庆飞, 等. 太湖不同生态型湖区湖鲚(Coilia ectenes taihuensis)食物组成及其季节变化[J]. 湖泊科学, 2016, 28(5): 1078-1085. WANG Y P, GU X H, ZENG Q F, et al. Seasonal variations in the diet composition of Coilia ectenes taihuensis in different ecotypes of Lake Taihu[J]. Journal of Lake Science, 2016, 28(5): 1078-1085. |
| [21] |
刘恩生, 刘正文, 陈伟民, 等. 太湖湖鲚渔获量变化与生物环境间相互关系[J]. 湖泊科学, 2005, 17(4): 340-345. LIU E S, LIU Z G, CHEN W M, et al. A study on the chaise of Lake Anchovy(Coilia ectenes taihuensis Yen et Lin) catches and its mutual relationship to the biological environment in Lake Taihu[J]. Journal of Lake Science, 2005, 17(4): 340-345. |
| [22] |
刘恩生, 刘正文, 鲍传和. 太湖鲫鱼数量变化的规律及与环境间关系的分析[J]. 湖泊科学, 2007, 19(3): 345-350. LIU E S, LIU Z W, BAO C H. The changes of catches of Carassius auratus and relationship with environment in Lake Taihu, China[J]. Journal of Lake Science, 2007, 19(3): 345-350. |
| [23] |
刘恩生, 刘正文, 陈伟民, 等. 太湖鱼类产量、组成的变动规律及与环境的关系[J]. 湖泊科学, 2005, 17(3): 251-255. LIU E S, LIU Z W, CHEN W M, et al. Changes in the yield and composition of the fish catches and their relation to the environmental factors in Lake Taihu[J]. Journal of Lake Science, 2005, 17(3): 251-255. |
| [24] |
朱广伟, 邹伟, 国超旋, 等. 太湖水体磷浓度与赋存量长期变化(2005—2018年)及其对未来磷控制目标管理的启示[J]. 湖泊科学, 2020, 32(1): 21-35. ZHU G W, ZOU W, GUO C X, et al. Long-term variations of phosphorus concentration and capacity in Lake Taihu, 2005-2018: Implications for future phosphorus reduction target management[J]. Journal of Lake Science, 2020, 32(1): 21-35. |
| [25] |
朱广伟, 许海, 朱梦圆, 等. 三十年来长江中下游湖泊富营养化状况变迁及其影响因素[J]. 湖泊科学, 2019, 31(6): 1510-1524. ZHU G W, XU H, ZHU M Y, et al. Changing characteristics and driving factors of trophic state in lakes at the middle and lower reaches of Yangtze River in the past 30 years[J]. Lake Sci, 2019, 31(6): 1510-1524. |
| [26] |
赵旭昊, 徐东坡, 任泷, 等. 基于Ecopath模型的太湖鲢鳙生态容量评估[J]. 中国水产科学, 2021, 28(6): 785-795. ZHAO X H, XU D P, REN L, et al. Assessment of the ecological carrying capacity of silver and bighead carp in the Taihu Lake based on Ecopath model[J]. Journal of Fishery Sciences of China, 2021, 28(6): 785-795. |
| [27] |
王华, 陈华鑫, 徐兆安, 等. 2010—2017年太湖总磷浓度变化趋势分析及成因探讨[J]. 湖泊科学, 2019, 31(4): 919-929. WANG H, CHEN H X, XU Z A, et al. Variation trend of total phosphorus and its controlling factors in Lake Taihu, 2010-2017[J]. Journal of Lake Science, 2019, 31(4): 919-929. |
| [28] |
刘建康, 谢平. 揭开武汉东湖蓝藻水华消失之谜[J]. 长江流域资源与环境, 1999(3): 85-92. LIU J K, XIE P. Unraveling the enigma of the disappearance of water bloom from the East Lake (Lake Donghu) of Wuhan[J]. Resour Environ Yangtze Basin, 1999(3): 85-92. |
| [29] |
周小玉, 张根芳, 刘其根, 等. 鲢、鳙对三角帆蚌池塘藻类影响的围隔实验[J]. 水产学报, 2011, 35(5): 729-737. ZHOU X Y, ZHANG G F, LIU Q G, et al. Effects of Hyriopsis cumingii and Aristichthys nobilis on the enclosures phytoplankton community of Hypophthalmichthys molitrix pond[J]. Journal of Fisheries of China, 2011, 35(5): 729-737. |
| [30] |
GUO L, WANG Q, XIE P, et al. A non-classical biomanipulation experiment in Gonghu Bay of Lake Taihu: control of Microcystis blooms using silver and bighead carp[J]. Aquaculture Research, 2015, 9(46): 2211-2224. |
| [31] |
唐汇娟, 谢平. 围隔中不同密度鲢对浮游植物的影响[J]. 华中农业大学学报, 2006(3): 277-280. TANG H J, XIE P. Enclosure experiment on the effects of silver carp (Hypophthalmichthys molitrix) density on the phytoplankton[J]. Journal of Huazhong Agricultural University, 2006(3): 277-280. |
| [32] |
王银平, 谷孝鸿, 曾庆飞, 等. 控(微囊)藻鲢、鳙排泄物光能与生长活性[J]. 生态学报, 2014, 34(7): 1707-1715. WANG Y P, GU X H, ZENG Q F, et al. Growth and photosynthetic activity of Microcystis colonies after gut passage through silver carp and bighead carp[J]. Acta Ecologica Sinica, 2014, 34(7): 1707-1715. |
| [33] |
梅雪英, VLADIMIR R, LARS G R, 等. 杂食性鱼类对浅水水体底栖-浮游生境耦合作用的影响: 微综述[J]. 湖泊科学, 2021, 33(3): 667-674. MEI X Y, VLADIMIR R, LARS G R, et al. Effects of omnivorous fish on benthic-pelagic habitats coupling in shallow aquatic ecosystems: A minireview[J]. Journal of Lake Science, 2021, 33(3): 667-674. |