 2022,
Vol. 31
2022,
Vol. 31



2. 国家海洋局北海预报中心, 山东 青岛 266061;
3. 自然资源部第一海洋研究所, 山东 青岛 266071;
4. 烟台市海洋经济研究院, 山东 烟台 264004
浮游植物是海洋中最重要的初级生产者,地球上近50%的初级生产量由其提供[1]。它们生长迅速,分布范围广、对环境变化反应敏感。其丰度和种类会随着周围海域环境的变化迅速发生改变[2],因此浮游植物常被作为海洋生态系统的生物指示种[3]。日照近岸海域位于黄海中部,其近岸海域海洋生物资源十分丰富[4]。我国对于黄海浮游植物的研究自20世纪60年代至今多有报道[5-7]。这些研究对黄海海域浮游植物的数量、种类组成、季节变化作了详细的分析和研究,但对浮游植物与环境因子之间的关系研究较少,或仅限于温度、盐度和营养盐等。本文利用2016年冬夏两季的浮游植物和水质调查资料,对日照近岸海域的浮游植物群落结构和20个环境因子进行初步分析,以期找出影响日照近岸海域浮游植物群落的关键环境因子,进而为研究该海域的生态系统状况和环境治理提供基础数据。
1 材料与方法 1.1 研究区域研究区域设置在日照市近岸海域(119°35′E~ 119°45′E,35°20′N~35°35′N),该片海域于2011年被国家批准为第1批国家级海洋公园。为获取该海域全面、系统、综合的基础资料,于2016年2月25日—26日和7月30日—8月2日在该海域进行了浮游植物和水质调查,从近岸海域向远岸海域布设A、B、C、D、E、F等6个断面18个站位。见图 1。

|
图 1 浮游植物调查站位图 Fig. 1 Sampling stations of phytoplankton |
浮游植物样品使用浅水Ⅲ型浮游生物网(网长140 cm,网口内径37 cm,网口面积0.1 m2,筛绢规格JP80,孔径近似值0.077 mm)自离底2 m至表层垂直拖曳。样品用缓冲甲醛溶液(40%商用甲醛加入同体积蒸馏水,1 dm3 20%的甲醛溶液加100 g六次甲基四胺)固定后带回实验室用浓缩计数法进行种类鉴定并计数。水温、水深和盐度采用DS5X型多参数水质仪现场测定。水环境要素中油类用QCC-1型采水器采集,其他水环境要素均用击开式采水器采集。当水深≥15 m时,取表、中、底层;当水深在5 m和15 m之间时,取表、底层;当水深≤5 m时,只取表层;油类和重金属只取表层样品。其分析方法按《海洋监测规范》(GB 17378—2007)执行。
1.3 评价方法 1.3.1 生态优势度浮游植物群落生态优势度采用以下公式:
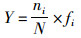 (1)
(1)
式中: Y为优势度;N为所有种的个体总数;ni为第i种的个体数;fi为第i种的出现频率,即该种出现的站位数与总站位数之比的百分数。将Y≥0.02的物种定为优势种[8]。
1.3.2 生物多样性利用Shannon-Wiener多样性指数(H′)[9]、Margalef的种类丰富度指数(D)[10]和Pielou均匀度指数(J)[11]来分析群落生物多样性。
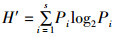 (2)
(2)
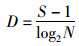 (3)
(3)
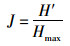 (4)
(4)
式(2)~式(4)中:S为浮游植物的种类总数;N为浮游植物总个数;Pi为第i种浮游植物占总个数的比例,Hmax为log2S。
1.3.3 群落结构及环境因子分析用Primer 5.0中的CLUSTER和MDS模块对浮游植物群落进行多元变量统计分析,在CLUSTER聚类分析时为减少机会种的影响,去除丰度过小的种类[12]。用SPSS 25.0对浮游植物丰度与环境因子做Pearson相关分析,在此之前用SPSS 25.0对水质环境因子做夏皮洛-威尔克正态性检测。若数据呈正态分布,则利用单因素ANOVA检验(显著水平为P<0.05)分析水质环境因子季节间有无显著差异;若数据不符合正态性,则利用单样本K-S检验(显著水平为P<0.05)分析水质环境因子季节间有无显著差异。
2 结果 2.1 环境因子特征本文所测水质环境因子共20项。其中冬季水温3.72~4.86 ℃,盐度30.77~31.60,pH 7.98~8.11,溶解氧(DO)质量浓度为7.67~8.00 mg/L,水深3.5~23.4 m,化学需氧量(COD)质量浓度为0.75 ~1.69 mg/L,油类质量浓度为0.012~0.082 mg/L。夏季水温25.39~28.99 ℃,盐度30.69~31.40,pH7.84~8.10,溶解氧质量浓度为6.72~8.13 mg/L,水深5.7~23.9 m,化学需氧量质量浓度为0.87 ~8.34 mg/L,油类质量浓度为0.016~0.099 mg/L。冬夏两季总悬浮物和营养盐水质监测结果见表 1、表 2。从分布趋势看COD、硝酸盐(NO3-N)、亚硝酸盐(NO2-N)、总无机磷(DIP)、和重金属中镉(Cd)、铜(Cu)、锌(Zn)汞(Hg)均呈由近岸向远海(A断面向F断面)逐渐降低的趋势。盐度、DO分布趋势与之相反。其他环境因子分布趋势不明显。夏皮洛-威尔克正态性检测显示数据呈正态分布的环境因子有水深、盐度、pH、铵盐(NH4-N)、油类和重金属中的铅(Pb)、汞(Hg)。单因素ANOVA检验和单样本K-S检验显示所测环境因子冬夏两季有显著差异的为水温、pH、COD、DO、总无机氮(DIN)、总铬(Cr)、砷(As)。

|
表 1 日照近岸海域冬季总悬浮物、营养盐和重金属监测结果 Tab.1 Monitoring results of total suspended solids, nutrients and heavy metals in the coastal waters of Rizhao in winter |

|
表 2 日照近岸海域夏季总悬浮物、营养盐和重金属监测结果 Tab.2 Monitoring results of total suspended solids, nutrients and heavy metals in the coastal waters of Rizhao in summer |
冬夏两季调查共鉴定浮游植物3门41属92种:硅藻门(Bacillariophy ta) 31属73种,甲藻门(Pyrrophy ta) 9属18种,金藻门(Chrysophy ta)1属1种,依次占总种数的79.35%、19.57%和1.09%。从属级上看:角毛藻属(Chaetoceros) 出现最多,为21种,占总种数的22.83%;海链藻属(Thalassiosira)和角藻属(Ceratium)均出现6种,各占6.52%;根管藻属(Rhizosolenia) 和原多甲藻属(Protoperidinium)均出现5种,各占5.44%;其他36属出现种类数均小于5种。从季节上看,冬季出现2门29属57种(硅藻门24属51种,甲藻门5属6种),夏季3门31属70种(硅藻门24属53种,甲藻门7属16种,金藻门1属1种)。
冬季A到F断面出现种类数A断面(39种)>C断面(36种)>B断面(29种)>F断面(25种)>E断面(24种)>D断面(22种);夏季A到F断面出现种类数C断面(41种)>A断面(38种)=D断面(38种)>B断面(34种)>F断面(32种)>E断面(27种)。从平面分布来看,冬夏两季出现种类数大致呈从近岸海域向远岸海域逐渐降低的趋势。
冬夏两季调查共出现优势种11种(表 2),其中, 冬季2种,夏季9种。两季优势种更替率为100.00%。在冬季航次中,柔弱几内亚藻(Guinardia delicatula)出现频率为1.00,优势度达0.80,在冬季航次中占据绝对优势;夏季航次中,除劳氏角毛藻(Chaetoceros lorenzianus)和深环沟角毛藻(Chaetoceros constrictus)优势度相对较高外,其他7个优势种优势度均小于0.1(表 3)。

|
表 3 日照近岸海域浮游植物优势种类及其优势度 Tab.3 Dominant species and its dominances of phytoplankton in the coastal waters of Rizhao |
冬夏两季调查日照近岸海域浮游植物丰度范围为1.71×105~6.12×107 cells/m3,均值为1.37×107 cells/m3(图 2)。浮游植物丰度季节变化较大:冬季丰度范围为2.20×106~6.12×107 cells/m3,平均值为2.61×107 cells/m3,这主要因为冬季调查出现了大量的柔弱几内亚藻,其丰度高达2.08×107 cells/m3。夏季丰度范围为1.71×105~5.70×106 cells/m3,平均值为1.22×106 cells/m3。从平面分布趋势来看:冬季C断面>B断面>A断面>E断面>D断面>F断面;最高值出现在调查海域中部近岸海域的A2号站,最低值出现在远岸的D1号站;高值区出现在A断面、B断面和C断面。夏季A断面>C断面>B断面>D断面>F断面>E断面;最高值出现在调查海域北部近岸海域的A1号站,最低值出现在中部近岸海域的B2号站;高值区出现在调查海域北部靠近吉利河口海域。冬夏两季浮游植物丰度分布总体上呈从近岸海域向远岸海域逐渐降低的趋势。

|
图 2 日照近岸海域浮游植物丰度平面分布 Fig. 2 Abundance distribution of phytoplankton in the coastal waters of Rizhao |
冬夏两季调查日照近岸海域浮游植物生物多样性指数(H′)为0.48~4.08,平均值为2.20(图 3)。日照近岸海域冬季浮游植物生物多样性指数明显低于夏季,变化范围为0.48~1.45,平均值为0.97;高值区出现在调查海域南部,最高值出现在南部远岸的F1号站,最低值出现在北部近岸的B1号站,整体呈南高北低的趋势;夏季多样性指数(H′)平面分布比较均匀,远岸稍高于近岸海域。两季调查均匀度指数(J)为0.11~0.88,平均值为0.50(图 4)。冬夏两季差别较大:冬季均匀度指数整体处在一个较低的水平,其中B断面3个站位均匀度指数平均值仅为0.17,最高的F断面平均值为0.27;夏季均匀度指数平面分布较均匀,为0.67~0.88,平均值为0.77。两季调查丰富度指数(D)为1.13~4.52,平均为2.53(图 5)。和多样性指数的平面分布相似,冬季明显低于夏季,冬季呈南北高中间低趋势分布,平均值为1.66;夏季平均值为3.40,平面分布和冬季相反,呈中间高南北低的趋势。

|
图 3 日照近岸海域浮游植物多样性指数(H′)分布 Fig. 3 Distribution of Shannon diversity index(H′) in the coastal waters of Rizhao |

|
图 4 日照近岸海域浮游植物均匀度指数(J)分布 Fig. 4 The distribution of Pielou's evenness index (J) in the coastal waters of Rizhao |

|
图 5 日照近岸海域浮游植物丰富度指数(D)分布 Fig. 5 The distribution of Margalef's species richness index(D) in the coastal waters of Rizhao |
当相似性尺度为55%(图 6)时,冬季18个调查站位划分为3个群落,其中:群落Ⅱ仅包含A2一个站位;群落Ⅰ包含C1、C2、D1、D3、F2共5个站位;群落Ⅲ包含A1、A3、B1、B2、B3、C3、C4、C5、D2、E1、E2、F1共12个站位。当相似性尺度为45%(图 6)时,夏季18个调查站位划分为3个群落,其中:群落Ⅰ包含B2、C4两个站位;群落Ⅱ包含A3、B3、C5、D3、E1、E2、F1、F2共8个站位;群落Ⅲ包含剩余8个站位。图 7是以Bray-Curtis相似性测量为基础的调查海域18个站位浮游植物组成相似MDS图。在压力系数为0.08和0.11时,冬夏两季MDS排序和群落聚类分析结果一致。在MDS排序图中两个生物群落结构越相似,它们之间的距离越近,因此和聚类分析相比MDS排序能更直观地反映不同站位之间的相似性。

|
图 6 日照近岸海域浮游植物聚类分析图 Fig. 6 Dendrogram for the similarity of phytoplankton in the coastal waters of Rizhao |

|
图 7 日照近岸海域浮游植物群落MDS分析 Fig. 7 MDS for the similarity of phytoplankton in the coastal waters of Rizhao |
浮游植物丰度与环境因子的关系采用SPSS 25.0中的Pearson相关分析模块。表 4为冬夏季浮游植物细胞丰度与环境因子Pearson相关性分析结果,冬季日照近岸海域浮游植物的丰度和盐度(r=-0.476,P=0.046)存在显著的负相关关系(P<0.05)。夏季日照近岸海域浮游植物的丰度和水深(r=-0.483,P=0.043)、DO(r=-0.519,P=0.027)存在显著的负相关关系(P<0.05);和NO3-N(r = 0.734,P=0.001)、NO2-N(r=10.699,P=0.001)、铵盐(NH4+-N)(r =0.591,P=0.010)、DIN(r=0.755,P=0.000 3)及重金属中的Zn(r=0.718,P=0.001)呈极显著的正相关关系(P<0.01)。

|
表 4 冬夏季浮游植物细胞丰度与环境因子Pearson相关性 Tab.4 Pearson correlation between phytoplankton cell abundance and environmental factors in winter and summer |
本研究中浮游植物种类数、丰度均呈由近岸向外海逐渐降低的趋势。这与所测环境因子COD、NO3-N、NO2-N、DIP、和重金属中Cd、Cu、Zn、Hg分布趋势相同,与盐度、水深、溶解氧分布趋势相反。丰度与环境因子的Pearson相关分析表明调查海域的浮游植物丰度与盐度、水深、溶解氧呈显著的负相关关系,与NO3-N、NO2-N、NH4+-N、DIN和重金属中的Zn呈显著的正相关关系。两者分析结果基本一致。分析表明日照近岸海域浮游植物种类数、丰度随着盐度、溶解氧浓度的增大而减小,随着NO3-N、NO2-N和重金属中的Zn浓度的增大而增大。日照近岸海域地处中纬度地区,为温带季风气候,冬季寒冷,夏季高温。温度对浮游植物的生长和优势种类影响较大[13-17]。本文冬季调查占绝对优势的种类为温带近岸性种类(柔弱几内亚藻),夏季随着温度的升高,优势种类变为温带至热带近岸性种[劳氏角毛藻、三角角藻(Ceratium tripos)]和沿岸广温性种[深环沟角毛藻、窄隙角毛藻(Chaetoceros affinis)、扭链角毛藻(Chaetoceros tortissimus)等]。营养盐也是影响近岸浮游植物生长的关键因子[18-21]。海洋中的浮游植物按Redfield系数吸收营养盐,当海水中营养盐比值偏离Redfield系数时,那么某一相对低的含量元素就会成为一个限制因子[22]。冬季由于海水垂直对流强烈加上沿岸河流营养盐的汇入,营养盐一般不会成为限制因子。到了夏季,海水垂直对流变弱,表层营养盐被大量消耗,底层营养盐又不能充分利用,营养盐成为浮游植物生长的限制因子。此时营养盐和浮游植物丰度呈正相关关系,这与本研究的结果相吻合。本文夏季调查甲藻种类数(16种)远大于冬季调查甲藻种类数(6种),这一方面是因为甲藻可在低氮、低磷的环境中生长和繁殖[23],营养盐不能成为其限制因子,另一方面是因为甲藻比硅藻对低氧环境的适应能力更强。另外盐度和水深也是影响调查海域浮游植物丰度的环境因子。曲静等[24]在研究青岛南部近海春、秋季浮游植物群落结构时发现浮游植物丰度与水深成显著正相关;王小平等[25]在研究春季红海湾水域盐度与浮游植物关系时发现表层和底层浮游植物丰度与盐度分别呈正相关和负相关。本文研究结论与上述两项报道不尽相同,这种差异与调查海域范围及调查季节有关,即不同的调查海域、不同的季节对浮游植物的影响因子也不同。
3.2 群落结构对海域水环境状况的指示一般来说当环境改变时,生物的群落结构也会发生变化。环境的好坏可以通过生物的群落结构特征来评价,但用不同的方法对处在同一环境的生物群落结构的同一监测数据进行评价时结果也不尽相同[26]。Shannon-Wiener多样性指数(H′)常用于生物群落结构的研究来评价水体受污染的程度[27]。一般说来:H′≥3,轻污染或无污染;1≤ H′ < 3,中污染;0≤ H′ < 1,重污染[28]。依此判断,冬季18个调查站位中11个为重污染,8个为中污染,分别占总站位的61.11%、38.89%,11个重污染站位分布在A、B、C、D、E、F共6个断面。夏季3个站位为中污染,15个站位为轻污染或无污染,3个中污染站位分布在B、C 2个断面。根据调查海域同步水质中pH、DO、COD、油类、DIN、DIP调查结果,冬季A3站位的DIN,A1、A2、D1的DIP,A1、A2、B3、C1、C3、C4、D1站位的油类和夏季A1、C1站位的DIN,A1、B1、C4站位的DIP,B2、B3、C3、D3、E1、F1站位的油类超过国家Ⅰ类海水水质标准。冬夏两季营养盐和油类超标站位主要集中在近岸海域,进一步分析表明,两季海水质量状况并无明显差别:按照《山东省海岸带海洋资源综合调查》中综合质量指数与环境质量分级的关系两季调查海域水质均为尚清洁范围。很显然,多样性指数值的大小并不能正确反映调查海域的水环境状况。多样性指数值的大小受物种总数和均匀度的双重影响,种数越多,个体分布越均匀多样性指数越大。冬季调查由于出现了大量的柔弱几内亚藻致使个体分布极不均匀,夏季调查浮游植物种类数较多且个体分布较均匀。有报道[29]表明生物多样性指数和均匀度的这种密切联系在诸如捕食、竞争、演替等生态过程中均会有所体现。除此以外,浮游植物的多样性指数与所处环境所受到的扰动强度紧密相关。根据“中度扰动假设”理论[30],环境在无扰动环境和强扰动环境下,物种数量会降低,多样性指数会变小,中等强度的扰动环境有利于浮游植物群落多样性达到较高水平。研究海域在不同站位不同季节受到的扰动强度的差异也是造成多样性指数不能很好地反映调查海域水环境状况的原因之一。
多元变量统计分析是分析群落结构变化的较灵敏的另一种方法。聚类分析结果:冬季群落Ⅱ包含的A2站位距岸最近且水质中DIN、油类均超过国家Ⅰ类海水水质标准,为18个站位中污染最为严重中的一个站位;群落Ⅰ包含的5个站位中C1站位油类超标,D1站位DIP和油类超标;群落Ⅲ包含的12个站位中,A1站位DIP超标,A1、B3、C3和C4站位油类超标。从聚类划分和水质调查结果来看,群落Ⅱ为营养盐和油类均超标海域,群落Ⅰ群落Ⅲ为营养盐和油类超标较轻或不超标海域。可见当海域的水环境污染(主要指营养盐和油类)较重时,聚类分析的这种群落划分可以很好地反映水质环境的变化,当污染较轻时,聚类分析不能通过群落划分的方式反映水质环境的变化情况。从夏季聚类划分和水质调查结果来看:2个DIN超标站位属于群落Ⅲ;3个DIP超标站位2个属于群落Ⅲ,1个属于群落Ⅰ;6个油类超标站位1个属于群落Ⅰ,4个属于群落Ⅱ,1个属于群落Ⅲ。相比于冬季聚类结构,夏季群落结构能更好地反映水环境污染状况:污染较严重站位属于群落Ⅰ,油类超标站位大部分集中在群落Ⅱ,营养盐超标站位大部分集中在群落Ⅲ。但是夏季调查海域浮游植物群落结构的划分也不能完全反映水环境质量状况,这主要是因为除了营养盐、油类这些污染因子外,浮游植物群落的结构还与水环境的温度、DO、盐度等因子以及海域所处地理位置有关。相对而言在日照近岸海域多元统计分析比Shannon-Wiener多样性指数(H′)分析能更好的反应水环境质量状况。
| [1] |
孙军. 海洋浮游植物与生物碳汇[J]. 生态学报, 2011, 31(18): 5372-5378. SUN J. Marine phytoplankton and biological carbon sink[J]. Acta Ecologica Sinica, 2011, 31(18): 5372-5378. |
| [2] |
BROGUEIRA M J, OLIVEIRA M D R, CABEÇADAS G. Phytoplankton community structure defined by key environmental variables in Tagus estuary, Portugal[J]. Marine Environmental Research, 2007, 64(5): 616-628. DOI:10.1016/j.marenvres.2007.06.007 |
| [3] |
GHINAGLIA L T, HERRERA-SILVEIRA J A, COMÍN F A. Structural variations of phytoplankton in the coastal seas of Yucatan, Mexico[J]. Hydrobiologia, 2004, 519(1/3): 85-102. |
| [4] |
王尽文, 黄娟, 陶卉卉, 等. 石臼港近岸海域春季鱼卵仔稚鱼调查研究[J]. 安徽农业科学, 2020, 48(21): 91-94. WANG J W, HUANG J, TAO H H, et al. Investigation on ichthyoplankton in offshore area of Shijiu Port in spring[J]. Journal of Anhui Agricultural Sciences, 2020, 48(21): 91-94. DOI:10.3969/j.issn.0517-6611.2020.21.024 |
| [5] |
朱树屏, 郭玉潔. 烟台、威海鲐鱼渔场及其附近海区角毛硅藻属的研究-Ⅰ. 分类的研究[J]. 海洋与湖沼, 1957, 1(1): 27-36. ZHU S P, GUO Y J. Studies on the genus Chaetoceros ehrenberg from the fishing ground of the mackerel, Pneumatophorus japonicus(Houttuyn), off the shantung coast from chefoo to Weihai part 1. a systematic study[J]. Oceanologia et Limnologia Sinica, 1957, 1(1): 27-36. |
| [6] |
王勇, 焦念志. 北黄海浮游植物营养盐限制的初步研究[J]. 海洋与湖沼, 1999, 30(5): 512-518. WANG Y, JIAO N Z. The primary research of nutrient limitation of phytoplankton in Northern Yellow Sea[J]. Oceanologia et Limnologia Sinica, 1999, 30(5): 512-518. DOI:10.3321/j.issn:0029-814X.1999.05.008 |
| [7] |
纪昱彤, 王宁, 陈洪举, 等. 2013年秋季渤黄海浮游植物的群落特征[J]. 中国海洋大学学报, 2018, 48(s2): 31-41. JI Y T, WANG N, CHEN H J, et al. Phytoplankton community structure in the Bohai and the Huanghai in autumn 2013[J]. Periodical of Ocean University of China, 2018, 48(s2): 31-41. |
| [8] |
陈亚瞿, 徐兆礼. 南黄海、东海鲐鲹鱼索饵场浮游动物生态特征[J]. 应用生态学报, 1990, 1(4): 327-332. CHEN Y Q, XU Z L. Ecological characteristics of zooplankton in feeding ground of mackerel and scads form South Yellow Sea and East China Sea[J]. Journal of Applied Ecology, 1990, 1(4): 327-332. |
| [9] |
SHANNON C E, WEAVER W. The mathematical theory of communication[M]. Urbana: The University of Illinois Press, 1949: 1-117.
|
| [10] |
MARGALEF R. Information theory in ecology[J]. General Systems, 1958, 3: 36-71. |
| [11] |
PIELOU E C. Ecological diversity[M]. New York: John Wiley & Sons Inc., 1975: 4-49.
|
| [12] |
FIELD J G, CLARKE K R, WARWICK R M. A practical strategy for analysing multispecies distribution patterns[J]. Marine Ecology Progress Series, 1982, 8(1): 37-52. |
| [13] |
龚玉艳, 张才学, 孙省利, 等. 2010年夏季雷州半岛海岸带浮游植物群落结构特征及其与主要环境因子的关系[J]. 生态学报, 2012, 32(19): 5972-5985. GONG Y Y, ZHANG C X, SUN X L, et al. Community characteristics of phytoplankton in the coastal area of Leizhou Peninsula and their relationships with primary environmental factors in the summer of 2010[J]. Acta Ecologica Sinica, 2012, 32(19): 5972-5985. |
| [14] |
王妍, 张永, 王玉珏, 等. 胶州湾浮游植物的时空变化特征及其与环境因子的关系[J]. 安全与环境学报, 2013, 13(1): 163-170. WANG Y, ZHANG Y, WANG Y J, et al. Spatial and temporal characteristics of phytoplankton and its relation with the environmental factors in Jiaozhou Bay[J]. Journal of Safety and Environment, 2013, 13(1): 163-170. DOI:10.3969/j.issn.1009-6094.2013.01.036 |
| [15] |
杨晓改. 海州湾及其邻近海域浮游生物群落结构及其与环境因子的关系[D]. 青岛: 中国海洋大学, 2015. YANG X G. Community structure of plankton in Haizhou Bay and adjacent waters and their relationships with environmental factors[D]. Qingdao: Ocean University of China, 2015. |
| [16] |
张家路, 王银平, 蔺丹清, 等. 扬中夹江浮游植物种群特征及其对环境因子的响应[J]. 上海海洋大学学报, 2019, 28(4): 607-615. ZHANG J L, WANG Y P, LIN D Q, et al. Phytoplankton community structure in Jiajiang in Yangzhong Estuary and its relation to environmental factors[J]. Journal of Shanghai Ocean University, 2019, 28(4): 607-615. |
| [17] |
上官欣欣, 薛俊增, 吴惠仙. 长江口浮游植物群落结构的特征[J]. 上海海洋大学学报, 2019, 28(5): 689-698. SHANGGUAN X X, XUE J Z, WU H X. Ecological features of phytoplankton community in the Yangtze River Estuary[J]. Journal of Shanghai Ocean University, 2019, 28(5): 689-698. |
| [18] |
胡好国, 万振文, 袁业立. 南黄海浮游植物季节性变化的数值模拟与影响因子分析[J]. 海洋学报, 2004, 26(6): 74-88. HU H G, WAN Z W, YUAN Y L. Simulation of seasonal variation of phytoplankton in the southern Huanghai (Yellow) Sea and analysis on its influential factors[J]. Acta Oceanologica Sinica, 2004, 26(6): 74-88. DOI:10.3321/j.issn:0253-4193.2004.06.009 |
| [19] |
杨世民, 董树刚, 汤志宏. 2005年夏季福建罗源湾浮游植物群落结构特征[J]. 海洋湖沼通报, 2010(2): 101-108. YANG S M, DONG S G, TANG Z H. Phytoplankton community of the Luoyuan bay in summer 2005[J]. Transactions of Oceanology and Limnology, 2010(2): 101-108. DOI:10.3969/j.issn.1003-6482.2010.02.017 |
| [20] |
姜庆宏, 王佳宁, 李卫平, 等. 包头南海湖非冰封期浮游植物的时空动态特征[J]. 水生态学杂志, 2020, 41(1): 30-36. JIANG Q H, WANG J N, LI W P, et al. Temporal and spatial dynamics of phytoplankton in Nanhai lake, Baotou, during the ice-free period[J]. Journal of Hydroecology, 2020, 41(1): 30-36. |
| [21] |
杨娜, 段元亮, 包炎琳, 等. 杭州湾人工潟湖夏季浮游植物群落演替与环境因子的关系[J]. 上海海洋大学学报, 2020, 29(5): 757-769. YANG N, DUAN Y L, BAO Y L, et al. Relationship between summer phytoplankton community succession and environment factors in the artificial lagoon of Hangzhou bay[J]. Journal of Shanghai Ocean University, 2020, 29(5): 757-769. |
| [22] |
HECKY R E, KILHAM P. Nutrient limitation of phytoplankton in freshwater and marine environments: a review of recent evidence on the effects of enrichment[J]. Limnology and Oceanography, 1988, 33(4): 796-822. |
| [23] |
张凡. 北黄海海洋牧场及周边海域浮游植物时空变动的研究[D]. 威海: 山东大学, 2019. ZHANG F. Study on the seasonal and variation of phytoplankton around marine ranching area of the Northern Yellow Sea[D]. Weihai: Shandong University, 2019. |
| [24] |
曲静, 宫相忠, 邢永泽, 等. 青岛南部近海春、秋季浮游植物群落结构的初步研究[J]. 中国海洋大学学报, 2009, 39(s1): 99-104. QU J, GONG X Z, XING Y Z, et al. Preliminary study on phytoplankton community structure in coastal waters south of Qingdao in spring and autumn[J]. Periodical of Ocean University of China, 2009, 39(s1): 99-104. |
| [25] |
王小平, 贾晓平, 林钦, 等. 红海湾养殖水域营养盐消长与温度、盐度、浮游植物量的关系[J]. 湛江海洋大学学报, 1999, 19(2): 18-24. WANG X P, JIA X P, LIN Q, et al. Relationship between nutrients, temperature, salinity and phytoplankton in the aquaculture waters of Honghai Bay[J]. Journal of Zhanjiang Ocean University, 1999, 19(2): 18-24. |
| [26] |
马藏允, 刘海, 王惠卿, 等. 底栖生物群落结构变化多元变量统计分析[J]. 中国环境科学, 1997, 17(4): 297-300. MA Z Y, LIU H, WANG H Q, et al. Multivariate analysis of community structure on macrobenthos[J]. China Environmental Science, 1997, 17(4): 297-300. DOI:10.3321/j.issn:1000-6923.1997.04.002 |
| [27] |
李永祺, 丁美丽. 海洋污染生物学[M]. 北京: 海洋出版社, 1991. LI Y Q, DING M L. Marine pollution biology[M]. Beijing: China Ocean Press, 1991. |
| [28] |
MAGNUSSEN S, BOYLE T J B. Estimating sample size for inference about the Shannon-Weaver and the Simpson indices of species diversity[J]. Forest Ecology and Management, 1995, 78(1/3): 71-84. |
| [29] |
郭劲松, 陈杰, 李哲, 等. 156 m蓄水后三峡水库小江回水区春季浮游植物调查及多样性评价[J]. 环境科学, 2008, 29(10): 2710-2715. GUO J S, CHEN J, LI Z, et al. Investigation of phytoplankton and assessment of algal diversity on backwater area of Xiaojiang river in Three Gorges reservoir after its initiate impounding to the water level of 156 m in spring[J]. Environmental Science, 2008, 29(10): 2710-2715. DOI:10.3321/j.issn:0250-3301.2008.10.004 |
| [30] |
BOITEAU R M, FITZSIMMONS J N, REPETA D J, et al. Detection of iron ligands in seawater and marine cyanobacteria cultures by high-performance liquid chromatography-inductively coupled plasma-mass spectrometry[J]. Analytical Chemistry, 2013, 85(9): 4357-4362. DOI:10.1021/ac3034568 |
2. The First Institute of Oceanography, MNR, Qingdao 266071, Shandong, China;
3. North China Sea Marine Forecasting Center of SOA, Qingdao Oceanic Environmental Monitoring Central Station of SOA, Qingdao 266061, Shandong, China;
4. Yantai Marine Economic Research Institute, Yantai 264004, Shandong, China