 2022,
Vol. 31
2022,
Vol. 31

2. 上海海洋大学 水产动物遗传育种上海市协同创新中心, 上海 201306;
3. 上海海洋大学 水产科学国家级实验教学示范中心, 上海 201306;
4. 北京大北农科技集团股份有限公司, 北京 100190
中华绒螯蟹(Eriocheir sinensis)是我国最重要经济蟹类之一,2019年中华绒螯蟹养殖总产量高达78万t[1]。在中华绒螯蟹的池塘养殖过程中,合理投放方形环棱螺(Bellamy aquadrata) 会显著提高中华绒螯蟹的产量[2]。目前生产上常常采用不同规格混杂的方式进行投放,但过大规格的方形环棱螺难以被中华绒螯蟹捕食[3],且会与中华绒螯蟹争夺饲料等资源[4],严重制约了中华绒螯蟹养殖经济效益的进一步提高。主要原因之一是不同规格的方形环棱螺对中华绒螯蟹捕食行为的影响尚不明确,难以形成一个投放方形环棱螺的合理标准。因此,了解中华绒螯蟹对于方形环棱螺的捕食过程至关重要。
研究[5-7]表明,蟹对于螺的捕食会受到螺规格和蟹规格的影响。如三叶真蟹(Carcinus maenas) 会优先选择规格较小的玉黍螺(Littorina irrorata) [8],蓝蟹(Callinectes sapidus) 对14~18 mm的玉黍螺捕食率显著高于更大规格的螺[9],然而,在螺的规格没有差异时,蓝蟹对螺的捕食率取决于蟹的规格[10],因为蟹的螯足必须能够损伤或粉碎螺壳才具有捕食螺的能力,而较大的蟹具有较大的螯强度[11]。
研究[12-13]发现,螺的规格和其反捕食行为会导致蟹的捕食行为发生改变,蟹对不同规格螺的行为时间分布和处理方式不同。如蓝蟹对玉黍螺的处理时间比随着螺壳宽的增大而增加[9],三叶真蟹通过粉碎处理方式捕食规格较小的玉黍螺而用孔探处理方式捕食规格更大的玉黍螺[8]。此外,螺会采取一系列的反捕食行为逃脱蟹的捕食。螺通过攀爬行为或打洞行为以逃避捕食者。如玉黍螺可以通过攀爬茎秆来躲避蓝蟹的捕食[14],加州角螺(Cerithideopsis californica) 可以通过增加打洞行为的比例来躲避厚纹蟹(Pachygrapsus crassipes) 的捕食[15]。
迄今为止,关于方形环棱螺规格对中华绒螯蟹捕食行为的影响还未见报道。本研究通过视频监控系统分析了大、小规格中华绒螯蟹对3种规格方形环棱螺的捕食过程,进而比较中华绒螯蟹对方形环棱螺的选择指数、捕食率、中华绒螯蟹处理方式和方形环棱螺逃逸情况,以期为后续中华绒螯蟹捕食行为的研究提供参考,并为中华绒螯蟹养殖过程中方形环棱螺的合理投放提供理论依据。
1 材料与方法 1.1 实验用蟹来源及暂养实验所用的中华绒螯蟹均取自上海海洋大学崇明养殖基地,捕获后置于循环水系统中暂养2周。暂养期间水温(25.0±0.5)℃、光照周期为12 h∶12 h (L∶D)。同时每天投喂1次方形环棱螺,以适应捕食环境。
实验所用的方形环棱螺取自于上海海洋大学滨海基地,采集后置于水温(25.0±1.1) ℃、光照周期为12 h∶ 12 h (L∶D) 的水族箱中暂养,每天换水1次并连续曝气。
1.2 实验设计暂养结束后,用游标卡尺(M150,上海精美量具厂,准确度= 0. 02 mm) 测量中华绒螯蟹的甲壳宽,筛选蟹体健壮、螯足及附肢完整活力足的中华绒螯蟹雄蟹供实验所用。选取甲壳宽分别为(38.0±0.5) mm和(43.0±0.5) mm的2种规格的中华绒螯蟹,分别记做小规格(small crab,SC) 和大规格(large crab,LC),选取螺壳宽分别为(5.5±0.5) mm、(7.5±0.5) mm和(9.5±0.5) mm的方形环棱螺,分别记做小规格(small snail,SS)、中规格(moderate snail,MS) 以及大规格(large snail,LS)。
1.2.1 规格选择实验规格选择实验旨为确定中华绒螯蟹对不同规格方形环棱螺的偏好。将中华绒螯蟹转移至具有循环水系统的圆形水族罐(50 cm×50 cm×70 cm,78.5 L) 中进行实验。水族罐每天持续循环曝气,水温为(25.0±0.9)℃,光照条件12 h∶12 h (L∶D)。每个圆形水族罐都提前4天放入1只中华绒螯蟹,以保证中华绒螯蟹有足够的时间适应水族罐环境,在此期间不做任何喂养以统一中华绒螯蟹的饥饿水平。实验开始后向水族罐投喂39只方形环棱螺(小、中、大规格的螺各13只),24 h后收集并测量未食用的方形环棱螺。每组实验设置4个重复。
1.2.2 捕食行为实验捕食行为实验旨为确定不同规格方形环棱螺对于中华绒螯蟹捕食行为的影响。实验共分为6个处理组,分别为小规格中华绒螯蟹-小规格方形环棱螺(SC-SS)、小规格中华绒螯蟹-中规格方形环棱螺(SC-MS)、小规格中华绒螯蟹-大规格方形环棱螺(SC-LS)、大规格中华绒螯蟹-小规格方形环棱螺(LC-SS)、大规格中华绒螯蟹-中规格方形环棱螺(LC-MS)、大规格中华绒螯蟹-大规格方形环棱螺(LC-LS),每组实验设置4个重复。
捕食行为实验装置和养殖管理与规格选择实验一致。实验开始后向水族罐投喂40只方形环棱螺。实验在96 h后结束,且每24 h收集并测量未食用的方形环棱螺,并加入鲜活的方形环棱螺以保持猎物密度。同时,为了观察中华绒螯蟹的捕食行为,每1次实验都通过水族罐上方的摄像机(Hikvision, DS-2CD3T86FD-L, China) 完整地记录下来,并设一相同条件下的水族罐放养相同密度的方形环棱螺,用来统计方形环棱螺24 h的自然死亡率,统计结果表明在没有中华绒螯蟹捕食的情况下,方形环棱螺自然死亡率为0。
1.3 测定指标及方法 1.3.1 捕食率和选择指数实验结束后统计水族罐内被捕食的方形环棱螺数量,捕食率(predation rate, PR) 为每只中华绒螯蟹每24 h捕食的方形环棱螺的数量。计算Chesson选择指数α[16]:
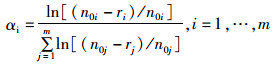 (1)
(1)
式中:i和j表示方形环棱螺的不同规格;r表示方形环棱螺的数量;n0表示实验开始时环境中的方形环棱螺数量;m表示方形环棱螺规格等级总数。
1.3.2 捕食行为指标本实验随机选取白天和晚上各1 h来观察和量化中华绒螯蟹的捕食行为[17]。统计相遇(在搜寻过程中蟹近距离接触螺)、捕获(蟹用螯足操纵螺)以及摄食(蟹通过螯足和腭足摄食螺肉)的频数来量化相遇概率(encounter rate,ER)、相遇后捕获概率(probability of capture upon encounter,PCE)、捕获后摄食概率(probability of consume upon capture,PCC)等指标;统计蟹所处的静止状态、移动状态、搜寻状态以及处理状态的持续时间来量化静止时间比(proportion of stationary time,RST)、移动时间比(proportion of moving time,RMT)、搜寻时间比(proportion of searching time,RSRT)、处理时间比(proportion of handling time,RHT)、平均处理时间(handling time per snail,TH)等指标。计算公式如下:
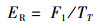 (2)
(2)
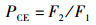 (3)
(3)
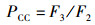 (4)
(4)
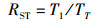 (5)
(5)
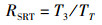 (6)
(6)
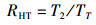 (7)
(7)
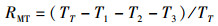 (8)
(8)
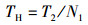 (9)
(9)
式中:ER为相遇概率;PCE为相遇后摄食概率;PCC为捕获后摄食概率;RST为静止时间比;RSRT为搜寻时间比;RHT为处理时间比;RMT为移动时间比;TH为平均处理时间。F1表示在搜寻过程中中华绒螯蟹与方形环棱螺的相遇次数; F2表示在观察时间内中华绒螯蟹对方形环棱螺的捕获次数; F3表示在观察时间内中华绒螯蟹对方形环棱螺的摄食次数;TT表示行为视频的观察时间的总时长; T1表示中华绒螯蟹静止状态的持续时间; T2表示中华绒螯蟹开始搜寻方形环棱螺到与其相遇的时间; T3表示中华绒螯蟹捕获方形环棱螺后对其处理的时间;N1表示在观察时间内方形环棱螺螺被捕食的总数量。
1.3.3 蟹处理方式与螺逃逸比例统计中华绒螯蟹对方形环棱螺的处理方式以及方形环棱螺的逃逸比例。其中,处理方式是根据中华绒螯蟹捕食后残留螺壳的形态(图 1)推断出来的:(1) 螺壳破碎(即只剩下螺壳碎片);(2) 螺壳边缘有一些组织缺失;(3) 不会对螺壳造成损害[18]。粉碎处理方式比(proportion of crushing tactics,PCRT)为螺壳破碎的数量与螺总数量的比值;壳削处理方式比(proportion of chipping tactics,PCHT)为螺壳损伤的数量与螺总数量的比值;孔探处理方式比(proportion of probing tactics,PPT)为螺壳未损伤的数量与螺总数量的比值;螺逃逸比例(proportion of successful avoidance,PSA)为螺“爬出”即在水面以上或水面上的数量与螺总数量的比值[19]。

|
图 1 中华绒螯蟹对方形环棱螺的不同开壳方式 Fig. 1 Illustration of shell conditions used to infer the handling behavior of Eriocheir sinensis |
观察中华绒螯蟹对方形环棱螺的捕食过程获得的选择指数、捕食率、相遇概率、相遇后捕获概率、捕获后摄食概率、静止时间比、移动时间比、搜寻时间比、处理时间比、粉碎处理方式比、平均处理时间、螺逃逸比例等指标均用平均值±标准差(Mean±SD) 表示,并使用IBM SPSS.22软件进行统计分析。采用单因素方差分析(One-Way ANOVA) 对相同规格中华绒螯蟹捕食不同规格的方形环棱螺的实验结果进行分析,然后使用Duncan氏检验进行多重比较,而大、小规格中华绒螯蟹捕食同一规格方形环棱螺时,采用独立样本t检验分析中华绒螯蟹的规格对实验结果的影响。显著性水平均设为P<0.05。在GraphPad Prism.8.3.0.538软件上绘制相关的柱状图。
2 结果 2.1 选择指数由图 2可知,中华绒螯蟹对3种规格的方形环棱螺的选择指数依次为小规格(α=0.68±0.21)、中规格(α=0.31±0.22)、大规格(α=0.04±0.01),且任意两组之间差异显著(P<0.05)。

|
图中小写字母的不同表示处理组之间差异显著(P<0.05)。 Difference of lowercase letters in the figure indicates that there are significant differences between the treatment groups (P < 0.05). 图 2 中华绒螯蟹对3种规格方形环棱螺的选择指数 Fig. 2 Selection index of Eriocheir sinensis to three size classes of Bellamy aquadrata |
由图 3可知,方形环棱螺的规格显著影响中华绒螯蟹的捕食率。SC-SS处理组的捕食率显著高于SC-MS和SC-LS处理组,且SC-MS处理组显著高于SC-LS处理组(P<0.05)。LC-SS处理组的捕食率显著高于LC-MS和LC-LS处理组,且LC-MS处理组显著高于LC-LS处理组(P<0.05)。仅在捕食大规格方形环棱螺时,中华绒螯蟹的规格影响了中华绒螯蟹的捕食率。LC-LS处理组的捕食率显著高于SC-LS处理组(P<0.05)。

|
图中小写字母的不同表示同一种规格的蟹处理不同规格螺之间差异显著(P<0.05),而不同规格蟹处理同一种规格螺的显著差异用“*”表示(P<0.05)。 Different lowercase letters in the figure indicate that there are significant differences between different sizes of snails treated by crabs of the same size (P < 0.05), while the significant differences between same sizes of snails treated by crabs of the different sizes are indicated by "*" (P < 0.05). 图 3 中华绒螯蟹的捕食率和相遇概率 Fig. 3 Predation rate and encounter rate of Eriocheir sinensis |
由图 4可知,方形环棱螺的规格也显著影响中华绒螯蟹的捕获后摄食概率: SC-LS和SC-SS处理组捕获后摄食概率显著高于SC-MS处理组(P<0.05);LC-SS处理组的相遇后捕获概率显著高于LC-MS和LC-LS处理组(P<0.05)。但大规格中华绒螯蟹捕食不同规格方形环棱螺时,其相遇概率、相遇后捕获概率以及捕获后摄食概率均有显著的影响。LC-LS和LC-MS处理组的相遇概率显著高于LC-SS处理组(P<0.05);LC-SS处理组的相遇后捕获概率显著高于LC-LS处理组(P<0.05);LC-SS处理组的相遇后捕获概率显著高于LC-MS和LC-LS处理组(P<0.05)。此外,不同规格的中华绒螯蟹也仅在捕食大规格螺时,其相遇概率、相遇后捕获概率以及捕获后摄食概率有显著的差异。LC-LS处理组的相遇概率显著高于SC-LS处理组;SC-LS处理组的相遇后捕获概率显著高于LC-LS处理组(P<0.05);SC-LS处理组的捕获后摄食概率显著高于LC-LS处理组(P<0.05)。

|
图中小写字母的不同表示同一种规格的蟹处理不同规格螺之间差异显著(P<0.05),而不同规格蟹处理同一种规格螺的显著差异用“*”表示(P<0.05)。 Different lowercase letters in the figure indicate that there are significant differences between different sizes of snails treated by crabs of the same size (P < 0.05), while the significant differences between same sizes of snails treated by crabs of the different sizes are indicated by "*"(P < 0.05). 图 4 中华绒螯蟹的相遇后捕获概率和捕获后摄食概率 Fig. 4 Probability of capture upon encounter and the probability of consumption upon capture of Eriocheir sinensis |
由图 5可知,方形环棱螺的规格显著影响中华绒螯蟹的移动时间比: SC-MS和SC-SS处理组的移动时间比显著高于SC-LS处理组(P<0.05);LC-MS处理组的移动时间比显著高于LC-SS和LC-LS处理组(P<0.05),但不影响搜寻时间比(P<0.05)。而仅在大规格中华绒螯蟹捕食3种规格的方形环棱螺过程中,其静止时间比和处理时间比有着显著的差异。LC-MS处理组的静止时间比显著高于LC-SS处理组(P<0.05)。LC-LS处理组的处理时间比显著高于LC-MS和LC-SS处理组(P<0.05)。此外,中华绒螯蟹的规格对于中华绒螯蟹状态行为时间分布没有显著的影响(P>0.05)。

|
图中小写字母的不同表示同一种规格的蟹处理不同规格螺之间差异显著(P<0.05),而不同规格蟹处理同一种规格螺的显著差异用“*”表示(P<0.05)。 Different lowercase letters in the figure indicate that there are significant differences between different sizes of snails treated by crabs of the same size (P < 0.05), while the significant differences between same sizes of snails treated by crabs of the different sizes are indicated by "*"(P < 0.05). 图 5 中华绒螯蟹的状态行为时间分布 Fig. 5 Temporal distribution of state behavior of Eriocheir sinensis |
由图 6可知,方形环棱螺的规格显著影响中华绒螯蟹的粉碎处理方式比和平均处理时间: 捕食小规格螺的粉碎处理方式比和显著高于捕食中规格和大规格组(P<0.05);捕食大规格组的平均处理时间显著高于中规格组和小规格组(P<0.05)。此外,仅在捕食大规格方形环棱螺时,蟹的规格显著影响中华绒螯蟹蟹的粉碎处理方式比和平均处理时间: LC-LS处理组的粉碎处理方式比显著高于LC-SS和LC-MS处理组,且LC-MS处理组的粉碎处理方式比显著高于LC-SS处理组(P<0.05);LC-LS处理组的平均处理时间显著高于LC-MS和LC-SS处理组(P<0.05);LC-MS处理组的壳削处理方式比显著高于LC-SS处理组、LC-LS处理组以及SC-MS处理组(P<0.05);大规格螺组的孔探处理方式比显著高于中、小规格螺组(P<0.05)。

|
图中不同小写字母表示同一种规格的蟹处理不同规格螺之间差异显著(P<0.05),而不同规格蟹处理同一种规格螺的显著差异用“*”表示(P<0.05)。 Different lowercase letters in the figure indicate that there are significant differences between different sizes of snails treated by crabs of the same size (P < 0.05), while the significant differences between same sizes of snails treated by crabs of the different sizes are indicated by "*" (P < 0.05). 图 6 中华绒螯蟹处理方式和平均处理时间的差异 Fig. 6 Differences in shell-breaking techniques and handing time per snail of Eriocheir sinensis |
由图 7可知: 方形环棱螺的规格显著影响螺的逃逸比例,小规格蟹捕食大规格螺的螺逃逸比例显著高于小规格组(P<0.05);仅在捕食大规格方形环棱螺时,LC-SS组和LC-MS处理组的逃逸比例显著高于LC-LS处理组(P<0.05)。

|
图中小写字母的不同表示同一种规格的蟹处理不同规格螺之间差异显著(P<0.05),而不同规格蟹处理同一种规格螺的显著差异用“*”表示(P<0.05)。 Different lowercase letters in the figure indicate that there are significant differences between different sizes of snails treated by crabs of the same size (P < 0.05), while the significant differences between same sizes of snails treated by crabs of the different sizes are indicated by "*" (P < 0.05). 图 7 方形环棱螺的逃逸比例 Fig. 7 Proportion of successful avoidance of Bellamya quadrata |
研究[3]表明,蟹对螺捕食率主要取决于捕食者和猎物的规格。蟹对不同规格螺的捕食率从高到低呈小规格螺>中规格螺>大规格螺的趋势[15]。本研究也有类似结果,方形环棱螺的规格越大,中华绒螯蟹对其选择系数和捕食率越低。造成这种差异可能有以下两种原因。第一,螺壳形态和螺壳强度是影响蟹对螺捕食率的重要因素,方形环棱螺壳强度随着方形环棱螺规格的降低而下降,方形环棱螺壳强度越低,越易被中华绒螯蟹捕食[20-21];第二,小规格的方形环棱螺能量收益更低,中华绒螯蟹需要更多地摄食小规格方形环棱螺的来满足自身的能量需要[22]。但仅在捕食大规格方形环棱螺时,大规格蟹的捕食率大于小规格蟹,这可能是由于小规格中华绒螯蟹的甲壳宽与大规格方形环棱螺的壳宽比值过低,限制了小规格中华绒螯蟹对大规格方形环棱螺的捕食。例如:蓝蟹一般在蟹的甲宽与螺的壳宽比值大于6的情况下对玉黍螺的捕食率较高。在较低的体型比例下,蟹对螺捕食率要低得多[23]。结果证明了捕食者和猎物的规格确实会影响到中华绒螯蟹对方形环棱螺的捕食率,尽管在实验室条件下可能高估了中华绒螯蟹对方形环棱螺的捕食率。一方面,中华绒螯蟹是杂食性甲壳动物,但本次实验只有单一种类的食物来源,没有大型藻类等替代食物,且实验室环境下不需要承担任何额外的能量消耗来搜寻方形环棱螺,实验室条件可能导致其捕食率偏高;另一方面,本次实验选取是中华绒螯蟹雄蟹,而相对于同一阶段的中华绒螯蟹雌性蟹来说,雄蟹有更大的螯足,更易捕食同等规格的方形环棱螺,使用中华绒螯蟹雄蟹可能高估了中华绒螯蟹对方形环棱螺的捕食率[11]。
研究[11]表明,捕食率主要由相遇概率、相遇后捕获概率以及捕获后摄食概率共同决定。在本研究中,相遇后捕获概率、捕获后摄食概率与捕食率的趋势一致,但相遇概率与捕食率的趋势相反。这说明中华绒螯蟹对不同规格的方形环棱螺的捕食率主要受相遇后捕获概率和捕获后摄食概率影响。与SUN等[17]主要由相遇后捕获概率决定的研究结果不一致,可能是三疣梭子蟹(Portunus trituberculatus) 和中华绒螯蟹的食性、螯足力量以及栖息环境的差异导致的,也有可能受方形环棱螺和菲律宾蛤仔(Ruditapes philippinarum) 的物种特异性和规格差异影响。此外,在本研究中,方形环棱螺的规格显著影响中华绒螯蟹的捕获后摄食概率,说明方形环棱螺的规格在中华绒螯蟹评估是否摄食过程有着重要影响,因为中华绒螯蟹主要通过视觉和嗅觉定位食物位置,而靠味觉和触觉评估是否捕食该猎物[24],而本实验选取为同一物种,味觉并不是影响其差异的主要原因,故方形环棱螺的规格通过影响触觉的评估来影响其捕获后摄食概率。此外,不同规格的中华绒螯蟹的相遇概率、相遇后捕获概率以及捕获后摄食概率均与捕食率一致,仅在捕食大规格方形环棱螺时差异显著。
研究[25]表明,状态行为时间分布是衡量捕食行为重要指标之一。许多关于蟹捕食硬壳猎物的研究表明,蟹更喜欢比预期更小的猎物,为了解释这一个差异而提出时间最小化的原则似乎是一种代替能量最大化选择标准。例如,ROVERO等[26]认为猎物处理时间相比于能量消耗是一种更精确的衡量标准。本研究发现,方形环棱螺的规格显著影响中华绒螯蟹的移动时间比,但不影响其搜寻时间比。这与WÓJCIK等[27]的研究结果一致,这可能由于实验水族箱和猎物密度限制了中华绒螯蟹的搜寻行为。此外,就处理时间比而言,大规格蟹捕食大规格螺的处理时间比显著高于其他规格,主要是由于大规格方形环棱螺壳宽较宽,限制了中华绒螯蟹螯足的处理,而大规格中华绒螯蟹处理中规格方形环棱螺的处理时间比最低,这似乎暗示了中规格方形环棱螺是大规格中华绒螯蟹的较适规格,这与COTE等[9]的结果吻合,尽管蟹更喜欢比预期更小的猎物,但由于捕食效率降低和螯足形态的局限性,可能会阻止蟹选择较小的腹足类动物。
研究[19]表明,螺会爬上植物或其他突出的表面以躲避捕食者。本研究观察到:方形环棱螺躲避中华绒螯蟹捕食的方式是缩进壳里,或者爬到水面上;方形环棱螺的规格和中华绒螯蟹的规格均会影响螺的逃逸比例,方形环棱螺规格越大,螺逃逸比例越高,可能是由于方形环棱螺的规格越大,其移动速度越快,螺壳强度越高,更难被捕食;中华绒螯蟹规格越大,螺逃逸比例低,这可能不同规格的中华绒螯蟹的螯足形态的差异导致的。
3.2 方形环棱螺规格对中华绒螯蟹猎物处理方式的影响研究[23]表明,蓝蟹主要通过3种处理方式来捕食玉黍螺。第一,孔探方式,绿蟹使用单侧螯足通过螺口进入螺壳内,将螺的软组织压在壳壁的内侧,并将其拖向螺口,从而将螺的一部分或全部组织撬出;第二,壳削方式,绿蟹首先会沿着螺壳边缘切割,然后用孔探处理方式进行取食;第三,粉碎方式,绿蟹首先会沿着螺壳边缘切割之后用螯足将螺壳破碎或直接用螯足夹碎螺壳,取出全部组织[23]。本研究也有类似结果,中华绒螯蟹通过变换上述3种方式处理方形环棱螺,即在采用粉碎处理方式之前,中华绒螯蟹已经尝试过壳削和孔探处理方式。本研究发现,中华绒螯蟹对小规格和中规格螺捕食粉碎处理比例显著高于大规格螺,且大规格螺组的孔探处理方式比显著高于中、小规格螺组。这说明当捕食小规格螺,蟹的粉碎处理方式占据了主导地位,当捕食大规格螺时,它们大多通过孔探处理方式进行捕食,螺的规格显著影响蟹对猎物的处理方式。
此外,研究[28]表明,处理方式和平均处理时间显著相关。在本研究中,蟹对不同规格螺的平均处理时间与粉碎处理方式的比例趋势相反,蟹对大规格螺的平均处理时间显著高于小规格螺和中规格螺,说明随着螺规格的增加,由于螺壳的增大与螯足的限制导致其处理方式发生转变,粉碎处理方式比例显著下降,平均处理时间显著增加。
综上,方形环棱螺的规格对中华绒螯蟹规格选择性、捕食率和状态行为的时间分布有着显著的影响,中华绒螯蟹对小规格方形环棱螺有着明显的捕食偏好,而捕获后能否摄食是影响中华绒螯蟹捕食的重要环节。此外,中华绒螯蟹对方形环棱螺的处理方式随方形环棱螺的规格发生改变。
| [1] |
农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会. 中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2020: 44-54. Fishery Administration Bureau of Ministry of Agriculture and Rural Areas, National Aquatic Technology Promotion Center, China Fisheries Society. China fishery statistical yearbook[M]. Beijing: China Agriculture Press, 2020: 44-54. |
| [2] |
王海候, 沈明星, 陆长婴, 等. 螺蛳替代商品饲料比例对中华绒螯蟹产量和水质的影响[J]. 水产科学, 2015, 34(11): 690-695. WANG H H, SHEN M X, LU C Y, et al. Effect of replacement percentage of commercial feed by snails on yield and water quality in Chinese mitten handed crab culture[J]. Fisheries Science, 2015, 34(11): 690-695. |
| [3] |
WRIGHT R A, CROWDER L B, MARTIN T H. Selective predation by blue crabs on the gastropod, Bittium varium: Confirmation from opercula found in the sediments[J]. Estuaries, 1996, 19(1): 75-81. DOI:10.2307/1352653 |
| [4] |
王海候, 金梅娟, 沈明星, 等. 螺蛳对蟹塘产量、水质的影响及其适宜投喂量研究[J]. 水产科技情报, 2016, 43(1): 37-40. WANG H H, JIN M J, SHEN M X, et al. Effects of snail on crab yield and water quality and the optimal feeding rate[J]. Fisheries Science & Technology Information, 2016, 43(1): 37-40. |
| [5] |
WEST D L, WILLIAMS A H. Predation by Callinectes sapidus (Rathbun) within Spartina alterniflora (Loisel) marshes[J]. Journal of Experimental Marine Biology and Ecology, 1986, 100: 75-95. DOI:10.1016/0022-0981(86)90156-5 |
| [6] |
VAUGHN C C, FISHER F M. Vertical migration as a refuge from predation in intertidal marsh snails: A field test[J]. Journal of Experimental Marine Biology and Ecology, 1988, 123: 163-176. DOI:10.1016/0022-0981(88)90167-0 |
| [7] |
STANHOPE H S, BANTA W C, TEMKIN M H. Size-specific emergence of the marsh snail, Littorina irrorata: Effect of predation by blue crabs in a virginia salt marsh[J]. Gulf Research Reports, 1982, 7(2): 179-182. |
| [8] |
ROCHETTE R, DOYLE S P, EDGELL T C. Interaction between an invasive decapod and a native gastropod: predator foraging tactics and prey architectural defenses[J]. Marine Ecology Progress Series, 2007, 330: 179-188. DOI:10.3354/meps330179 |
| [9] |
COTE J, RAKOCINSKI C F, RANDALL T A. Feeding efficiency by juvenile blue crabs on two common species of micrograzer snails[J]. Journal of Experimental Marine Biology and Ecology, 2001, 264: 189-208. DOI:10.1016/S0022-0981(01)00315-X |
| [10] |
BOULDING E G. Crab-resistant features of shells of burrowing bivalves: decreasing vulnerability by increasing handling time[J]. Journal of Experimental Marine Biology and Ecology, 1984, 76: 201-223. DOI:10.1016/0022-0981(84)90189-8 |
| [11] |
YAMADA S B, BOULDING E G. Claw morphology, prey size selection and foraging efficiency in generalist and specialist shell-breaking crabs[J]. Journal of Experimental Marine Biology and Ecology, 1998, 220: 191-211. DOI:10.1016/S0022-0981(97)00122-6 |
| [12] |
ROSEWARNE P J, MORTIMER R J G, NEWTON R J, et al. Feeding behaviour, predatory functional responses and trophic interactions of the invasive Chinese mitten crab (Eriocheir sinensis) and signal crayfish (Pacifastacus leniusculus)[J]. Freshwater Biology, 2016, 61(4): 426-443. DOI:10.1111/fwb.12717 |
| [13] |
SEITZ R D, LIPCIUS R N, HINES A H, et al. Density-dependent predation, habitat Variation, and the persistence of marine bivalve prey[J]. Ecology, 2001, 82(9): 2435-2451. DOI:10.1890/0012-9658(2001)082[2435:DDPHVA]2.0.CO;2 |
| [14] |
WARREN J H. Climbing as an avoidance behaviour in the salt marsh periwinkle, Littorina irrorata (Say)[J]. Journal of Experimental Marine Biology and Ecology, 1985, 89: 11-28. DOI:10.1016/0022-0981(85)90079-6 |
| [15] |
LORDA J, HECHINGER R F, COOPER S D, et al. Intraguild predation by shore crabs affects mortality, behavior, growth, and densities of California horn snails[J]. Ecosphere, 2016, 7(5): e01262. |
| [16] |
WONG M C, PETERSON C H, KAY J. Prey size selection and bottom type influence multiple predator effects in a crab-bivalve system[J]. Marine Ecology Progress Series, 2010, 409: 143-156. DOI:10.3354/meps08621 |
| [17] |
SUN Y F, WANG F, SU X P, et al. Size matters for predator-prey relationships: The case of swimming crab Portunus trituberculatus preying on Manila clam Ruditapes philippinarum[J]. Aquaculture, 2017, 479: 638-644. DOI:10.1016/j.aquaculture.2017.07.008 |
| [18] |
MILLS C D, CLARK P F, MORRITT D. Flexible prey handling, preference and a novel capture technique in invasive, sub-adult Chinese mitten crabs[J]. Hydrobiologia, 2016, 773: 135-147. DOI:10.1007/s10750-016-2700-3 |
| [19] |
ALEXANDER J E, COVICH A P. Predator avoidance by the freshwater snail Physella virgata in response to the crayfish Procambarus simulans[J]. Oecologia, 1991, 87: 435-442. DOI:10.1007/BF00634603 |
| [20] |
DUDAS S E, MCGAW I J, DOWER J F. Selective crab predation on native and introduced bivalves in British Columbia[J]. Journal of Experimental Marine Biology and Ecology, 2005, 325: 8-17. DOI:10.1016/j.jembe.2005.04.015 |
| [21] |
MIRANDA N A F, MEASEY G J, PEER N, et al. Shell crushing resistance of alien and native thiarid gastropods to predatory crabs in South Africa[J]. Aquatic Invasions, 2016, 11(3): 303-311. DOI:10.3391/ai.2016.11.3.08 |
| [22] |
COLPO K D, LÓPEZ-GRECO L S. Dynamics of energy reserves and the cost of reproduction in female and male fiddler crabs[J]. Zoology, 2018, 126: 1-9. DOI:10.1016/j.zool.2017.11.001 |
| [23] |
SCHINDLER D E, JOHNSON B M, MACKAY N A, et al. Crab: snail size-structured interactions and salt marsh predation gradients[J]. Oecologia, 1994, 97: 49-61. DOI:10.1007/BF00317908 |
| [24] |
LEE P G, MEYERS S P. Chemoattraction and feeding stimulation in crustaceans[J]. Aquaculture Nutrition, 1996, 2: 157-164. DOI:10.1111/j.1365-2095.1996.tb00055.x |
| [25] |
BARDERA G, OWEN M A G, POUNTNEY D, et al. The effect of short-term feed-deprivation and moult status on feeding behaviour of the Pacific white shrimp (Litopenaeus vannamei)[J]. Aquaculture, 2019, 511: 734222. DOI:10.1016/j.aquaculture.2019.734222 |
| [26] |
ROVERO F, HUGHES R N, CHELAZZI G. When time is of the essence: choosing a currency for prey-handling costs[J]. Journal of Animal Ecology, 2000, 69: 683-689. DOI:10.1046/j.1365-2656.2000.00426.x |
| [27] |
WÓJCIK D, NORMANT M, DMOCHOWSKA B, et al. Impact of Chinese mitten crab Eriocheir sinensis on blue mussel Mytilus edulis trossulus-laboratory studies of claw strength, handling behavior, consumption rate, and size selective predation[J]. Oceanologia, 2015, 57: 263-270. DOI:10.1016/j.oceano.2015.03.003 |
| [28] |
CALDERWOOD J, O'CONNOR N E, ROBERTS D. Breaking and entering: Examining the role of stress and aerial exposure in predator-prey relationships between the common shore crab (Carcinus maenas) and cultivated blue mussels (Mytilus edulis)[J]. Aquaculture, 2016, 452: 217-223. DOI:10.1016/j.aquaculture.2015.11.009 |
 ,
SUN Yunfei1,2,3,
HAN Wenfeng1,2,3,
CHENG Fangzhou1,2,3,
CHU Yanwei4,
YANG Zhigang1,2,3,
CHENG Yongxu1,2,3
,
SUN Yunfei1,2,3,
HAN Wenfeng1,2,3,
CHENG Fangzhou1,2,3,
CHU Yanwei4,
YANG Zhigang1,2,3,
CHENG Yongxu1,2,3
2. Shanghai Collaborative Innovation Center for Aquatic Animal Genetics and Breeding, Shanghai Ocean University, Shanghai 201306, China;
3. National Demonstration Centre for Experimental Fisheries Science Education, Shanghai Ocean University, Shanghai 201306, China;
4. Beijing DBN Science and Technology Group CO., LTD, Beijing 100190, China