 2022,
Vol. 31
2022,
Vol. 31



2. 上海海洋大学 农业农村部鱼类营养与环境生态研究中心, 上海 201306;
3. 湖州海皇生物科技有限公司, 浙江 湖州 313105;
4. 上海希杰商贸有限公司, 上海 201228
大口黑鲈(Micropterus salmoides),又名加州鲈,是目前中国市场上主要的淡水肉食性鱼类之一,因适应性强、养殖周期短、肉质鲜美、营养价值高等特点,从而具有良好的市场前景。鱼粉作为饲料中重要的蛋白源,有蛋白含量高、适口性好、抗营养因子少、必需氨基酸组成平衡、钙磷碘硒含量高和维生素种类多等特点,是水产饲料中不可缺少的优质蛋白源[1],但因价格高,常被限制使用,从而使得替代鱼粉的动植物蛋白源开发研究一直方兴未艾。豆粕作为优质的植物蛋白之一,因蛋白质含量高、氨基酸丰富,有异黄酮等活性物质[2-3],是现阶段饲料行业中替代鱼粉蛋白的一种主要的植物蛋白原料。大口黑鲈饲料中蛋白质的水平较高(一般要>37%)[4]、鱼粉用量较大而提高了养殖成本,阻碍了大口黑鲈产业的快速发展,因而开展大口黑鲈饲料鱼粉替代研究意义重大。发酵豆粕在大口黑鲈配合饲料替代研究已有报道:王新霞[5]研究认为发酵豆粕替代鱼粉可有效提高加州鲈的特定生长率、饲料系数和蛋白质的表观消化率;赵鹏飞[6]发现16%的发酵豆粕替代鱼粉,大口黑鲈的增重率、特定生长率和蛋白效率达到最大值;JIANG等[7]的研究结果表明发酵豆粕替代40%的普通豆粕显著提高大口黑鲈的总末重、生长率和蛋白质效率。虽然目前有很多发酵豆粕替代鱼粉的研究,但以枯草芽孢杆菌全程有氧发酵的豆粕发酵产品的研究还少有报道。枯草芽孢杆菌是芽孢杆菌属的代表种,革兰氏阳性细菌。枯草芽孢杆菌在自然界中分布广泛,可以分泌多种胞外酶,在分解豆粕的过程中可以产生多种活性酶和枯草素等抗生素。枯草芽孢杆菌主要进行有氧呼吸,在营养复杂的环境下可进行无氧呼吸,但生长繁殖速度很弱。枯草芽孢杆菌作为豆粕发酵的常用菌种之一,根据其生活特性,充足的氧气持续供应能显著提高产品质量。普通发酵时,物料水分过低会导致氧气不足,降低豆粕发酵率,且发酵过程中物料粘连,基质多孔性降低,物料内的热量散发和氧气传递受到影响,抑制住菌的生长和繁殖[8]而影响豆粕发酵的质量。枯草芽孢杆菌有氧发酵得到的产品质量有很大的提升,相比于以厌氧乳酸菌为主的发酵豆粕产品,其抗营养因子特别是大分子蛋白(主要是抗原蛋白)的去除能力明显提高。本实验所用的枯草芽孢杆菌有氧发酵后的产品,分子质量为10~30 ku的小分子蛋白含量达26.38%(表 1)。

|
表 1 枯草芽孢杆菌有氧发酵对豆粕蛋白的降解情况 Tab.1 Degradation of soybean meal protein by anaerobic fermentation of lactic acid bacteria and aerobic fermentation of Bacillus |
本实验以枯草芽孢杆菌有氧发酵豆粕替代大口黑鲈饲料中不同梯度的鱼粉,从生长性能、饲料效率、消化酶活力和表观消化率等方面综合探究枯草芽孢杆菌全程有氧发酵豆粕替代鱼粉在大口黑鲈饲料中的可行性和适宜替代量,为发酵豆粕替代鱼粉的生产应用提供理论依据。
1 材料与方法 1.1 饲料的制备根据大口黑鲈的营养需求,设计了5组等氮等能的饲料(表 2),以鱼粉含量50%为基础饲料(FM),另外设计了4种发酵豆粕含量为10% (FSM10)、15%(FSM20)、20%(FSM30)和25%(FSM40)的实验饲料,替代鱼粉的量依次为10%、20%、30%和40%。发酵豆粕(表 3)由希杰商贸有限公司提供,其他原料由湖州海皇生物科技有限公司提供。原料经超微粉碎机粉碎后,60目筛网过筛,按饲料配方称重,逐级混匀,加入适量的水,混匀后过40目网筛,用环模式颗粒饲料机(HKJ-218,无锡市华明机器厂)制成直径2 mm的沉性颗粒饲料,烘箱60 ℃烘干15 h,置于-20 ℃冰箱保存备用。

|
表 2 实验饲料的配方与营养组成(%,干饲料) Tab.2 Formula and nutritional composition of experimental diets (%, dry diet) |

|
表 3 枯草芽孢杆菌有氧发酵豆粕营养组成 Tab.3 Nutritional composition and protein degradation of aerobic fermented soybean meal by Bacillus |
实验在上海海洋大学滨海基地进行,实验开始前,水泥池与网箱等用漂白粉消毒,幼鱼苗运到基地后暂养4周,最后1周用基础饲料进行驯化。暂养结束后,挑选形态正常,体质健康,平均体质量(19.83±0.33)g的幼鱼450尾随机放入15个网箱中。实验共分5个组,每组3个平行,每平行30尾。每天饱食投喂两次(07:00和18:00),投喂后1 h收集残饵。每周测定水质指标,养殖期间水温为(26±3.5)℃,pH为8.5±0.5,溶解氧质量浓度为4.0~5.0 mg/L,氨氮质量浓度为0~0.03 mg/L。养殖实验持续75 d。
1.3 样品采集实验开始前,共取6尾大口黑鲈幼鱼用于体组成分析;在实验期间,每次投喂结束2 h后开始收集粪便;实验结束后,停食24 h,分别统计每平行大口黑鲈的尾数和总重;每个平行取5尾鱼置于-20 ℃保存用于全鱼体组成分析;每平行取3尾鱼,取肠道、肝脏和胃用于消化酶活力测定。
1.4 测定指标与方法 1.4.1 生长指标实验期结束后捞出网箱中鱼,计总数并称重,计算饵料效益及特定生长率。
1.4.2 全鱼体成分分析粗蛋白采用凯氏定氮法测定;水分采用105 ℃恒温烘干失重法测定;粗脂肪采用氯仿-甲醇法测定;灰分采用马弗炉于550 ℃灼烧法测定。
1.4.3 消化率检测与分析饲料与粪便中的Cr2O3含量测定参照DIVAKARAN等[9]的方法,常规成分测定与体成分方法相同。
1.4.4 消化酶测定蛋白酶、脂肪酶和淀粉酶的测定根据南京建成试剂盒说明书进行,组织匀浆液的蛋白质质量浓度采用考马斯亮蓝染色法进行测定。
1.5 指标计算试验中使用的指标包括:饲料系数(feed coefficient ratio)、特定生长率(specific growth rate)、蛋白质效率(protein efficiency)、蛋白质沉积率(protein deposition rate)、粗脂肪沉积率(lipid deposition rate)、干物质表观消化率(apparent dry matter digestibility)、粗蛋白消化率(protein digestibility)和粗脂肪消化率(lipid digestibility)。计算公式:
 (1)
(1)
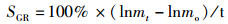 (2)
(2)
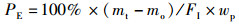 (3)
(3)
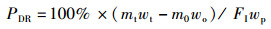 (4)
(4)
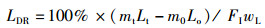 (5)
(5)
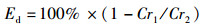 (6)
(6)
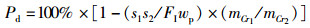 (7)
(7)
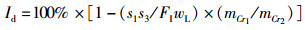 (8)
(8)
式中:FER为饲料系数,%;SGR为特定生长率,%/d;PE为蛋白效率;PDR为蛋白质沉积率,%;LDR为脂肪沉积率,%;Ed为干物质表观消化率,%;Pd为蛋白质表观消化率,%;Id为脂肪表观消化率,%;mo、mt为试验初始和终末鱼体质量, g;t为试验时间, d;wp为饲料粗蛋白质量分数, %;wt和wo分别为试验初始和终末鱼体蛋白质质量分数, %;wL为饲料粗脂肪质量分数, %;Lt和Lo分别为试验初始和终末鱼体蛋白质质量分数, %;mCr1和mCr2分别为饲料与粪便中Cr2O3含量,mg;FI为摄食量,g;s1为粪便的质量, g;s2和s3为粪便中粗蛋白和粗脂肪的质量分数, %。
1.6 数据处理实验数据用SPSS 21.0软件进行单因子方差分析(One-Way ANOVA, LSD),若存在显著差异则进行Duncan氏多重比较。所有统计值均以平均值±标准差(Mean±SD)表示,显著水平定为P < 0.05。
2 结果 2.1 发酵豆粕替代鱼粉对大口黑鲈生长性能的影响由图 1可知: 发酵替代鱼粉时,FSM20和FSM30组的均末质量和特定生长率(P < 0.05)均随着发酵豆粕替代鱼粉用量的增加呈现先上升后下降的趋势,在FSM20组时达到最大值; 各实验组的饲料系数并没有出现显著差异(P>0.05),FSM20和FSM30组保持一致,出现最低值。

|
柱状图上方的小写字母不同表示组间差异显著(P < 0.05)。 Different lower case letters at the top of the histogram indicate significant differences between groups (P < 0.05). 图 1 发酵豆粕替代鱼粉对大口黑鲈生长性能的影响 Fig. 1 Effects of fermented soybean meal instead of fish meal on growth performance of largemouth bass |
发酵豆粕替代鱼粉后大口黑鲈的摄食量显著提高(P < 0.05); 各实验组间蛋白质效率差异不显著(P>0.05)。蛋白质沉积率呈现先上升后下降的变化趋势,FSM20出现最大值,FSM40组的蛋白沉积率显著低于对照组(P < 0.05)。随着替代量的增加,脂肪沉积率呈现下降的变化趋势,FSM40组显著低于对照组。见表 4。

|
表 4 发酵豆粕替代鱼粉对大口黑鲈饲料效率的影响 Tab.4 Effect of fermented soybean meal instead of fish meal on feed efficiency of largemouth bass |
发酵豆粕替代鱼粉对大口黑鲈鱼体粗蛋白含量呈先上升后下降的变化,在FSM20组时出现最大值,但各组之间没有显著性差异(P>0.05);全鱼粗脂肪含量呈现下降变化趋势,并且FSM组显著低于FM组(P < 0.05); 全鱼水分FSM20组显著低于其他组(P < 0.05),其他各组之间差异不显著(P>0.05)。见表 5。

|
表 5 发酵豆粕替代鱼粉对大口黑鲈全鱼营养组成的影响 Tab.5 Effect of fermented soybean meal instead of fish meal on the nutritional composition of whole largemouth bass |
发酵豆粕替代鱼粉后,FSM30和FSM40组的干物质表观消化率显著低于对照组(P < 0.05);替代组与对照组之间蛋白质表观消化率无显著性差异(P>0.05);粗脂肪表观消化率虽出现下降的变化趋势,但并未出现差异性。见表 6。

|
表 6 发酵豆粕替代鱼粉对大口黑鲈表观消化率的影响 Tab.6 Effect of fermented soybean meal instead of fish meal on apparent digestibility of largemouth bass |
由表 7可知:随着发酵豆粕替代鱼粉含量的增加,肝脏与胃蛋白酶活力先上升后下降,皆在FSM20组达到最大值,其中胃蛋白酶在FSM30组开始显著性降低(P < 0.05);随着替代鱼粉含量的增加,肠道、肝脏和胃的脂肪酶活力呈下降的变化趋势,且胃脂肪酶的活力在FSM40组显著降低(P < 0.05); 肠道淀粉酶活力也随替代鱼粉的增加呈下降的变化趋势;肝脏的淀粉酶活力先上升后下降但并未出现显著性;胃淀粉酶活力FSM30组开始显著性降低。

|
表 7 发酵豆粕替代鱼粉对大口黑鲈消化酶活性的影响 Tab.7 Effect of fermented soybean meal instead of fish meal on enzyme activity of largemouth bass |
现有的研究表明,适量的发酵豆粕替代鱼粉可以提高水产动物的生长性能和饲料利用。杨艳玲等[10]发现用发酵豆粕替代14%的鱼粉,花鲈的末均质量和特定生长率显著增加;DING等[11]用发酵豆粕替代日本沼虾25%的鱼粉,饲料系数会优于对照组;何娇娇等[12]发现发酵豆粕替代45%鱼粉时会显著提高大黄鱼幼鱼的摄食量。
发酵豆粕替代鱼粉可以有效提高摄食量,主要因为发酵豆粕部分碳水化合物被微生物降解,结构变得疏松,提高了适口性。另一方面,豆粕蛋白在发酵过程中会产生小分子肽和游离的氨基酸,其中一部分肽会呈现为鲜味肽和甜味肽,一部分氨基酸表现为甜味氨基酸(苏氨酸、丙氨酸、甘氨酸)和鲜味氨基酸(谷氨酸、天门冬氨酸)会给发酵豆粕带来芳香气味和鲜味,提高其诱食性[13-15]。
本实验所用的芽孢杆菌有氧发酵豆粕中相对分子质量较小的蛋白含量丰富(< 10 ku蛋白达35.2%),其中有大量的小肽。相较于鱼粉中完整的蛋白质结构,这些小分子的蛋白在被降解为氨基酸的过程中会消耗较低的能量和营养物质,从而使得机体在调控蛋白质合成时达到节约能量的目的。另外,小肽可以被鱼类的胃和肠黏膜细胞完整地吸收,有着比游离氨基酸更快的吸收速度[16-17],小肽还可以直接参与蛋白质的合成并且可以对氨基酸残基的吸收有促进作用,推测这可能是适量的芽孢杆菌有氧发酵豆粕替代鱼粉后生长性能和蛋白质利用率提高的主要原因。本研究中发酵豆粕在提高蛋白质沉积率的同时,还降低了脂肪沉积率,防止了鱼体脂肪的过度堆积从而损害鱼体健康[18]。这是因为发酵豆粕中的大豆肽可以通过调节机体代谢起到消耗脂肪的作用,研究表明大豆肽可以活化交感神经,阻止脂肪吸收和促进脂肪分解代谢,此外大豆肽还可以刺激产热能的褐色脂肪组织(BAT)活性,提高血液中甲状腺素浓度,从而提高基础代谢水平,促进水产动物体内的多余的脂肪消耗[19]。
3.3 发酵豆粕替代鱼粉对大口黑鲈全鱼营养组成的影响本研究中,适量芽孢杆菌有氧发酵豆粕替代鱼粉并不会影响全鱼蛋白质含量。程成荣等[20]发现发酵豆粕替代50%鱼粉时罗非鱼全鱼蛋白质含量显著降低。全鱼蛋白质含量有所上升是因为芽孢杆菌有氧发酵豆粕中的肽类更有利于鱼体吸收利用,可以有效提高鱼体蛋白质的合成效率。本试验中适量的芽孢杆菌有氧发酵豆粕替代鱼粉会导致全鱼脂肪含量显著降低,该结果与脂肪沉积率变化趋势一致。造成这一结果的原因可能是:发酵豆粕中的肽类有着阻碍脂肪吸收的效果;同时大豆中的非淀粉多糖会降低脂类物质的乳化作用,影响脂肪吸收利用[21];一些非淀粉多糖有结合胆盐、脂类和胆固醇的能力,会影响小肠的脂类代谢[22]。祝焱彬等[23]发现发酵豆粕替代鱼粉会降低梭鱼鱼体的脂肪含量;GARCÍA-ORTEGA等[24]发现巨型石斑鱼随着发酵豆粕替代鱼粉含量的增加,鱼体的脂肪含量随之降低。
3.4 发酵豆粕替代鱼粉对大口黑鲈消化酶活力和表观消化率的影响不少研究都表明,发酵豆粕替代鱼粉会对大口黑鲈的消化酶活力和表观消化率产生影响。殷海成等[25]对黄河鲤的研究发现,发酵豆粕替代鱼粉可以有效提高蛋白酶活力。黄峰等[26]对斑点叉尾
| [1] |
姚俊鹏, 肖勤. 水产养殖中鱼粉替代蛋白源的研究进展[J]. 饲料研究, 2015(24): 39-46. YAO J P, XIAO Q. Research progress of fish meal substitute protein sources in aquaculture[J]. Feed Research, 2015(24): 39-46. |
| [2] |
KISHIDA T, ATAKI H, TAKEBE M, et al. Soybean meal fermented by Aspergillus awamori increases the cytochrome P-450 content of the liver microsomes of mice[J]. Journal of Agricultural and Food Chemistry, 2000, 48(4): 1367-1372. DOI:10.1021/jf9905830 |
| [3] |
JANNATHULLA R, DAYAL J S, AMBASANKAR K, et al. Effect of protein solubility of soybean meal on growth, digestibility and nutrient utilization in Penaeus vannamei[J]. Aquaculture International, 2017, 25(5): 1693-1706. DOI:10.1007/s10499-017-0147-9 |
| [4] |
李二超, 陈立侨. 大口黑鲈的营养需要研究进展[J]. 现代农业科技, 2011(21): 312-316, 318. LI E C, CHEN L Q. Research advances on nutrient requirements of Micropterus salmoides[J]. Modern Agricultural Science and Technology, 2011(21): 312-316, 318. DOI:10.3969/j.issn.1007-5739.2011.21.198 |
| [5] |
王新霞. 发酵豆粕替代鱼粉在加州鲈饲料中的研究[J]. 饲料与畜牧: 新饲料, 2009(1): 58-61. WANG X X. Research on the replacement of fish meal with fermented soybean meal in the feed of California sea bass[J]. Feed and Livestock: New Feed, 2009(1): 58-61. |
| [6] |
赵鹏飞. 发酵豆渣在大口黑鲈饲料中应用的初步研究[D]. 重庆: 西南大学, 2018. ZHAO P F. Preliminary study on the application of fermented soybean dregs in the feed of largemouth bass[D]. Chongqing: Southwest University, 2018. |
| [7] |
JIANG Y, ZHAO P F, LIN S M, et al. Partial substitution of soybean meal with fermented soybean residue in diets for juvenile largemouth bass, Micropterus salmoides[J]. Aquaculture Nutrition, 2018, 24(4): 1213-1222. DOI:10.1111/anu.12659 |
| [8] |
TECHARANG J, APICHARTSRANGKOON A. Physical, chemical and rheological parameters of pressurized swai-fish (Pangasius hypophthalmus) emulsion incorporating fermented soybeans[J]. Food and Bioproducts Processing, 2015, 94: 649-656. DOI:10.1016/j.fbp.2014.09.002 |
| [9] |
DIVAKARAN S, OBALDO L G, FORSTER I P. Note on the methods for determination of chromic oxide in shrimp feeds[J]. Journal of Agricultural and Food Chemistry, 2002, 50(3): 464-467. DOI:10.1021/jf011112s |
| [10] |
杨艳玲, 陈冰, 彭凯, 等. 发酵豆粕替代鱼粉及添加包膜氨基酸、植酸酶对花鲈幼鱼生长性能及养殖水质的影响[J]. 饲料工业, 2019, 40(10): 47-53. YANG Y L, CHEN B, PENG K, et al. Evaluation of replacing fish meal by fermented soybean meal, adding coated amino acids and phytase on the growth performance and water quality for Lateolabrax japonicus[J]. Feed Industry, 2019, 40(10): 47-53. |
| [11] |
DING Z L, ZHANG Y X, YE J Y, et al. An evaluation of replacing fish meal with fermented soybean meal in the diet of Macrobrachium nipponense: Growth, nonspecific immunity, and resistance to Aeromonas hydrophila[J]. Fish & Shellfish Immunology, 2015, 44(1): 295-301. |
| [12] |
何娇娇, 王萍, 冯建, 等. 发酵豆粕对大黄鱼生长、肠道结构及肠道微生物菌群的研究[J]. 水生生物学报, 2018, 42(5): 919-928. HE J J, WANG P, FENG J, et al. Effects of fermented soybean meal on the growth and intestinal histology and microbiota of juvenile large yellow croaker Larimichthys crocea[J]. Acta Hydrobiologica Sinica, 2018, 42(5): 919-928. |
| [13] |
NJOKU H O, OKEMADU C P. Biochemical changes during the natural fermentation of the African oil bean (Pentaclethra macrophylla) for the production of Ugba[J]. Journal of the Science of Food and Agriculture, 1989, 49(4): 457-465. DOI:10.1002/jsfa.2740490408 |
| [14] |
曾欢, 陶宁萍, 张晶晶, 等. 生物发酵对大口黑鲈的脱腥效果研究[J]. 上海海洋大学学报, 2021, 30(6): 1142-1152. ZENG H, TAO N P, ZHANG J J, et al. Effect of microbial fermentation on removing fishy odors in largemouth bass (Micropterus salmoides) meat[J]. Journal of Shanghai Ocean University, 2021, 30(6): 1142-1152. |
| [15] |
Subcommittee on Warmwater Fish Nutrition, Committee on Animal Nutrition, Board on Agriculture, National Research Council. Nutrient requirements of warmwater fishes and shellfishes[M]. Washington, D.C.: National Academy Press, 1983.
|
| [16] |
王建峰, 乐国伟. 小肽在动物营养中的应用[J]. 饲料工业, 2006, 27(7): 9-11. WANG J F, LE G W. Application of the small peptides in animal nutrition[J]. Feed Industry, 2006, 27(7): 9-11. |
| [17] |
张会平, 雷宁利. 小肽营养及应用研究[J]. 畜禽业, 2006(24): 20-23. ZHANG H P, LEI N L. The advance on the nutrition and utilization of small-pepeide[J]. Livestock and Poultry Industry, 2006(24): 20-23. |
| [18] |
HILLESTAD M, JOHNSEN F. High-energy/low-protein diets for Atlantic salmon: effects on growth, nutrient retention and slaughter quality[J]. Aquaculture, 1994, 124(1/4): 109-116. |
| [19] |
侯丽. 大豆多肽与运动医学研究进展[J]. 甘肃医药, 2012, 31(12): 907-909. HOU L. Research progress of soy peptides and sports medicine[J]. Gansu Medical Journal, 2012, 31(12): 907-909. |
| [20] |
程成荣. 杂交罗非鱼饲料中发酵豆粕和肉骨粉替代鱼粉的研究[D]. 广州: 中山大学, 2004. CHENG C R. Replacement of fish meal by fermented soybean meal and meat and bone meal in practical diets for juvenile hybrid tilapia, Oreochromis niloticus ♀×O. aureus ♂[D]. Guangzhou: Sun Yat-sen University, 2004. |
| [21] |
徐境羚, 赵宇红. 饲用酶制剂对饲料中非淀粉多糖的作用[J]. 吉林畜牧兽医, 2003(2): 4-6. XU J L, ZHAO Y H. Effect of feed enzyme preparation on non-starch polysaccharide in feed[J]. Jilin Animal Husbandry and Veterinary Medicine, 2003(2): 4-6. |
| [22] |
张永库, 王裕玉. 大豆非淀粉多糖研究进展[J]. 黑龙江水产, 2012(3): 39-42. ZHANG Y K, WANG Y Y. Research progress of soybean non-starch polysaccharide[J]. Heilongjiang Fisheries, 2012(3): 39-42. |
| [23] |
祝焱彬, 於叶兵, 杨文平, 等. 发酵豆粕替代鱼粉对梭鱼生长、血液生化指标和体成分的影响[J]. 盐城工学院学报(自然科学版), 2018, 31(4): 63-68. ZHU Y B, YU Y B, YANG W P, et al. Effects of fermented soybean meal replacing fish meal on growth, blood biochemical Index and body composition of barracuda[J]. Journal of Yancheng Institute of Technology (Natural Science Edition), 2018, 31(4): 63-68. |
| [24] |
GARCÍA-ORTEGA A, KISSINGER K R, TRUSHENSKI J T. Evaluation of fish meal and fish oil replacement by soybean protein and algal meal from Schizochytrium limacinum in diets for giant grouper Epinephelus lanceolatus[J]. Aquaculture, 2016, 452: 1-8. |
| [25] |
殷海成, 贾峰, 张哲, 等. 饲料中发酵豆粕替代鱼粉对黄河鲤生长及消化酶活性的影响[J]. 粮食与饲料工业, 2014(3): 48-51. YIN H C, JIA F, ZHANG Z, et al. Effects of replacement of dietary fish meal by fermented soybean meal on growth performance and digestive enzyme activity of Cyrinus carpio[J]. Cereal & Feed Industry, 2014(3): 48-51. |
| [26] |
黄峰, 李惠, 刘军, 等. 发酵豆粕替代鱼粉对斑点叉尾 消化酶活性的影响[J]. 水利渔业, 2008, 28(4): 38-40.HUANG F, LI H, LIU J, et al. Effect of replacing soybean meal with fermented soybean meal on digestive enzyme activity of Ictalurus punctatus[J]. Reservoir Fisheries, 2008, 28(4): 38-40. |
| [27] |
王崇, 雷武, 解绶启, 等. 饲料中豆粕替代鱼粉蛋白对异育银鲫生长、代谢及免疫功能的影响[J]. 水生生物学报, 2009, 33(4): 740-747. WANG C, LEI W, XIE S Q, et al. Effect of dietary replacement of fishmeal protein by soybean meal protein on Growth performance, metabolism and immunity of gibel carp (Carassius auratus gibelio)[J]. Acta Hydrobiologica Sinica, 2009, 33(4): 740-747. |
| [28] |
李惠, 黄峰, 胡兵, 等. 发酵豆粕替代鱼粉对斑点叉尾 生长和饲料表观消化率的影响[J]. 淡水渔业, 2007, 37(5): 41-44.LI H, HUANG F, HU B, et al. Effects of replacement of fish meal with fermented soybean in the diet for channel catfish (Ictalurus punctatus) on growth performance and apparent digestibility of feed[J]. Freshwater Fisheries, 2007, 37(5): 41-44. |
| [29] |
VISESSANGUAN W, BENJAKUL S, POTACHAREON W, et al. Accelerated proteolysis of soy proteins during fermentation of thua-nao inoculated with Bacillus subtilis[J]. Journal of Food Biochemistry, 2005, 29(4): 349-366. |
| [30] |
吕云云, 常青, 陈四清, 等. 发酵豆粕对圆斑星鲽生长及消化能力的影响[J]. 水生生物学报, 2016, 40(1): 10-18. LYU Y Y, CHANG Q, CHEN S Q, et al. The effects of dietary fermented soybean meal on the growth and digestive ability of spotted halibut, Verasper variegatus[J]. Acta Hydrobiologica Sinica, 2016, 40(1): 10-18. |
| [31] |
HARA H, NARAKINO H, KIRIYAMA S, et al. Induction of pancreatic growth and proteases by feeding a high amino acid diet does not depend on cholecystokinin in rats[J]. The Journal of Nutrition, 1995, 125(5): 1143-1149. |
| [32] |
BAEVERFJORD G, KROGDAHL A. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: a comparison with the intestines of fasted fish[J]. Journal of Fish Diseases, 1996, 19(5): 375-387. |
| [33] |
李向, 华雪铭, 魏翔, 等. 饲料中维生素D3含量对大口黑鲈生长和抗氧化能力的影响[J]. 上海海洋大学学报, 2021, 30(1): 94-102. LI X, HUA X M, WEI X, et al. Effects of dietary vitamin D3 on the growth and antioxidant capacity of largemouth bass (Micropterus salmoides)[J]. Journal of Shanghai Ocean University, 2021, 30(1): 94-102. |
| [34] |
MATHEW A G, FRANKLIN M A, UPCHURCH W G, et al. Influence of weaning age on ileal microflora and fermentation acids in young pigs[J]. Nutrition Research, 1996, 16(5): 817-827. |
| [35] |
吴莉芳, 秦贵信, 孙泽威, 等. 饲料中去皮豆粕替代鱼粉对埃及胡子鲇消化酶活力和肠道组织的影响[J]. 中山大学学报(自然科学版), 2010, 49(4): 99-105. WU L F, QIN G X, SUN Z W, et al. Effect of dietary dehulled soyabean meal replacing fish meal on the activity of digestive enzyme and the intestinal tissue of Clarias lazera[J]. Acta Scientiarum Naturalium Universitatis Sunyatseni, 2010, 49(4): 99-105. |
2. Shanghai Ocean University Fishery Nutrition and Environmental Ecology Research Center, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China;
3. Huzhou Haihuang Biological Technology Co., Ltd., Huzhou 313105, Zhejiang, China;
4. Shanghai Xijie Trading Company, Shanghai 201228, China