 2021,
Vol. 30
2021,
Vol. 30



2. 上海海洋大学 水产科学国家级实验教学示范中心, 上海 201306
滁河流域位于江淮之间,源于安徽东南部的丘陵山区,沿途流经安徽省合肥市、滁州市、马鞍山市和江苏省南京市,于江苏南京六合区大河口汇入长江,全长269.2 km,系长江下游北岸一级支流[1-2]。滁河襄河口至马汊河段位于安徽省滁州境内,地处淮河与长江之间,河段总长约121 km,河段内有汊河集船闸,整个河段由上游滁河干流(上游河段)、清流河、来安河以及下游滁河干流(下游河段)组成,各河段上一节点位置依次为滁河襄河口、清流河谢郢、来安河水口以及滁河三汊湾[3];下游滁河干流经马汊河与长江连通。因此,调查水体同时具备江湖水体特性,具有丰富的渔业资源储备[4]。
目前,关于滁河渔业资源的研究较少,现有报道多集中于滁河河道整治[5]、流域水质污染评价[6]、悬浮泥沙迁移[7]、蓄洪调度和防洪效应[8]等方面。作为滁河重要河段之一,滁河襄河口至马汊河段鱼类群落多样性是研究长江一级支流滁河生态环境变化的重要前提之一。本研究对滁河汊河集船闸改造前的襄河口至马汊河段渔业资源进行调查,初步掌握滁河襄河口至马汊河段渔业资源现状,丰富滁河渔业资源本底资料,为河流类型水域鱼类群落时空分布格局及其形成机制探究提供参考,为滁河段今后评估船闸改建对水生生物的影响提供理论支撑,同时为滁河鱼类物种多样性保护与管理提供科学依据。
1 材料与方法 1.1 采样方法于2015年12月和2016年5月对滁河襄河口至马汊河段鱼类群落进行调查,在以汊河集船闸为节点的4个辐射状河段设置20个采样点(图 1),分为上游河段,汊河集至襄河口,河道全长约39 km,1~5号样点分布在上游河段;下游河段,汊河集至马汊河口,河道全长约25 km,6~10号样点分布在下游河段;来安河段,汊河集至来安河水口,河道全长约21 km,11~15号样点分布在来安河段;清流河段,汊河集至清流河谢郢,河道全长约36 km,16~20号样点分布在清流河段。利用鱼簖和地笼在各河段开展渔业资源调查,鱼簖拦网长50 m,网高3 m,囊网网目尺寸1 cm,放网时间为24 h;地笼网高和网宽均为30 cm,网长50 m,网目0.8 mm,放网时间为24 h。渔获物收集后立即放置于冰盒内,带回实验室依据相关文献[9-10]鉴定到种,利用数显游标卡尺和电子天平测定所有鱼类体长和体质量。体长精确至0.01 mm,体质量精确至0.1 g。

|
图 1 渔业资源采样断面图 Fig. 1 Distribution of sampling stations in Chuhe River |
按照鱼类的栖息环境和洄游方式,调查水域鱼类存在3种生态类群[11]。参照鱼类食性文献[9-10]和食性类型划分方法,将滁河水域鱼类分为肉食性、杂食性、植食性、浮游食性和碎屑食性等5类。根据已有文献[12],将初次性成熟小于2龄,最大体长小于24 cm的鱼类划为小型鱼类。根据鱼类在垂直空间的分布差异[11],将其分为中上层、中下层和底层鱼类。
1.2.2 鱼类优势种鱼类优势种利用相对重要性指数[13](index of relative importance, IRI)进行表征,公式为
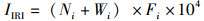 (1)
(1)
式中:IIRI为相对重要性指数;Ni为第i种鱼的尾数占总尾数的百分比,%;Wi为第i种鱼的质量占总质量的百分比,%;Fi为第i种鱼出现次数占总调查样点的百分比,%。
将IRI≥500的物种定为优势种,100≤IRI < 500的物种定为常见种,10≤IRI < 100的物种定为一般种,IRI < 10的物种定为少见种[14]。
1.2.3 生物多样性指数鱼类物种多样性指数包括Shannon-Weiner多样性指数(H)[15]、Wilhm丰富度指数(H′) [16]、Pielou均匀度指数(E) [17]、Simpson优势度指数(C) [14],计算公式为
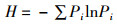 (2)
(2)
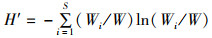 (3)
(3)
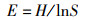 (4)
(4)
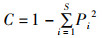 (5)
(5)
式中: Pi为鱼类群落中第i种鱼的个体数占所有鱼类总个体数的比例; W为渔获物总生物量; Wi为第i个物种生物量; S代表所有鱼类种类数。
采用单因素方差分析(One-way analysis of variance, ANOVA)对渔获物数量及质量、物种数季节及采样区域间的差异进行检验分析,若差异显著,再次进行TUKEY多重比较。利用SPSS 24.0进行数据统计分析。
2 结果 2.1 物种组成共采集渔获物20.05 kg,共计12 037尾,鱼类45种,隶属于6目13科36属(表 1)。渔获物以鲤形目(Cypriniformes)鱼类为主(表 1),共25属30种,占总物种数的55.56%;鲤形目渔获数量和质量在渔获物中均占优势地位,分别占总渔获数量和质量的78.12%和82.49%。鲈形目(Perciformes)渔获数量次之,共1 613尾,质量为4.60 kg,分别占渔获物总数量和总质量的13.40%和2.30%;鳗鲡目(Clupeiformes)物种数、渔获数量和质量均最少,分别占总数的2.17%、0.01%和0.03%(表 2)。渔获物中初次性成熟小于2龄,最大体长小于24 cm的小型鱼类有25种,占总物种数的55.56%(表 1)。空间上,4个河段共有种有18种,鱼类物种数呈显著差异(P < 0.001),其中上游段最多为34种,下游段次之, 32种,来安河最少,为26种。冬季和春季鱼类物种数分别为36种和37种,其中共有物种28种。

|
表 1 滁河水域鱼类物种组成和生态类群 Tab.1 Species composition and ecological guilds of fish community in Chuhe River |

|
表 2 滁河水域鱼类物种、数量和质量百分比 Tab.2 Percentage of fish species, quantity and mass in Chuhe River |
调查水域淡水定居性鱼类最多,共计41种,占总物种数的89.13%,江湖半洄游鱼类仅采集到鲢1种,出现于滁河上游、下游河段,江海洄游性鱼类2种,不同河段物种洄游类型鱼类种类组成差异不显著(P=0.10)。从栖息水层看,调查水域底层鱼类占优势,有21种,中上层和中下层鱼类相对较少,分别有13和12种,不同河段同一栖息水层的鱼类物种组成无显著差异(P=0.33);从摄食习性看,调查水域鱼类以杂食性和碎屑食性为主,占总物种数63.04%;植食性鱼类仅2种,出现于滁河上游、下游河段,不同河段同一摄食特性鱼类种类组成无显著差异(P=0.03)。见图 2。

|
图 2 滁河水域鱼类生态类群 Fig. 2 Fish ecological guilds in Chuhe River |
总体来讲,滁河水域鱼类的相对重要性指数(IRI)值高于500的有鲫(C.auratus)、鲤(C.carpio)、棒花鱼(A.rivularis)和黄颡鱼(P.fulvidraco)4种,为滁河水域优势鱼类,占鱼类总数量的30.07%,占总质量的68.95%(表 3)。IRI在100~500间的常见种有6种,最常见的物种为淡水定居似鳊(P.simoni)和斑条鱊(A.taenianalis),光泽黄颡鱼(P.nitidus)和红鳍原鲌(C.erythropterus)为一般常见种。优势种随时间和空间变化有所差异,春、冬两季优势种各3种,共有优势种为鲫、鲤,春季黄颡鱼为群落优势种之一,而棒花鱼冬季在群落中占优势。上游段优势种为鲫、鲤、光泽黄颡鱼和斑条鱊,而下游段为鲫;来安河段、清流河段优势种均为鲫和棒花鱼、鲤和黄颡鱼。

|
表 3 滁河水域渔业生物优势种组成 Tab.3 Composition of dominant fish species in Chuhe River |
滁河水域渔获数量群落特征值分别如下:丰富度指数(H′)为4.92,Shannon-Wiener指数(H)为2.22,优势度指数(C)为0.19,均匀度指数(E)为0.18。滁河鱼类多样性水平春季低于冬季,具体表现为优势度高,均匀度低,但季节间差异不明显。鱼类群落多样性的空间差异较小:上游段多样性水平最高,H为2.18;下游河段最低,H为2.05。见表 4。

|
表 4 滁河水域鱼类物种多样性指数 Tab.4 Species diversity in dices of fish communities in Chuhe River |
运用ANOSIM检验时空因素对鱼类群落结构的影响,结果表明:鱼类群落结构具有显著时空变化(P < 0.05);不同季节间鱼类群落存在明显重叠,几乎不分离(r=0.15),不同河段间鱼类群落分离程度相对较高(r=0.20)。不同河段群落结构的两两比较结果显示:上游段渔获物群落结构与其他河段间均存在显著差异(P < 0.05),而其他两两河段间鱼类群落结构差异不显著(P>0.05)。上游段与其他各河段间、下游段与来安河段群落分离程度相对较高(r=0.25),其中,上游段与来安河段分离程度最高(r=0.48),而清流河段与下游段、来安河段群落结构几乎不分离(r < 0.25)。见图 3。

|
图 3 滁河水域渔获物群落结构差异 Fig. 3 Variation of fish community structure in Chuhe River |
鱼类群落结构空间差异分析显示,上游段和下游段群落结构存在显著性差异。运用相似性百分比(SIMPER)分析造成上、下游段群落结构差异的关键贡献物种(累计贡献率达90%的物种),结果显示造成两河段间群落差异的主要物种有27种(如表 5),其中,上游段关键种是斑条鱊、鲫、光泽黄颡鱼,下游段为似鳊、鲫、斑条鱊,光泽黄颡鱼、棒花鱼丰度变化的贡献率均较高,是造成两河段群落组成差异的最主要物种。

|
表 5 滁河渔获物群落结构空间差异 Tab.5 Spatial variationof fish community structure in Chuhe River |
渔获物以鲤形目鱼类为优势种,单尾均重 < 50 g的小型鱼类占渔获物总数的74%,在渔获物群落中占绝对优势,这与其他水域相关研究[9, 18-19]结果类似。滁河襄河口至马汊河段水域鱼类调查历史文献觤较少,本次调查共计鉴定出鱼类45种,高于相邻怀洪新河太湖银鱼保护区的37种和淮河荆涂峡鲤、长吻
与湖泊、水库等静水系统不同,河流系统受水文、水位等的变动,往往具有显著季节性变动,因此会对流域内物种的群落组成及数量产生重要影响[28-29]。此外物种自身的繁殖、死亡等周期性生活史事件也会影响群落结构及其多样性[30]。本次调查发现,冬季渔获重量远高于春季,主要是由于冬季水温低,鱼类等迁移至深水区越冬,活动强度弱,渔获物捕获率上升[31-32]。同时,经过秋季育肥后,物种规格变大,因此渔获重量高。春季水温升高,鱼类活动强度增强,物种集中于岸边浅水区繁殖,单次渔获物捕获量下降,并且经越冬期、繁殖期的消耗,物种质量降低[33]。渔获数量、群落多样性水平季节间差异也类似,表现为冬季大于春季,但群落多样性水平差异主要归因于优势物种种类较少,群落向少数优势种集中,群落均匀性下降,多样性水平低。冬季优势物种的群落优势度降低,群落优势度分布相对均匀,多样性水平高,这与秋浦河、长江安庆段观察到的结果相似[12, 34]。调查发现,两季渔获物种数相同,但群落组成差异显著,经SIMPER分析发现,主要是由各物种的平均丰度差异引起的,其中光泽黄颡鱼、棒花鱼丰度变化贡献率较高(表 5),是造成春、冬两季群落组成差异的主要贡献物种。
3.3 渔业群落的空间格局在河流生态系统中,物种群落结构的复杂性与栖息地的稳定性、多样性成正相关[35]。本次调查发现,上游段渔业群落结构与清流河和来安河差异显著,可能是支流生境较为复杂、环境稳定,因此物种丰富多样。上游段与下游段渔业群落结构也存在显著差异,表现为上游段物种丰富,有研究指出,在自然环境下,河流下游物种较上游更为丰富多样[36],而本研究结果与之相反,说明调查河段生境遭到破坏,考虑到上游与下游间建有船闸(不通航),故推测船闸的存在是导致上下游物种数分布差异的非生物因素之一[37]。通过ANOSIM检验发现滁河上游、下游段物种群落组成存在显著的空间差异,可能与闸坝改变水域水文情势,破坏鱼类适宜栖息生境,阻断物种洄游通道有关[37]。SIMPER分析结果显示,闸坝通过改变上下游物种的平均丰度,限制鱼类自由活动空间,逃逸空间也受到巨大影响,同时也削弱了鱼类群落资源补偿、基因交流机制,最终导致闸坝上下游群落差异显著[38]。光泽黄颡鱼、棒花鱼丰度变化贡献率较高,是造成两河段差异的主要物种;小黄黝鱼、黑鳍鳈和光泽黄颡鱼在上游段的平均丰度分别为下游段的83倍、66倍、13倍,贝氏
| [1] |
何翠敏, 刘菊, 柏正林. 安徽滁河雨洪特性及与长江洪水遭遇组合分析[J]. 水科学与工程技术, 2015(4): 6-9. HE C M, LIU J, BAI Z L. Characteristics of storms and floods of Chuhe Rive and analysis of meeting together of the Yangtze River flood[J]. Water Sciences and Engineering Technology, 2015(4): 6-9. DOI:10.3969/j.issn.1672-9900.2015.04.002 |
| [2] |
王政祥, 丁志立. 滁河流域2003年梅雨期暴雨洪水分析[J]. 水资源研究, 2004, 25(4): 22-24, 28. WANG Z X, DING Z L. Analysis of the storm and flood of the Chuhe River basin in 2003 during the Meiyu Period[J]. Journal of Water Resources Research, 2004, 25(4): 22-24, 28. |
| [3] |
王庭槐. 自然地理志[M]. 南京: 南京出版社, 1992: 269-270. WANG T H. Local history compilation committee[M]. Nanjing: Nanjing Publishing House, 1992: 269-270. |
| [4] |
茹辉军, 刘学勤, 黄向荣, 等. 大型通江湖泊洞庭湖的鱼类物种多样性及其时空变化[J]. 湖泊科学, 2008, 20(1): 93-99. RUI H J, LIU X Q, HUANG X R, et al. Diversity of fish species and its spatio-temporal variations in Lake Dongting, a large Yangtze-connected lake[J]. Journal of Lake Sciences, 2008, 20(1): 93-99. DOI:10.3321/j.issn:1003-5427.2008.01.014 |
| [5] |
丁胜祥, 游中琼, 宁磊. 2016年滁河洪水对新修订洪水调度方案的检验[J]. 人民长江, 2017, 48(4): 32-36. DING S X, YOU Z Q, NING L. Verification of newly revised flood dispatching scheme by flood control practice of 2016 Chuhe River flood[J]. Yangtze River, 2017, 48(4): 32-36. |
| [6] |
杜艳, 袁学华. 滁河全椒段河流水质污染评价及驱动因子解析[J]. 安徽农业科学, 2017, 45(31): 72-75. DU Y, YUAN X H. Temporal variation of water quality and driving factors in Chuhe watershed of Quanjiao[J]. Journal of Anhui Agricultural Sciences, 2017, 45(31): 72-75. DOI:10.3969/j.issn.0517-6611.2017.31.024 |
| [7] |
柏正林, 程家友, 尹家星. 安徽省滁河流域悬移质泥沙分析[J]. 江淮水利科技, 2011(4): 33-34, 39. BAI Z L, CHENG J Y, YIN J X. Analysis of suspended load and sediment in Chuhe basin of Anhui province[J]. Jianghuai Water Resources Science and Technology, 2011(4): 33-34, 39. DOI:10.3969/j.issn.1673-4688.2011.04.017 |
| [8] |
吕平. 2015年滁河流域荒草圩蓄洪区行蓄洪水文还原分析[J]. 江淮水利科技, 2017(1): 46-48. LYU P. The analysis of the flood storage in the waste storage area of the Chuhe River basin in 2015[J]. Jianghuai Water Resources Science and Technology, 2017(1): 46-48. DOI:10.3969/j.issn.1673-4688.2017.01.020 |
| [9] |
朱松泉. 中国淡水鱼类检索表[M]. 南京: 江苏科学技术出版社, 1995. ZHU S Q. Synopsis of freshwater fishes of China[M]. Nanjing: Jiangsu Science and Technology Publishing House, 1995. |
| [10] |
伍汉霖, 邵广昭, 赖春福. 拉汉世界鱼类名典[M]. 基隆: 水产出版社, 1999. WU H L, SHAO G Z, LAI C F. Latin-Chinese dictionary of fishes names[M]. Jilong: Fisheries Press, 1999. |
| [11] |
茹辉军, 王海军, 赵伟华, 等. 黄河干流鱼类群落特征及其历史变化[J]. 生物多样性, 2010, 18(2): 169-176. RU H J, WANG H J, ZHAO W H, et al. Fishes in the mainstream of the Yellow River: assemblage characteristics and historical changes[J]. Biodiversity Science, 2010, 18(2): 169-176. |
| [12] |
张晓可, 于道平, 王慧丽, 等. 长江安庆段江豚主要栖息地鱼类群落结构[J]. 生态学报, 2016, 36(7): 1832-1839. ZHANG X K, YU D P, WANG H L, et al. Fish community structure in main habitat of the finless porpoise, the Anqing section of Yangtze River[J]. Acta Ecologica Sinica, 2016, 36(7): 1832-1839. |
| [13] |
PINKAS L, OLIPHANT M S, INVERSON I L K. Food habits of albacore, bluefin tuna, and bonito in California waters[M]. California: California Fish Game, 1971: 1-105.
|
| [14] |
陈国宝, 李永振, 陈新军. 南海主要珊瑚礁水域的鱼类物种多样性研究[J]. 生物多样性, 2007, 15(4): 373-381. CHEN G B, LI Y Z, CHEN X J. Species diversity of fishes in the coral reefs of South China Sea[J]. Biodiversity Science, 2007, 15(4): 373-381. DOI:10.3321/j.issn:1005-0094.2007.04.006 |
| [15] |
SHANNON C E, WEAVER W. The Mathematical theory of communication[M]. Urbana: University of Illinois Press, 1949.
|
| [16] |
WILHM J L. Use of biomass units in Shannon's formula[J]. Ecology, 1968, 49(1): 153-156. DOI:10.2307/1933573 |
| [17] |
PIELOU E C. Ecological diversity[M]. New York: Wiley, 1975: 1-165.
|
| [18] |
毛志刚, 谷孝鸿, 曾庆飞, 等. 太湖鱼类群落结构及多样性[J]. 生态学杂志, 2011, 30(12): 2836-2842. MAO Z G, GU X H, ZENG Q F, et al. Community structure and diversity of fish in Lake Taihu[J]. Chinese Journal of Ecology, 2011, 30(12): 2836-2842. |
| [19] |
李其芳, 严云志, 储玲, 等. 太湖流域河流鱼类群落的时空分布[J]. 湖泊科学, 2016, 28(6): 1371-1380. LI Q F, YAN Y Z, CHU L, et al. Spatial and temporal patterns of stream fish assemblages within Taihu Basin[J]. Journal of Lake Sciences, 2016, 28(6): 1371-1380. |
| [20] |
刘凯, 段金荣, 许萌霆, 等. 淮河荆涂峡鲤、长吻鮠国家级水产种质资源保护区冬季渔业群落多样性研究[J]. 安徽农业大学学报, 2015, 42(3): 424-428. LIU K, DUAN J R, XU M T, et al. Winter biodiversity of the fishery community in Huaihe Cyprinuscarpio and Leiocassislongirostris national aquatic germplasm resources conservation area[J]. Journal of Anhui Agricultural University, 2015, 42(3): 424-428. |
| [21] |
刘军, 曹文宣, 常剑波. 长江上游主要河流鱼类多样性与流域特征关系[J]. 吉首大学学报(自然科学版), 2004, 25(1): 42-47. LIU J, CAO W X, CHANG J B. The relationship between species diversity of fish and basin characteristics of main rivers in the upper reaches of the Yangtze River[J]. Journal of Jishou University (Natural Science Edition), 2004, 25(1): 42-47. DOI:10.3969/j.issn.1007-2985.2004.01.011 |
| [22] |
田佳丽, 王银平, 李佩杰, 等. 长江近口段近岸段鱼类群落多样性现状[J]. 上海海洋大学学报, 2021, 30(2): 320-330. TIAN J L, WANG Y P, LI P J, et al. Species diversity of fish in the section near the Yangtze river estuary[J]. Journal of Shanghai Ocean University, 2021, 30(2): 320-330. |
| [23] |
CABRAL H N, COSTA M J, SALGADO J P. Does the Tagus estuary fish community reflect environmental changes?[J]. Climate Research, 2001, 18(1/2): 119-126. |
| [24] |
毛志刚, 谷孝鸿, 龚志军, 等. 洪泽湖鱼类群落结构及其资源变化[J]. 湖泊科学, 2019, 31(4): 1109-1119. MAO Z G, GU X H, GONG Z J, et al. The structure of fish community and changes of fishery resources in Lake Hongze[J]. Journal of Lake Sciences, 2019, 31(4): 1109-1119. |
| [25] |
王银平. 太湖湖鲚种群特征及鱼类种间关系研究[D]. 北京: 中国科学院大学, 2016. WANG Y P. Studies on the population feature of Coilianasustaihuensis and inter specific relationship among fishes in Lake Taihu[D]. Beijing: University of Chinese Academy of Sciences, 2016. |
| [26] |
肖俊, 罗永巨. 我国鲫鱼研究进展[J]. 广西水产科技, 2010(4): 12-18. XIAO J, LUO Y J. Research progress of cruciancarp in China[J]. Guangxi Fisheries Science and Technology, 2010(4): 12-18. |
| [27] |
PERGA ME, ARFI R, GERDEAUX D. Seasonal variations in fish δ13C and δ15N in two West African reservoirs, Sélingué and Manantali (Mali): modifications of trophic links in relation to water level[J]. Isotopes in Environmental and Health Studies, 2005, 41(2): 109-123. DOI:10.1080/10256010500131734 |
| [28] |
CHADWICK M A. Stream ecology: Structure and function of running waters[J]. Freshwater Biology, 2008, 53(9): 1914-1914. |
| [29] |
LEIRA M, CANTONATI M. Effects of water-level fluctuations on lakes: an annotated bibliography[J]. Hydrobiologia, 2008, 613(1): 171-184. DOI:10.1007/s10750-008-9465-2 |
| [30] |
朱仁, 司春, 储玲, 等. 基于栖息地斑块尺度的青弋江河源溪流鱼类群落的时空格局[J]. 水生生物学报, 2015, 39(4): 686-694. ZHU R, SI C, CHU L, et al. The spatio-temporal distribution of fish population in the head waters of the Qingyi river: a study based on the habitat patchs[J]. Acta Hydrobiologica Sinica, 2015, 39(4): 686-694. |
| [31] |
周兴安, 乔永民, 王赛, 等. 洱海鱼类群落结构特征及其与环境因子关系[J]. 生态学杂志, 2016, 35(6): 1569-1577. ZHOU X A, QIAO Y M, WANG S, et al. Characteristics of fish assemblages associated with environmental factors in the Lake Erhai[J]. Chinese Journal of Ecology, 2016, 35(6): 1569-1577. |
| [32] |
曲慧敏, 王蕾, 邱盛尧. 乳山近海渔业资源种类组成及变化[J]. 水产科学, 2017, 36(3): 323-329. QU H M, WANG L, QIU S Y. Species composition and variation in fishery resources in the coastal Rushan waters[J]. Fisheries Science, 2017, 36(3): 323-329. |
| [33] |
何光喜, 张峻德, 刘其根, 等. 基于单层刺网渔获物的千岛湖鱼类群落结构初步研究[J]. 水产科技情报, 2011, 38(4): 197-203. HE G X, ZHANG J D, LIU Q G, et al. Preliminary study on fish community structure in Qiandao Lake based on single-layer gillnets[J]. Fisheries Science & Technology Information, 2011, 38(4): 197-203. DOI:10.3969/j.issn.1001-1994.2011.04.010 |
| [34] |
王文剑, 储玲, 司春, 等. 秋浦河源国家湿地公园溪流鱼类群落的时空格局[J]. 动物学研究, 2013, 34(4): 417-428. WANG W J, CHU L, SI C, et al. Spatial and temporal patterns of stream fish assemblages in the Qiupu Headwaters National Wetland Park[J]. Zoological Research, 2013, 34(4): 417-428. |
| [35] |
陆健健. 河口生态学[M]. 北京: 海洋出版社, 2003. LU J J. Estuary ecology[M]. Beijing: China Ocean Press, 2003. |
| [36] |
王桂华. 水利工程对长江中下游江段鱼类生境的影响研究[D]. 南京: 河海大学, 2008. WANG G H. Study on the impact of water conservancy projects on fish habitat in the middle and lower reaches of the Yangtze River[D]. Nanjing: Hohai University, 2008. |
| [37] |
林鹏程, 高欣, 刘春池, 等. 葛洲坝上下游江段鱼类时空分布特征研究[J]. 长江流域资源与环境, 2011, 20(9): 1047-1053. LIN P C, GAO X, LIU C C, et al. Hydroacoustic surveys on temporal and spatial distribution of fishesin the upper and lower reaches of the Gezhouba Dam[J]. Resources and Environment in the Yangtze Basin, 2011, 20(9): 1047-1053. |
| [38] |
李捷, 李新辉, 贾晓平, 等. 连江鱼类群落多样性及其与环境因子的关系[J]. 生态学报, 2012, 32(18): 5795-5805. LI J, LI X H, JIA X P, et al. Relationship between fish community diversityand environmental factors in the Lianjiang River, Guangdong, China[J]. Acta Ecologica Sinica, 2012, 32(18): 5795-5805. |
2. National Demonstration Center for Experimental Fisheries Science Education, Shanghai Ocean University, Shanghai 201306, China