 2019,
Vol. 28
2019,
Vol. 28



2. 上海海洋大学 农业农村部鱼类营养与环境生态研究中心, 上海 201306;
3. 上海海洋大学 农业农村部淡水水产种质资源重点实验室, 上海 201306;
4. 青田愚公农业科技有限公司, 浙江 青田 323900
浙江青田的稻-鱼共生系统已有1 300多年的历史[1],2005年联合国粮农组织将其确定为全球重要农业遗产(global important agriculture heritage system, GIAHS)试点项目。作为全球首批GIAHS项目之一(亚洲的第一个GIAHS项目),传统的青田稻-鱼共生系统为联合国粮农组织启动全球范围的GIAHS计划奠定了基础,同时也引起了许多学者[2-3]的关注。与水稻单作相比,稻-鱼共生模式能大幅减少化肥和农药的使用[2, 4],有利于保护稻田生物多样性[5-6],稻-鱼共生系统也因此具有更好的稳定性和可持续性[2]。GIAHS倡导“动态保护”,这种基于保护层面的优化有利于青田稻-鱼共生系统的可持续发展,对这种传统农业方式在当地的传承和推广产生了积极的影响。
栽培密度对水稻生长发育和群体构成具有双重调控作用[7],其改变会引起稻田环境中温、光、水、肥等生态因子的变化,从而对水稻群体产量构成产生影响[8]。栽培密度低时,植株间空隙大,通风透光条件好,土壤养分充足,有利于水稻的分蘖和生长,有效穗数和成穗率都能得到有效保证;栽培密度高时,单位面积穗数增加,田间通风透光条件差,湿度大,病虫害发生严重,个体间的生存矛盾加剧,导致穗粒结构失调[7, 9-10]。水稻密度的控制可以通过调整每穴基本苗数、单位面积穴数或株行距配置方式来实现[11]。在实际生产中,水稻合理密植被认为是一种稳产的手段[12]。传统的青田稻-鱼共生模式考虑到养殖田鱼的活动空间,一般采用水稻稀植的方式,水稻株行距常选择40 cm×36 cm[1],但这种低密度栽培方式是否会影响水稻产量?吴敏芳等[13]研究发现30 cm×30 cm的水稻密度可以更加充分地利用系统生产力。适用于青田稻-鱼共生系统的最佳水稻密度具体如何科学地选择?为此,我们通过调整栽培株行距来控制水稻密度,研究不同株行距下水稻生长和产量构成的变化,记录了各栽培密度下青田田鱼(Cyprinus carpio var. color)的存活和生长情况,以期为传统的青田稻-鱼共生系统中最适水稻密度的选择提供一定科学依据。
1 材料与方法 1.1 实验区概况实验于浙江省青田县仁庄镇(28°01'N,120°31'E)进行,地处浙江省东南部,瓯江中下游,属于典型亚热带季风气候区,年平均气温18.3 ℃,年均日照1 712~1 825 h,降水量1 400~2 100 mm。稻田属于洪积性泥沙田,耕层厚度约20 cm[17]。
1.2 实验设计实验采用随机大田块设计,稻田种养殖模式均为稻-鱼共生模式。水稻栽插均为单穴双株,设置3个水稻密度处理组,即3种栽培株行距:20 cm×30 cm(以下简称“高密度”)、30 cm×30 cm(以下简称“中密度”)和40 cm×30 cm(以下简称“低密度”),株距为南北走向。每种密度处理各设置3个重复,共计9块稻田。
种养殖稻田均采用平板式稻田。各田块田埂用水泥砖砌成,深入地下30 cm,高出水面50 cm,能有效阻隔各田块之间的土壤养分交流。灌溉方式为山区梯田式流水灌溉,实验稻田对角各开一口,分别用作进水口和出水口,进出水口用铁丝网拦住,防止田鱼逃逸,实验田块水源一致。灌溉水位始终保持在15~20 cm。实验田块上方架设防鸟网。
水稻品种为“甬优17”,养殖鱼品种为青田田鱼(Cyprinus carpio var. color)。水稻于4月30日播种,5月21日移栽,10月15日收割。水稻移栽前对实验田块施用有机肥(猪粪)作为基肥,施用量为600 kg/hm2,水稻生长过程中不施追肥。水稻移栽稻田20 d(6月10日)后投放青田田鱼,田鱼鱼种来源一致,规格为50 g左右,投放密度为9 000尾/hm2。分别记录每块稻田投放田鱼的尾数和质量。田鱼投放3 d后开始投喂饲料,日投喂量为田鱼体质量的2%~5%,分早晚两次投喂,并根据田鱼生长和摄食情况同步增加饲料投喂量。
1.3 样品采集与测算水稻移栽后,在每块稻田随机选取2行,每行连续选择10穴水稻定点标记,每隔15天观察其分蘖数,并统计分析水稻分蘖动态。水稻生物量的计算方法为测算水稻地上部分的干物质:于水稻返青期、拔节期、抽穗期和成熟期在每块稻田随机选择长势一致的5穴水稻,取其地上部分洗净后60 ℃、48 h烘干并称量[14]。
水稻收割前,进行水稻茎秆构成各要素的测定,并根据中华人民共和国农业行业标准NY/T1300—2007(农作物品种区域试验技术规范水稻)和TSURUTA等[15]的方法测定水稻产量构成要素。水稻产量的测算方法为,在水稻收割前每块田随机选取20穴水稻,捋下稻谷,39 ℃、48 h烘干称量,进行推算。水稻产量构成主要计算公式如下:
 (1)
(1)
式中:NH为每公顷穴数;L1为水稻栽培株距,m;L2为水稻栽培行距, m。
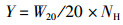 (2)
(2)
式中:Y为水稻产量;W20为20穴水稻质量;NH为每公顷穴数。
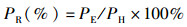 (3)
(3)
式中:PR为成穗率;PE为有效穗数;PH为最高苗。
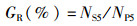 (4)
(4)
式中:GR为穗粒数;NS5为5穴总粒数;NP5为5穴总穗数。
 (5)
(5)
式中:SER为结实率;NFG为每穗实粒数;NTG为每穗总粒数。
田鱼于水稻收割前回捕。分别统计每块稻田回捕田鱼的尾数并称量,计算每块稻田田鱼的存活率SR和特定生长率SGR。计算公式如下:
 (6)
(6)
式中:RS为存活率;N2为回捕田鱼数量;N1为投放田鱼数量。
 (7)
(7)
式中:RSG为特定生长率;Wt2为回捕时田鱼的平均体质量;Wt1为投放时田鱼的平均体质量;D为整个养殖时间。
1.4 数据分析与统计水稻产量与水稻茎秆构成(株高、秆长、穗长、秆基部外径和穗基部外径)、产量构成因素(千粒重、有效穗数、成穗率、结实率、穗粒数、生物量和每公顷穴数)关系的分析采用灰色关联分析法(grey relational analysis, GRA)。灰色关联分析法是一种针对抽象系统的分析方法,其根据因素数据列的几何形状、发展态势接近的程度来衡量因素间关联程度[16]。该方法由邓聚龙教授于1982年提出,是灰色系统理论的重要组成部分。具体分析步骤如下:
(1)建立相关数列
将原始数据排列成相关数列,确定一个因变量因素(水稻产量)和多个自变量因素(水稻茎秆构成因素和产量构成因素)。
(2)原始数据标准化
根据公式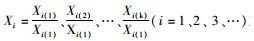
(3)计算绝对差值
分别计算水稻产量与水稻茎秆构成因子、产量构成因子的绝对差值Δi(k),Δi(k)=|X0(k)-Xi(k)|。
(4)计算关联系数
计算公式为
 (8)
(8)
式中:ξi(k)即为Xi(k)对X0(k)的关联系数,范围值为0 < ξi(k) < 1;ρ为分辨系数。
(5)计算关联度
计算公式为
 (9)
(9)
式中:n为比较序列的数据量,计算后进行相关性排序,即为水稻产量与各水稻茎秆构成因子、各产量构成因子之间的灰色关联度。
水稻相关数据均在Excel中建立数据库,并用SPSS 21.0统计软件进行单因素方差分析(One-way ANOVA)和非参数检验(Kruskal-Wallis),多重比较采用LSD法。P<0.05表示差异显著,P<0.01表示差异极显著。数据结果以平均值±标准误(Mean±SE)表示。
2 结果 2.1 不同水稻栽培密度下水稻分蘖动态变化规律“甬优17”属于晚稻品种,全生育期一般153 d左右。实验中,水稻生长各时期如下:播种育秧(4月30日),移栽(5月21日),返青期(5月25日),分蘖期(5月31日),拔节期(7月13日),抽穗期(8月19日),成熟期(9月27日),收割(10月15日)。
由图 1可知,3种栽培密度下的水稻分蘖数随水稻的生长呈先快速增长后缓慢降低的趋势。从水稻移植后到6月28日(分蘖期),3个处理组的水稻分蘖数变化趋势相似,均快速增加,其中,水稻分蘖数均为低密度>中密度>高密度,差异随水稻的生长逐渐变大,但处理组间差异未达到显著性水平(P>0.05)。在7月19日左右,各处理组的水稻每穴分蘖数均达到峰值,低、高、中密度每穴总分蘖数分别为12.33、10.65、8.28,低密度和高密度组的水稻分蘖数产生显著差异(P<0.05)。由7月19日以后,各处理组的水稻分蘖数略微下降且逐渐趋于稳定,低密度下的水稻分蘖数均显著高于高密度(P<0.05),尤其在9月19日左右(抽穗期)时二者差异达到极显著水平(P<0.01)。

|
图 1 不同水稻栽培密度下每穴水稻总分蘖数动态变化 Fig. 1 Dynamic changes of total tiller number of rice per hill at different planting densities |
植物净初级生产力的高低取决于植物自身生理生态特征与外界环境因子的相互作用,是评价生态系统功能结构特征的重要指标[17],而净初级生产力的表现形式也包括植物地上部分干物质质量,即生物量。水稻群体生物量是稻谷产量的基础和先决条件,水稻产量高低尤其取决于抽穗期后的光合生产能力,而干物质积累是提高光合生产力必要的群体结构物质基础[18]。图 2中,3种栽培密度下的水稻生物量在时间上的变化趋势相似,均表现为随水稻的生长而快速增加。返青期的各处理水稻生物量无显著差异(P>0.05);拔节期、抽穗期和成熟期水稻生物量积累由高至低均依次为低密度、中密度和高密度,低密度组的水稻生物量显著高于高密度组(P<0.05),且在抽穗期和成熟期时二者差异达到极显著水平(P<0.01)。拔节期和成熟期时,低密度栽培的水稻生物量显著高于中密度(P<0.05)。青田地区在7—9月光照和水温均达到水稻生长的理想条件,因此各处理组水稻生物量积累主要出现在拔节期-成熟期阶段。

|
图 2 不同发育时期的水稻生物量 Fig. 2 Rice biomass production in different development stages |
茎秆构成是组成水稻形态特征的重要因素[19],粗壮的茎秆是水稻物质积累和获得稻谷产量的保证[20]。提高光能利用率是获得水稻高产的关键[21],水稻茎秆构成在水稻的光能利用上具有重要意义[20, 22],而栽培密度的改变会影响水稻的茎秆构成和光合利用特征[23]。茎秆构成虽然是水稻品种的遗传特性,但生态条件也是重要的影响因素[24],保持合适的栽培密度有利于水稻发育成理想株型,对水稻提高产量和增加抗倒伏能力都有利[8, 20, 25]。实验结果(表 1)表明:低密度栽培条件下的水稻株高、秆长和秆基部外径均极显著高于中、高密度(P<0.01),而在中、高密度下均无显著差异(P>0.05);在水稻穗长表现方面,高密度组的穗长显著高于中密度组(P<0.05)、低密度组极显著高于中密度组(P<0.01),高、低密度组的穗长差异不显著(P>0.05);各处理组间的穗基部外径均无显著差异(P>0.05)。综合来看,低密度处理组与其他处理组相比(穗基部外径除外)具有显著的性状优势。

|
表 1 不同栽培密度下的水稻茎秆构成 Tab.1 Rice stem characters at different planting densities |
水稻产量主要由单位面积穗数、每穗粒数、结实率与千粒重等因素构成[26],水稻密度是影响水稻群体产量及其构成因素的重要因素[10]。于水稻收割前取各实验田块的水稻样品,分别计算水稻的产量构成相关指标(表 2)。低密度组的成穗率显著高于中、高密度组(P<0.05);有效穗数在3个处理组均表现出极显著差异(P<0.01),从大到小依次为:低密度>中密度>高密度。各处理组在其他产量构成指标中均未表现出显著性差异(P>0.05)。其中高密度组的水稻产量和千粒重略高于中、低密度组。

|
表 2 3种水稻栽培密度下水稻的产量构成 Tab.2 Rice yield composition at different planting densities |
各水稻茎秆构成因子和产量构成因子对水稻产量的重要性以关联度大小来表示,关联度大的表明该因子对水稻产量贡献大;反之,贡献小。根据表 3可知,水稻产量和各产量构成因子均有不同程度的关联性,灰色关联度总体分布在0.561~0.832之间。各水稻茎秆构成因子与水稻产量关联度由大到小排序依次为穗长(0.832)>秆基部外径(0.828)>株高(0.822)>秆长(0.820)>穗基部外径(0.812),其中与水稻产量关联度最大的茎秆构成因子是穗长,关联度最小的是穗基部外径;水稻产量构成因子与水稻产量关联度由大到小排序依次为千粒重(0.812)>成穗率(0.808)>结实率(0.807)>穗粒数(0.785)>生物量(0.672)>有效穗数(0.660)>每公顷穴数(0.561),其中与水稻产量关联度最大的产量构成因子是千粒重,关联度最小的是每公顷穴数。根据关联度大小可以较好地区分开水稻茎秆构成因子和产量构成因子各自与水稻产量的关联度,总体来讲,茎秆构成因子与水稻产量的关联度相对更加密切,为0.812~0.832。

|
表 3 关联度矩阵 Tab.3 Correlation matrix |
水质是稻-鱼共生系统中的动态指标,受物理、化学和生物等多重因素影响。水质情况影响着稻田水生环境和稻-鱼共生系统的生产力。稻-鱼共生系统中水体的pH、溶解氧(dissolved oxygen, DO)和温度在不同水稻密度下的变化如图 3所示。整体来看,随着水稻的生长,稻田水体的pH和DO降低、温度升高。拔节期后中密度的pH、DO和温度均是3个密度中最高的,但3个水体参数在不同密度组间差异均不显著(P>0.05)。

|
图 3 不同水稻生长时期稻田水体的pH(a)、溶解氧(b)和温度(c) Fig. 3 pH (a), dissolved oxygen (b) and temperature (c) of water in paddy field at different growth stages of rice |
田鱼的SR随水稻密度的降低呈略下降趋势,但各处理组间未达到显著差异水平(P>0.05)。中密度时田鱼的SGR达到最高,且极显著高于高密度组(P < 0.01)。见表 4。

|
表 4 3种水稻栽培密度稻田养殖田鱼的存活率和特定生长率 Tab.4 SR and SGR of fish at different planting densities |
水稻密度是栽培学中用于调节产量的一种有效手段[27]。在传统的青田稻-鱼共生系统中,当地农民常采用相对较低的水稻密度[1],这种较宽的间距有利于养殖田鱼的活动。研究测定了高、中、低3种不同水稻密度下的水稻生长及其产量构成, 结果表明,在每穴基本苗确定的条件下,栽插密度大,其单位面积的基本苗起点高,单穴土壤营养面积小,对水稻的生长发育会造成不利影响。因而在青田地区传统的稻-鱼共生模式中适当的降低栽插密度不会影响水稻产量。
在7月19日前,3种栽培模式下的水稻分蘖数均随时间快速增加(图 1),之后分蘖数略有下降是由分蘖后期无效分蘖的死亡所致。3个密度组的水稻从6月28日后先后开始进入无效分蘖期,分蘖期和拔节期两个时期出现重叠。实验表明,低密度组的水稻分蘖具有明显优势,这与一些研究结果[28-29]相似。在分蘖前期,水稻分蘖数受栽培密度的影响不大;而在分蘖后期,随栽培密度增加,每穴的分蘖和穗数有规律地减少,但单位面积总分蘖数和总穗数随密度增加而增加[8]。单位面积水稻穴数不同导致对土壤养分需求量有差异,加上不同栽培密度下水稻遮阴程度不同导致的光合生产力不同,不同密度下水稻生物量积累产生差异。在低密度条件下,单株水稻养分充足,叶片数增多,叶片长度和披垂角度也随之增加[30-31],水稻株间遮蔽较少、光照充足,水稻生物量积累的优势明显。在高密度栽培时,单位面积水稻株数较多,单株对土壤养分需求产生竞争,水稻间互相遮阴导致单株水稻光照减少,阻碍了水稻的生长发育及干物质积累。此外,水稻密植导致稻田通风条件较差,形成了阴暗潮湿的环境,适宜水稻害虫生存,加上水稻过密限制了田鱼的觅食等活动,不能充分摄食植株上的害虫和致病菌丝[2],更是增加了水稻感染病虫害的风险[11],进一步加剧了水稻营养的缺失。
水稻的理想株型要求茎秆构成适宜,其中:适当的株高以保证较高的生物学产量及中下部叶片较好的通风受光条件,保证叶片充足的伸展空间,增加抽穗后整株受光量;较大的秆基部外径保证植株坚韧、不易倒伏。在株高、秆长、穗长和秆基部外径方面,低密度组显著优于中、高密度组(表 1),表明稀植条件下水稻具有更为显著的性状优势,与之相比密植条件下的水稻个体间生存矛盾加剧,从而弱化了水稻的茎秆性状。本实验对不同水稻密度下水稻株高的测量结果与徐敏等[29]的研究结果相似,均表现为低密度条件下水稻株高性状优势明显,而与李静[8]在雅安开展的不同水稻密度的研究结果不同,可能是因为水稻品种和生长环境不同,本研究实验地点处于长江中下游生态区的青田,光照充足,雅安处于四川盆地的弱光区域。该研究中对西昌和雅安的研究结果不一样也侧面验证了这一点,本实验对水稻穗长的测定结果与该研究相似。栽培密度对水稻秆基部外径影响较大,杨世民等[32]研究认为,随栽插密度的增加,水稻茎秆基部节间变细长,茎壁变薄,垄鞘中淀粉、纤维素和木质素含量降低,充实度变差。
在水稻各产量构成因素上,栽培密度仅对成穗率和有效穗数产生显著影响,对水稻产量、千粒重、结实率和穗粒数并未产生显著影响(表 2)。有效穗数受水稻生长环境的影响较大,其变化也相对较大;而千粒重等指标一般与水稻种质有关,受外在环境影响较小,数值相对稳定[33]。水稻最高苗和成穗率是水稻有效穗的决定因素[8],本实验中栽培密度对水稻最高苗和成穗率产生显著影响。在稀植条件下,水稻光照和养分等环境资源充足,有利于水稻个体分蘖的发生,使分蘖成穗率明显提高、单株有效穗数增加[34]。XIE等[2]研究发现,青田田鱼有撞击水稻植株茎秆的行为,这一活动使水稻飞虱大量掉落水中,为鱼类取食。田鱼在早晨的撞击行为会摇落稻叶上的露滴,减少稻瘟病的孢子萌发及菌丝渗透。此外,田鱼可以破坏或直接吃掉水稻纹枯病的致病菌(立枯丝核菌)菌丝,一定程度上抑制了纹枯病的发生。水稻稀植使田鱼有了更多的活动空间,为田鱼的这种撞击行为提供了有利条件。本实验对水稻产量、千粒重和结实率的测定结果与徐敏等[29]的研究相似,均表现为在不同栽培密度下差异不显著。密植在水稻栽培学中被认为是一种稳产的手段[27],但在本实验中,栽培密度的不同并未对水稻产量产生显著影响,在密植条件下的水稻产量虽略高于在稀植条件下的水稻产量,但优势却并不明显,导致各处理组产量差异并不显著,可以结合水稻产量与茎秆构成因素、产量构成因素关系(表 3)推测原因如下:(1)茎秆构成与水稻产量的关联度更高(0.812~0.832),对水稻产量影响较大,而低密度栽培的水稻具有明显的茎秆构成性状优势,因此在稀植条件下的水稻产量不会显著低于密植条件下;(2)在产量构成因素中,对水稻产量贡献较大的是千粒重(0.812)、成穗率(0.808)和结实率(0.807),而千粒重和结实率在不同水稻密度下均没有显著差异,成穗率在低密度组显著高于中、高密度组,这也能保证在稀植条件下的水稻产量;(3)在产量构成因素中单位面积穴数与产量的关联度最低,这与白良明等[35]的研究结果相似,表明本实验中水稻密度对最终产量的影响不大;(4)此外,因为水稻植株间通风差、缺少光照等原因形成了阴暗潮湿的环境[7],加上活动空间受限、田鱼无法充分取食水稻植株上的害虫和破坏致病菌菌丝,高密度组的稻田于7、8月份高温天气出现局部水稻稻瘟病,且伴有不同程度的稻飞虱为害,这也进一步削弱了高密度组的水稻产出。本实验设置低密度处理的株行距为30 cm×40 cm,植株间距过大导致单位面积总穗数偏低,因此推测在本研究条件下限制低密度组获得高产量的主要因素是单位面积穗数[36],低密度栽培获得较多的每穴有效穗数和较高的成穗率并不能弥补单位面积总穗数减少造成的损失[9],从而使在稀植条件下并未像金传旭等[37]的实验一样得到理想的产量。青田稻-鱼共生系统在实际生产中,适当降低水稻密度可以在保证水稻产量的同时节约劳动成本。
鱼类的生长与环境的光照、水温、pH和DO等有关。在水稻密植的情况下:植株叶片阻碍了阳光的穿透,导致稻田水温偏低;较少的阳光穿透使浮游生物的光合作用变弱,导致稻田水体的DO较低[38],而较低的温度和溶氧均不利于鱼类的生长[39-40]。因此,在传统的稻-鱼共生模式中,青田当地农民经常采用较宽的水稻株行距,这种较宽的间距不仅可以使田鱼得到更多的活动空间,还可以使稻田中的环境因子(如水温和溶氧等)更适合鱼类的生存和生长。在水稻开始生长时,稻田水体的pH(8.40~8.58)和DO含量(3.55~5.54 mg/L)均处于较高水平(图 3),表明自养途径在稻田的水生环境中占主导地位[38];在拔节期后pH和DO含量的下降表明水生植物光合作用减弱,自养途径弱化。而限制稻田水生生物光合作用最重要的原因是水稻植株的遮阴,这尤其体现在密植稻田中[38]。随着水稻栽培密度的降低,株行距增加,单株水稻的叶片数量、长度和下垂角度均会增加,稻叶的遮阴区域也会相应增加。在水稻拔节期后,中密度栽培时,稻田水体的pH、DO和温度均略高于高、低密度,这可能是因为中密度栽培时水稻遮阴面积更少、稻叶不能完全封行。水稻栽培密度与水稻生长(包括叶片数、叶面积和地上部分生物量等)之间可能存在动态平衡[41],结合水稻生长情况和水稻间距来看,在同一阶段,高密度时水稻间距较小、低密度时水稻生长性能较好(由图 1、表 1中的水稻分蘖数、株高、秆长、穗长和秆基部外径等可知)均会阻碍阳光穿透,从而在稻田水面形成大面积的密封遮光,光照较少导致水温和水生光合作用下降。此外,吴敏芳等[13]发现,30 cm×30 m的水稻栽培密度配合较低的田鱼投放密度在一定程度上可以提高鱼类的饵料系数,充分利用稻-鱼共生系统中的各种资源。在本实验中,中密度栽培时田鱼SGR达到最高,且极显著高于高密度组(P < 0.01),表明中密度稻田由于阳光穿透而获得了适高的水温[42]、较强的光照[43]、较多的溶氧和较多的可以被鱼类摄食的浮游动植物[44],从而提高了田鱼的摄食水平[45-46],加快了鱼类的生长速率。此外,中、低密度组植株间距较大,田鱼较活跃,摄食水稻上的害虫等较多,也有利于田鱼的能量获得。
研究表明,适低的栽培密度可以使水稻获得显著的性状优势,青田稻-鱼共生系统采用30 cm×30 cm的栽培株行距可以有效提高田鱼的生长速率,且不会造成水稻减产。本实验为达到推广应用的目的而采用大田块设计,因此密度等控制因子的划分不够细致,后续研究可以考虑选用小区田块进行实验。通过对青田传统稻-鱼共生系统内在机理和哲学的不断探索和研究,我们可以更好地保护这个传统的农业方式并使其创造更大的价值。
| [1] |
焦雯珺. 全球重要农业文化遗产:浙江青田稻鱼共生系统[M]. 北京: 中国农业出版社, 2015. JIAO W J. Globally important agricultural heritage systems:the rice-fish coculture system of Qingtian, Zhejiang[M]. Beijing: The Agricultural Press of China, 2015. |
| [2] |
XIE J, HU L L, TANG J J, et al. Ecological mechanisms underlying the sustainability of the agricultural heritage rice-fish coculture system[J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(50): E1381-E1387. DOI:10.1073/pnas.1111043108 |
| [3] |
FULLER A M, MIN Q W, JIAO W J, et al. Globally important agricultural heritage systems (GIAHS) of China:the challenge of complexity in research[J]. Ecosystem Health and Sustainability, 2015, 1(2): 1-10. |
| [4] |
FREI M, KHAN M A M, RAZZAK M A, et al. Effects of a mixed culture of common carp, Cyprinus carpio L., and Nile tilapia, Oreochromis niloticus (L.), on terrestrial arthropod population, benthic fauna, and weed biomass in rice fields in Bangladesh[J]. Biological Control, 2007, 41(2): 207-213. |
| [5] |
ALTIERI M A. Linking ecologists and traditional farmers in the search for sustainable agriculture[J]. Frontiers in Ecology and the Environment, 2004, 2(1): 35-42. |
| [6] |
REN W Z, HU L L, GUO L, et al. Preservation of the genetic diversity of a local common carp in the agricultural heritage rice-fish system[J]. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(3): E546-E554. DOI:10.1073/pnas.1709582115 |
| [7] |
王寅, 徐建伟, 张丹丹, 等. 与栽培密度相关的水稻形态基因研究进展[J]. 核农学报, 2011, 25(5): 951-958. WANG Y, XU J W, ZHANG D D, et al. Research progress on plant morphological genes related to planting density in rice (Oryza sativa L.)[J]. Journal of Nuclear Agricultural Sciences, 2011, 25(5): 951-958. |
| [8] |
李静.生态条件和栽培密度对水稻群体特征、产量和品质的影响[D]. 成都: 四川农业大学, 2013. LI J. Effects of ecological conditions and transplanting density on the rice population characteristics, yield and quality[D]. Chengdu: Sichuan Agricultural University, 2013. http://cdmd.cnki.com.cn/Article/CDMD-10626-1014102122.htm |
| [9] |
HUANG M, YANG C L, JI Q M, et al. Tillering responses of rice to plant density and nitrogen rate in a subtropical environment of southern China[J]. Field Crops Research, 2013, 149: 187-192. DOI:10.1016/j.fcr.2013.04.029 |
| [10] |
戴正元, 李爱宏, 肖宁, 等. 超级杂交稻扬两优6号不同栽插密度下的群体生长特性[J]. 江苏农业学报, 2010, 26(1): 22-27. DAI Z Y, LI A H, XIAO N, et al. Population growth characteristics of super hybrid rice Yangliangyou 6 with different transplanting densities[J]. Jiangsu Journal of Agricultural Sciences, 2010, 26(1): 22-27. DOI:10.3969/j.issn.1000-4440.2010.01.005 |
| [11] |
苏祖芳, 霍中洋. 水稻合理密植研究进展[J]. 耕作与栽培, 2006(5): 6-9. SU Z F, HUO Z Y. Advances in research on rational close planting of rice[J]. Tillage and Cultivation, 2006(5): 6-9. DOI:10.3969/j.issn.1008-2239.2006.05.002 |
| [12] |
樊红柱, 曾祥忠, 吕世华. 水稻不同移栽密度的氮肥效应及氮素去向[J]. 核农学报, 2009, 23(4): 681-685. FAN H Z, ZENG X Z, LYU S H. Nitrogen utilization efficiency and fate of fertilizer under different transplanting density[J]. Journal of Nuclear Agricultural Sciences, 2009, 23(4): 681-685. |
| [13] |
吴敏芳, 张剑, 陈欣, 等. 提升稻鱼共生模式的若干关键技术研究[J]. 中国农学通报, 2014, 30(33): 51-55. WU M F, ZHANG J, CHEN X, et al. Practical technology for improving rice-fish co-culture system[J]. Chinese Agricultural Science Bulletin, 2014, 30(33): 51-55. |
| [14] |
ROTHUIS A J, VROMANT N, XUAN V T, et al. The effect of rice seeding rate on rice and fish production, and weed abundance in direct-seeded rice-fish culture[J]. Aquaculture, 1999, 172(3/4): 255-274. |
| [15] |
TSURUTA T, YAMAGUCHI M, ABE S I, et al. Effect of fish in rice-fish culture on the rice yield[J]. Fisheries Science, 2011, 77(1): 95-106. |
| [16] |
邓聚龙. 灰色系统基本方法[M]. 武汉: 华中理工大学出版社, 1988. DENG J L. Basic method of grey system[M]. Wuhan: Huazhong University of Science & Technology Press, 1988. |
| [17] |
王尚明, 张崇华, 胡逢喜, 等. 水稻地上部生物量及净初级生产力的定量分析[J]. 中国农学通报, 2008, 24(8): 201-205. WANG S M, ZHANG C H, HU F X, et al. Quantitative analysis on biomass and net primary productivity of aboveground parts in rice[J]. Chinese Agricultural Science Bulletin, 2008, 24(8): 201-205. |
| [18] |
凌启鸿, 张洪程, 蔡建中, 等. 水稻高产群体质量及其优化控制探讨[J]. 中国农业科学, 1993, 26(6): 1-11. LING Q H, ZHANG H C, CAI J Z, et al. Investigation on the population quality of high yield and its optimizing control programme in rice[J]. Scientia Agricultura Sinica, 1993, 26(6): 1-11. |
| [19] |
吕川根, 邹江石. 两个超级杂交稻与汕优63光合株型的比较分析[J]. 中国农业科学, 2003, 36(6): 633-639. Lü C G, ZOU J S. Comparative analysis on plant type of two super hybrid rice and Shanyou63[J]. Scientia Agricultura Sinica, 2003, 36(6): 633-639. DOI:10.3321/j.issn:0578-1752.2003.06.006 |
| [20] |
苏祖芳, 许乃霞, 孙成明, 等. 水稻抽穗后株型指标与产量形成关系的研究[J]. 中国农业科学, 2003, 36(1): 115-120. SU Z F, XU N X, SUN C M, et al. Study on the relationship between rice plant type indices after heading stage and yield formation[J]. Scientia Agricultura Sinica, 2003, 36(1): 115-120. DOI:10.3321/j.issn:0578-1752.2003.01.021 |
| [21] |
夏仲炎, 姜文武. 水稻群体结构与生产性能的研究[J]. 安徽农业大学学报, 1995, 22(s1): 39-42. XIA Z Y, JIANG W W. Rice population structure and the production performance study[J]. Journal of Anhui Agricultural University, 1995, 22(s1): 39-42. |
| [22] |
TIROL-PADRE A, LADHA J K, SINGH U, et al. Grain yield performance of rice genotypes at suboptimal levels of soil N as affected by N uptake and utilization efficiency[J]. Field Crops Research, 1996, 46(1/3): 127-143. |
| [23] |
张圣喜, 贺再新, 李涛, 等. 水稻中优281栽培密度与产量的相关性研究[J]. 湖南农业科学, 2009(8): 30-31, 33. ZHANG S X, HE Z X, LI T, et al. Study on the correlation of planting density with yield of rice Zhongyou 281[J]. Hunan Agricultural Sciences, 2009(8): 30-31, 33. DOI:10.3969/j.issn.1006-060X.2009.08.011 |
| [24] |
邹江石, 姚克敏, 吕川根, 等. 水稻两优培九株型特征研究[J]. 作物学报, 2003, 29(5): 652-657. ZOU J S, YAO K M, LYU C G, et al. Study on individual plant type character of Liangyoupeijiu rice[J]. Acta Agronomica Sinica, 2003, 29(5): 652-657. DOI:10.3321/j.issn:0496-3490.2003.05.003 |
| [25] |
周丽华, 吴厚雄, 刘辉, 等. 杂交水稻茎秆形态学优势性状与抗倒伏能力研究[J]. 种子, 2006, 25(6): 10-13. ZHOU L H, WU H X, LIU H, et al. Study on dominant morphological properties and lodging resistance of culm in hybrid rice[J]. Seed, 2006, 25(6): 10-13. |
| [26] |
杨滨娟, 黄国勤, 王超, 等. 稻田冬种绿肥对水稻产量和土壤肥力的影响[J]. 中国生态农业学报, 2013, 21(10): 1209-1216. YANG B J, HUANG G Q, WANG C, et al. Effects of winter green manure cultivation on rice yield and soil fertility in paddy field[J]. Chinese Journal of Eco-Agriculture, 2013, 21(10): 1209-1216. |
| [27] |
王成瑷, 王伯伦, 张文香, 等. 栽培密度对水稻产量及品质的影响[J]. 沈阳农业大学学报, 2004, 35(4): 318-322. WANG C A, WANG B L, ZHANG W X, et al. Effect of planting density on grain yield and quality of rice[J]. Journal of Shenyang Agricultural University, 2004, 35(4): 318-322. DOI:10.3969/j.issn.1000-1700.2004.04.006 |
| [28] |
钱银飞, 张洪程, 吴文革, 等. 移栽密度组合对机插水稻淮稻5号生长发育及产量形成的影响[J]. 江西农业大学学报, 2009, 31(1): 41-48. QIAN Y F, ZHANG H C, WU W G, et al. Effects of different transplanting density combinations on growth and yield of machine-transplanted rice Huaidao 5[J]. Acta Agriculturae Universitatis Jiangxiensis, 2009, 31(1): 41-48. DOI:10.3969/j.issn.1000-2286.2009.01.010 |
| [29] |
徐敏, 马旭洲, 王武. 稻蟹共生系统水稻栽培模式对水稻和河蟹的影响[J]. 中国农业科学, 2014, 47(9): 1828-1835. XU M, MA X Z, WANG W. Effects of different cultivation patterns on rice yield and crab in rice-crab culture system[J]. Scientia Agricultura Sinica, 2014, 47(9): 1828-1835. DOI:10.3864/j.issn.0578-1752.2014.09.018 |
| [30] |
林洪鑫, 潘晓华, 石庆华, 等. 栽插密度与施氮量对双季稻上部三叶叶长和叶角的影响[J]. 作物学报, 2010, 36(10): 1743-1751. LIN H X, PAN X H, SHI Q H, et al. Effects of nitrogen application amount and planting density on angle and length of top three leaves in double-cropping rice[J]. Acta Agronomica Sinica, 2010, 36(10): 1743-1751. DOI:10.3969/j.issn.1000-2561.2010.10.017 |
| [31] |
刘武, 谢明德, 黄林, 等. 氮肥用量和移栽密度对超级早稻干物质积累及叶蘖生长的影响[J]. 作物研究, 2008, 22(4): 243-248. LIU W, XIE M D, HUANG L, et al. Effects of transplanting density and nitrogen rate on dry matter accumulation and growth of leaves and tillers in super early rice[J]. Crop Research, 2008, 22(4): 243-248. DOI:10.3969/j.issn.1001-5280.2008.04.009 |
| [32] |
杨世民, 谢力, 郑顺林, 等. 氮肥水平和栽插密度对杂交稻茎秆理化特性与抗倒伏性的影响[J]. 作物学报, 2009, 35(1): 93-103. YANG S M, XIE L, ZHENG S L, et al. Effects of nitrogen rate and transplanting density on physical and chemical characteristics and lodging resistance of culms in hybrid rice[J]. Acta Agronomica Sinica, 2009, 35(1): 93-103. |
| [33] |
姜廷波, 李荣田, 崔成焕, 等. 水稻穗型构成性状的相关与通径分析[J]. 东北农业大学学报, 1995, 26(4): 330-335. JIANG T B, LI R T, CUI C H, et al. Correletion and path analysis between characters of rice panicle structure[J]. Journal of Northeast Agricultural University, 1995, 26(4): 330-335. |
| [34] |
陈惠哲, 朱德峰, 林贤青, 等. 稀植条件下杂交稻分蘖成穗规律和穗粒结构研究[J]. 杂交水稻, 2004, 19(6): 51-54. CHEN H Z, ZHU D F, LIN X Q, et al. Studies on the tillering dynamics, panicle formation and composition of panicles of hybrid rice under sparse transplanting density[J]. Hybrid Rice, 2004, 19(6): 51-54. DOI:10.3969/j.issn.1005-3956.2004.06.022 |
| [35] |
白良明, 张凤鸣. 水稻产量及构成因素的灰色关联度分析初探[J]. 农业系统科学与综合研究, 2001, 17(1): 65-66. BAI L M, ZHANG F M. Grey correlation analysis of yield and yield components in rice[J]. System Sciences and Comprehensive Studies in Agriculture, 2001, 17(1): 65-66. DOI:10.3969/j.issn.1001-0068.2001.01.021 |
| [36] |
袁奇, 于林惠, 石世杰, 等. 机插秧每穴栽插苗数对水稻分蘖与成穗的影响[J]. 农业工程学报, 2007, 23(10): 121-125. YUAN Q, YU L H, SHI S J, et al. Effects of different quantities of planting seedlings per hill on outgrowth and tiller production for machine-transplanted rice[J]. Transactions of the CSAE, 2007, 23(10): 121-125. DOI:10.3321/j.issn:1002-6819.2007.10.021 |
| [37] |
金传旭, 钟芹辅, 黄大英, 等. 栽插密度与穴栽苗数对水稻产量及其构成因素的影响[J]. 贵州农业科学, 2012, 40(4): 85-87, 90. JIN C X, ZHONG Q F, HUANG D Y, et al. Effects of transplanting density and seedling number per hole on yield and yield composition of rice[J]. Guizhou Agricultural Sciences, 2012, 40(4): 85-87, 90. DOI:10.3969/j.issn.1001-3601.2012.04.024 |
| [38] |
CHAKRABORTY B K, MIRZA M J A. Growth and yield of fish under high-density stocking and phased harvesting in rice-fish system[J]. Journal of Crop and Weed, 2009, 5(1): 1-10. |
| [39] |
NURDIANI R, ZENG C S. Effects of temperature and salinity on the survival and development of mud crab, Scylla serrata (Forsskål), larvae[J]. Aquaculture Research, 2007, 38(14): 1529-1538. DOI:10.1111/j.1365-2109.2007.01810.x |
| [40] |
ZHANG W, CAO Z D, PENG J L, et al. The effects of dissolved oxygen level on the metabolic interaction between digestion and locomotion in juvenile southern catfish (Silurus meridionalis Chen)[J]. Comparative Biochemistry and Physiology Part A:Molecular & Integrative Physiology, 2010, 157(3): 212-219. |
| [41] |
CHAUHAN B S, ABUGHO S B. Effects of water regime, nitrogen fertilization, and rice plant density on growth and reproduction of lowland weed Echinochloa crus galli[J]. Crop Protection, 2013, 54: 142-147. DOI:10.1016/j.cropro.2013.08.005 |
| [42] |
KILAMBI R V, ROBISON W R. Effects of temperature and stocking density on food consumption and growth of grass carp Ctenopharyngodon idella, Val[J]. Journal of Fish Biology, 1979, 15(3): 337-342. DOI:10.1111/j.1095-8649.1979.tb03614.x |
| [43] |
李大鹏, 庄平, 严安生, 等. 光照、水流和养殖密度对史氏鲟稚鱼摄食、行为和生长的影响[J]. 水产学报, 2004, 28(1): 54-61. LI D P, ZHUANG P, YAN A S, et al. The influences of illumination, water current and stocking density on feeding, behavior and growth in juveniles Acipenser schrenckii[J]. Journal of Fisheries of China, 2004, 28(1): 54-61. |
| [44] |
VROMANT N, CHAU N T H. Overall effect of rice biomass and fish on the aquatic ecology of experimental rice plots[J]. Agriculture, Ecosystems & Environment, 2005, 111(1/4): 153-165. |
| [45] |
周显青, 牛翠娟, 李庆芬, 等. 光照强度对中华鳖稚鳖摄食和生长的影响[J]. 动物学报, 1998, 44(2): 157-161. ZHOU X Q, NIU C J, LI Q F, et al. The effects of light intensity on daily food consumption and specific growth rate of the juvenile soft-shelled turtle, Trionyx sinensis[J]. Acta Zoologica Sinica, 1998, 44(2): 157-161. |
| [46] |
STEINBACHER P, MARSCHALLINGER J, OBERMAYER A, et al. Temperature-dependent modification of muscle precursor cell behaviour is an underlying reason for lasting effects on muscle cellularity and body growth of teleost fish[J]. Journal of Experimental Biology, 2011, 214(11): 1791-1801. DOI:10.1242/jeb.050096 |
2. Centre for Research on Environmental Ecology and Fish Nutrition, Ministry of Agriculture and Rural Affairs, Shanghai Ocean University, Shanghai 201306, China;
3. Key Laboratory of Freshwater Aquatic Genetic Resources, Ministry of Agriculture and Rural Affairs, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China;
4. Qingtian Yugong Agricultural Technology Co. Ltd., Qingtian 323900, Zhejiang, China