 2019,
Vol. 28
2019,
Vol. 28



2. 国家远洋渔业工程技术研究中心, 上海 201306;
3. 大洋渔业资源可持续开发教育部重点实验室, 上海 201306;
4. 农业农村部大洋渔业开发重点实验室, 上海 201306
太平洋褶柔鱼(Todarodes pacificus)是世界上最早被大规模开发利用的头足类种类之一,广泛分布于西北太平洋20°N ~60°N海域[1]。其富有丰富的蛋白质和微量元素[2]以及较高的可食比重[3]而成为我国主要的鱿钓捕捞对象。太平洋褶柔鱼一年四季均有产卵[4],根据产卵高峰期和迁徙模式的不同可分为夏生群、秋生群和冬生群[5],且以秋生群和冬生群为主[6]。1970年以前,冬生群产量较高,之后冬生群资源衰退,秋生群成为主要捕捞对象[7]。秋生群主要渔获来自日本海,其雄性大约在9月成熟,雌性10月以后开始成熟,主要于10—12月在日本北陆沿岸的对马海峡海域附近的东海进行产卵,新生群体于次年5—9月洄游回育幼场[8]。
环境对短生命周期生物的影响明显,这一影响可能超过捕捞影响[9]。不同生长阶段的太平洋褶柔鱼对环境的适应能力不同,以日本列岛海域情况为例,其北上群体的索饵适温范围为10~17 ℃,成体交配适温为13~18 ℃,而南下群体产卵的适温范围为15~20 ℃[10]。产卵场表层水温在一定程度上会影响太平洋褶柔鱼补充量的变化[11];海表面温度对单位捕捞努力量渔获量(catch per unit effort, CPUE)也具有显著影响[12]。此外,陆地河流、降水[13]、叶绿素浓度[14]等环境因素的变化也会对短生命周期生物产生一定影响。
自1980年头足类与环境关系被CADYY[15]和ROWELL等[16]提出后,研究者们就开始不断地探索资源与环境之间的关系,并尝试用环境变量解释其资源变动,到目前为止,用鱿鱼与环境的关系来预测柔鱼资源仍较为困难[17]。有关鱿鱼资源评估,目前对衰减模型的研究相对较深,对其他模型的研究尚属探索阶段[18]。本文选取日本海海域平均海表面温度(SST)作为环境影响因子,用剩余产量模型(Schaefer模型和Fox模型)对太平洋褶柔鱼渔获量进行预测,以期为太平洋褶柔鱼的可持续利用和管理提供依据。
1 材料与方法 1.1 数据选择渔业数据来自日本海海洋渔业研究所《平成29(2017)年太平洋褶柔鱼秋生群落资源评估报告》,包括:1985—2014年的总渔获量、捕捞努力量及CPUE[8](表 1)。捕捞时间为5—9月。

|
表 1 1985—2014年日本海太平洋褶柔鱼秋生群渔业数据以及5—9月平均海表面温度 Tab.1 Fishery data of the autumn cohort of Todarodes pacificus in Japan Sea and the average sea surface temperature from May to September in 1985-2014 |
日本太平洋褶柔鱼渔业开发始于20世纪60年代后半期,70年代中期资源出现下降,90年代又开始恢复,2003年后又出现下降[8]。韩国的太平洋褶柔鱼渔业在1990年之后迅速发展,1993年之后开始波动下降,1993—2003年波动上升,2003年之后再次波动下降[8]。因此根据日韩发展情况,本文以1993和2003年为界对1985—2014年的总渔获量进行分段分析,分别为:1985—1993年(相对稳定段)、1994—2002年(相对波动段)和2003—2014年(相对稳定段)。
温度数据来自OceanWatch(http://oceanwatch.pifsc.noaa.gov/las/servlets/dataset)(表 1)。因36°N ~39°N,130°E~138°E海域的海表面温度对太平洋褶柔鱼CPUE具有显著影响[12]且秋生群一般在5—9月洄游回育幼场,因而本文选取36°N ~39°N,130°E ~138°E海域内1985—2014年5—9月的月平均海表面温度(sea surface temperature, SST)进行分析(表 1)。
1.2 剩余产量模型由Schaefer模型和Fox模型得出渔获量、捕捞努力量和CPUE的关系式如下:
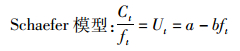 (1)
(1)
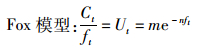 (2)
(2)
式中:Ct为t年渔获量观测值,t;ft为t年捕捞努力量,d;Ut为t年的单位捕捞努力量渔获量,t/d;a,b,m,n为参数。
根据Ct、ft和式(1)、(2)求出相应参数a、b、m和n, 得到如下公式:
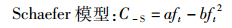 (3)
(3)
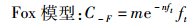 (4)
(4)
式中:C-S为t年Schaefer模型求得的渔获量预测值;C-F为t年Fox模型的渔获量预测值。
1.3 基于环境因子的剩余产量模型假设渔获量观测值与预测值之间的差值与捕捞努力量的比值ΔU(CPUE残差)是由SST引起[19]且呈线性关系,则:
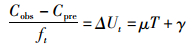 (5)
(5)
式中:Cobs为t年渔获量观测值;Cpre为t年渔获量预测值;μ、γ为常量;T表示的是海表面温度SST。
由式(3)(5)或式(4)(5)可以得到基于环境因子的渔获量预测值公式见表 2。

|
表 2 1985—2014年基于SST的日本海太平洋褶柔鱼秋生群Schaefer模型和Fox模型 Tab.2 Schaefer model and Fox model based on SST for the autumn cohort of Todarodes pacificus in Japan Sea from 1985 to 2014 |
模型拟合效果用和方差(sum of squared errors,SSE)、均方差(mean squared error,MSE)、均方根(root mean square error,RMSE)、确定系数R2和赤池信息量(akaike information criterion,AIC)来比较。
(1) 和方差SSE:又称残差平方和,是拟合数据和原始数据对应点的误差的平方和。计算公式如下:
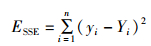 (8)
(8)
式中:yi指的是原始数据,Yi指的是预测数据。
(2) 均方差MSE:又称方差,是预测数据和原始数据对应点误差的平方和的均值,也就是SSE/n。计算公式如下:
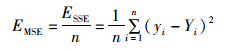 (9)
(9)
式中:n指的预测数据数或原始数据数。
(3) 均方根RMSE:也叫回归系统的拟合标准差,是MSE的平方根。计算公式如下:
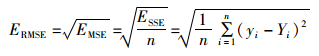 (10)
(10)
(4) 确定系数R2计算公式如下:
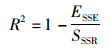 (11)
(11)
式中:SSSR为离差平方和(total sum of squared residual,SSR),为原始数据和其均值之差的平方和。公式如下:
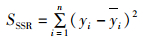 (12)
(12)
式中:yi为原始数据的均值。
(5) AIC:Akaike information criterion,赤池信息量。公式如下[20]:
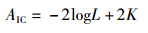 (13)
(13)
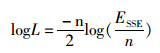 (14)
(14)
式中:K为模型中的参数个数;logL为对数似然值。
SSE、MSE、RMSE和AIC值越小,表明模型的拟合效果更好。R2的正常取值范围为[0, 1],越接近1,表明预测数据对原始数据的解释能力越强,这个模型对数据拟合得也较好。
2 结果与分析 2.1 渔获量与平均海表面温度渔获量1985—1993年增长较快;1993—2003年呈波动状态,且起伏较大;2003—2014年下降较快。SST 1985—1993年呈波动下降趋势;1993—2003年整体呈波动状态,且尤以1993—1995、2002—2003年起伏较大;2003—2014年呈波动上升趋势。
渔获量与温度极显著相关(P<0.01)且2003—2014年呈极显著负相关(P<0.01)。渔获量整体在1985—2014年随温度升高而降低,随温度降低而升高(图 1)。

|
图 1 1985—2014年日本海太平洋褶柔鱼秋生群的总渔获量及5—9月平均海表面温度(SST) Fig. 1 Total catch of the autumn cohort of T.pacificus in Japan Sea and the average sea surface temperature (SST) from May to September from 1985 to 2014 |
经回归计算,得Schaefer模型(P<0.01,R2=0.59)和Fox模型(P<0.01,R2=0.68)的参数见表 3。

|
表 3 1985—2014年日本海太平洋褶柔鱼秋生群Schaefer模型和Fox模型的参数 Tab.3 Parameter of Schaefer model and Fox model based on dates of the autumn cohort of T.pacificus in Japan Sea from 1985 to 2014 |
Schaefer模型和Fox模型在1985—1993年和2003—2014年对观测值的拟合相对较好,在1993—2003年的拟合较差。两个模型在1985—1993年的预测值呈波动上升,2003—2014呈波动下降,1993—2003年波动起伏。两个模型在相对稳定段(1985—1993年和2003—2014年)的预测值整体高于观测值,在波动段(1993—2003年)的预测值小于观测值且相差较大,Fox模型整体波动趋于平缓,Schaefer模型波动相对较大。见图 2。

|
图 2 1985—2014年基于日本海太平洋褶柔鱼秋生群渔业数据的Schaefer和Fox模型的预测值与观测值 Fig. 2 The predicted and observed catch of Schaefer and Fox model based on the fishery data of the autumn cohort of T.pacificus in Japan Sea from 1985 to 2014 |
Schaefer模型和Fox模型公式为
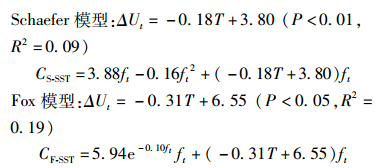
ΔU和SST整体呈负线性相关关系(P<0.01),且分段效果较为明显。见图 3。

|
图 3 1985—2014年基于日本海太平洋褶柔鱼秋生群渔业数据的剩余产量模型的5—9月平均海表面温度(SST)和ΔU关系 Fig. 3 Relationship between average sea surface temperature (SST)from May to September and ΔU in the surplus yield model based on the fishery data of the autumn cohort of T.pacificus in Japan Sea from 1985 to 2014 |
不分段的基于SST的两个模型在相对稳定段的预测值与观测值较为接近,尤以2003—2014年段明显,在相对波动段的拟合相对较差,见图 4。

|
图 4 1985—2014年基于5—9月平均海表面温度SST的日本海太平洋褶柔鱼秋生群不分段剩余产量模型的预测值与观测值比较图 Fig. 4 Comparison of predicted and observed values of the not segmented surplus yield model of the autumn cohort of T.pacificus in Japan Sea based on average sea surface temperature (SST)from May to September from 1985 to 2014 |
本文以1993和2003年为界对1985—2014年的情况进行分段分析,分别为1985—1993年(相对稳定段)、1994—2002年(相对波动段)和2003—2014年(相对稳定段)。见表 4。

|
表 4 1985—2014年基于5—9月平海表温SST的日本海太平洋褶柔鱼秋生群的Schaefer模型和Fox模型 Tab.4 Schaefer model and Fox model based on average sea surface temperature SST from May to September of the autumn cohort of T.pacificus in Japan Sea from 1985 to 2014 |
分段基于SST的两个模型的预测值跟观测值都较为接近。分段Schaefer model-SST在相对稳定段的预测值整体高于观测值;分段Fox model-SST在1985—1993预测值整体低于观测值,2007年后的预测值整体高于观测值:两个模型在相对波动段的拟合相对都不是很好,见图 5。

|
图 5 1985—2014年基于5—9月平均海表面温度SST的日本海太平洋褶柔鱼秋生群剩余产量模型的预测值与观测值比较图 Fig. 5 Comparison of predicted and observed values of the surplus yield model in the Japan sea the autumn cohort of T.pacificus based on average sea surface temperature (SST)from May to September from 1985 to 2014 |
拟合效果为分段Schaefer model-SST最好。不基于AIC的分析结果表明拟合效果依次是分段Foxmodel-SST、Schaefer model、不分段Schaefer model-SST、Fox model,最后是不分段的Foxmodel-SST;基于AIC的分析结果表明拟合效果随参数增多而变差,无论参数变量为多少,Schaefer模型的拟合效果均好于Fox模型的拟合效果。见表 5。

|
表 5 1985—2014年日本海太平洋褶柔鱼秋生群六种模型误差参数比较 Tab.5 Comparison of error parameters of six models of the autumn cohort of T. pacificus in Japan Sea from 1985 to 2014 |
短生命周期的生物生长与环境密切相关,目前太平洋褶柔鱼的相关环境影响因子涉及温度、盐度、叶绿素浓度等。关于温度的研究基本贯穿其生长发育过程(以日本列岛海域为例):索饵适温(10~17 ℃)、交配适温(13~18 ℃)、产卵适温(15~20 ℃)、渔获适温(10~17 ℃,尤以12~16 ℃渔获效果更佳)、胚胎适宜温度(具高温依赖性,12.5 ℃以下会发育失败)[7, 21-25],胚胎正常发育适温14~26 ℃、胚胎发育最高存活率适温14.7~22.2 ℃[26],幼仔适宜温度17~23 ℃[27]。不同的性别对温度的适应能力也不同,雄性一般在13 ℃成熟,雌性在15 ℃成熟,性腺的成熟会随着温度的升高而加快[7, 10]。不同地区不同季节的渔获适温范围不同,例如秋季的九州西北海域在表温为12~22 ℃渔获较多,冬季的九州海域、东海北部和中部在表温为12~22 ℃的渔获较多[7]。
本文渔场研究范围在日本海,主要位于新宫千臣等研究的太平洋褶柔鱼作业渔场的渔场2、渔场4和小部分的渔场5[7],位于日本列岛周围海域内,主要渔期在5—9月,温度研究范围为18~23 ℃。渔获量与SST呈极显著相关,且尤以2003—2014年呈极显著负相关,这与其索饵适温范围为10~17 ℃相一致并与日本列岛海域渔获适温10~17 ℃的结论相一致。整体渔获量在18~23 ℃范围之内会随着温度的升高而降低。
3.2 六种模型比较资源的周期波动主要与环境、种群密度、捕捞等密切相关。环境变化会导致短生命周期生物资源量与渔获量剧烈波动,这一观念目前已被国际普遍认可[28-29]。此外,头足类在生态系统中又占据着重要的承上启下作用[30],因而探究头足类资源与环境的关系变得极为重要。目前,关于短生命周期资源评估的研究相对较深的只有衰减模型,而衰减模型本身又存在很多问题,比如未考虑环境因子影响、对自然死亡率相对敏感等[31]。因而研究一种更为合理的短生命周期物种资源评估模型已成为未来的一种趋势。与此同时,剩余产量模型在被广泛应用的同时又有着未考虑环境因子的缺点,因而基于环境因子的剩余产量模型不失为未来的一种可行性的资源评估方法。
本文选择36°N ~39°N,130°E ~138°E海域5—9月的平均海表面温度SST作为环境因子,做出了不基于SST的两种剩余产量模型和基于SST的4种剩余产量模型(分段和不分段),其中分段Schaefer model-SST的拟合程度最好。这一结果表明,在研究年份较多的情况下,模型加入环境因子的过程中应该根据研究对象的不同年段的整体情况不同而进行分段分析,这样不仅可以更为贴切的表现数据之间的关系,而且可以提高拟合效果,而未考虑相关情况时不建议将环境因子引入到相关模型中,因为可能会造成较差的拟合效果。这一结果与李纲等[19]的观点较为不一致,可能因为研究鱼种不同,也可能因为时间段的设置问题。
此外,未分段且加入SST和未加入SST的4种剩余产量模型在相对稳定段的预测值整体高于观测值,表明剩余产量模型在未加入环境因子或者加入环境因子设置较为不合理的情况下,结果会有偏高的趋势。加入SST的4种剩余产量模型在2003—2014年的拟合程度相对更好。6种模型对相对波动段的拟合程度相较而言都不是很好,对相对稳定段的拟合程度相对较好,这一现象可能因为:在相对稳定段,不论渔获量处于上升趋势(1985—1993年)还是下降趋势(2003—2014年),整体捕捞努力量水平都趋于稳定,也即捕捞对该资源的影响趋于稳定,因而环境的影响相对较明显;而在波动段,捕捞努力量处于不稳定波动增长状态,且1994—2002年的资源量也处于快速增长状态[8],观测渔获量值与捕捞努力量和环境之间的关系较难预测,所以导致结果较为不理想。
3.3 模型的应用因2003—2014年的渔获量与SST之间呈极显著负相关(P<0.01),且分段Schaefer model-SST的拟合程度最好,所以本文选择2003—2014年的分段Schaefer model-SST进行计算相应的最大持续产量(maximum sustainable yield,MSY)和最大持续产量对应的捕捞努力量fMSY,其结果为
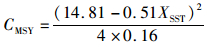 (15)
(15)
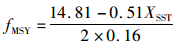 (16)
(16)
式中:CMSY为最大持续产量值MSY;CMSY和fMSY会随着SST的变化而进行变化,因而可以为相关渔业管理部门调整渔业管理措施提供一定的借鉴。
3.4 模型的优点和不足本研究基于有限数据所建立的模型,较好地描述了日本海太平洋褶柔鱼秋生群相对稳定段(也即捕捞努力量相对稳定段)渔获量、捕捞努力量与SST之间的关系。未来在各国状况不会发生较大变化的情况下(也即捕捞努力量不会进行不稳定起伏波动的情况下),2003—2014年的分段Schaefer model-SST可被用于相关预测,从而对于产品市场进行相应调控。
对于太平洋褶柔鱼渔业预测,本文尚未考虑其他环境因素对结果带来的影响,也没有从最大经济效益等方面考虑问题,所建议的在渔业模型建立过程中应该根据不同阶段情况进行不同分析,可能会增加相应计算量及复杂程度。虽然模型在假设前提下较为合理地模拟出了渔获量,但限于数据量,未对更多阶段的状况进行分析从而归纳总结出最适宜的模型,因此,期望在后续的数据更新中能进一步突破相关内容。
| [1] |
KIM J J, STOCKHAUSEN W, KIM S, et al. Understanding interannual variability in the distribution of, and transport processes affecting, the early life stages of Todarodes pacificus using behavioral-hydrodynamic modeling approaches[J]. Oceanography, 2015, 138: 571-583. |
| [2] |
郝振林, 唐雪娇, 宋坚, 等. 太平洋褶柔鱼胆固醇与牛磺酸含量分析[J]. 河北渔业, 2013(7): 10-11. HAO Z L, TANG X J, SONG J, et al. Analysis of cholesterol and taurine content in Todarodes pacificus[J]. Hebei Fisheries, 2013(7): 10-11. DOI:10.3969/j.issn.1004-6755.2013.07.004 |
| [3] |
曲映红, 陈新军, 刘志东, 等. 太平洋西北海域柔鱼理化特性的初步分析[J]. 食品工业, 2014, 35(10): 191-193. QU Y H, CHEN X J, LIU Z D, et al. Primary analysis on physical and chemical properties of Ommastrephes bartramii in the northwestern pacific ocean[J]. TheFood Industry, 2014, 35(10): 191-193. |
| [4] |
杨林林, 姜亚洲, 程家骅. 东海太平洋褶柔鱼生殖群体的空间分布及其与环境因子的关系[J]. 生态学报, 2010, 30(7): 1825-1833. YANG L L, JIANG Y Z, CHENG J H. Spatial distribution of mature stock of Pacific squid, Todarodes pacificus and its relation to environmental factors in the East China Sea[J]. ActaEcologicaSinica, 2010, 30(7): 1825-1833. |
| [5] |
OKUTANI T, WATANABE T. Stock assessment by larval surveys of the winter population of Todarodes pacificus steenstrup (Cephalopoda:Ommastrephidae), with a review of early works[J]. Biological Oceanography, 1983, 2(2/4): 401-431. |
| [6] |
MURATA M. Population assessment, management and fishery forecasting for the Japanese common squid, Todarodes pacificus[M]//CADDY JF. Marine Invertebrate Fisheries: Their Assessment and Management. New York, USA: John Wiley and Sons, 1989: 613-636.
|
| [7] |
王尧耕, 陈新军. 世界大洋性经济柔鱼类资源及其渔业[M]. 北京: 海洋出版社, 2005: 160-188. WANG Y G, CHEN X J. World oceanic economic squid resources and fisheries[M]. Beijing: China Ocean Press, 2005: 160-188. |
| [8] |
加賀敏樹, 山下紀生, 岡本俊.平成29(2017)年度スルメイカ秋季発生系群の資源評価[R].北海道: 北海道区水産研究所, 2017. KAGA T, YAMASHITA N, OKAMOTO S. Evaluation of the resources of the autumn cohort of Todarodes pacificus in 2017[R].Hokkaido: Hokkaido Fisheries Research Institute, 2017. |
| [9] |
KING M. Fisheries biology, assessment and management[M]. Oxford, UK: Fishing News Books Press, 1995: 192-197.
|
| [10] |
董正之. 世界大洋经济头足类生物学[M]. 济南: 山东科学技术出版社, 1991: 100-118. DONG Z Z. Biology of the economic species of cephalopods in the world oceans[M]. Jinan: Shandong Science and Technology Press, 1991: 100-118. |
| [11] |
SAKURAI Y, KIYOFUJI H, SAITOH S, et al. Changes in inferred spawning areas of Todarodes pacificus (Cephalopoda:Ommastrephidae) due to changing environmental conditions[J]. ICES Journal of Marine Science, 2000, 57(1): 24-30. DOI:10.1006/jmsc.2000.0667 |
| [12] |
唐峰华, 史赟荣, 朱金鑫, 等. 海洋环境因子对日本海太平洋褶柔鱼渔场时空分布的影响[J]. 中国水产科学, 2015, 22(5): 1036-1043. TANG F H, SHI Y R, ZHU J X, et al. Influence of marine environment factors on temporal and spatial dis-tribution of Japanese common squid fishing grounds in the Sea of Japan[J]. Journal of Fishery Sciences of China, 2015, 22(5): 1036-1043. |
| [13] |
BAKUN A, CSIRKE J. Environmental processes and recruitment variability[M]//RODHOUSE P G, DAWE E G, O'DORR K. Squid Recruitment Dynamics. The Genus Illex as a Model, The Commercial Illex Species andInfluences on Variability. Rome: FAO, 1998: 273.
|
| [14] |
ICHII T, MAHAPATRA K, SAKAI M, et al. Life history of the neon flying squid:effect of the oceanographic regime in the North Pacific Ocean[J]. Marine Ecology Progress Series, 2009, 378: 1-11. DOI:10.3354/meps07873 |
| [15] |
CADDY J F. The cephalopods: factors relevant to their population dynamics and to the assessment and management of stocks[M]//CADDYJ F. Advances in Assessment of World Cephalopod Resources. Rome: FAO, 1983: 416-449.
|
| [16] |
ROWELL T W, YOUNG J H, POULARD J C, et al. Changes in the distribution and biological characteristics of Illex illecebrosus on the Scotian shelf, 1980-83[J]. NAFO Scientific Council Studies, 1985, 9: 11-26. |
| [17] |
PIERCE G J, BOYLE P R. Empirical modelling of interannual trends in abundance of squid (Loligoforbesi) in Scottish waters[J]. Fisheries Research, 2003, 59(3): 305-326. DOI:10.1016/S0165-7836(02)00028-0 |
| [18] |
陈新军, 曹杰, 田思泉, 等. 鱿鱼类资源评估与管理研究现状[J]. 上海海洋大学学报, 2009, 18(4): 495-501. CHEN X J, CAO J, TIAN S Q, et al. Review on stock assessment and management of the squids[J]. Journal of Shanghai Ocean University, 2009, 18(4): 495-501. |
| [19] |
李纲, 郑晓琼, 朱国平, 等. 基于水温因子的东、黄海鲐鱼剩余产量模型建立[J]. 上海海洋大学学报, 2011, 20(1): 108-113. LI G, ZHENG X Q, ZHU G P, et al. Establishment of surplus production model for chub mackerel (Scomber japonicus) in the East China Sea and Yellow Sea based on the sea surfacetemperature[J]. Journal of Shanghai Ocean University, 2011, 20(1): 108-113. |
| [20] |
BURNHAM K P, ANDERSON D R, HUYVAERT K P. AIC model selection and multimodel inference in behavioral ecology:some background, observations, and comparisons[J]. Behavioral Ecology and Sociobiology, 2011, 65(1): 23-35. DOI:10.1007/s00265-010-1029-6 |
| [21] |
BOYLEP R. Cephalopod life cycles[M]. London: Academic Press, 1987: 5-31.
|
| [22] |
O'DOR R K, BALCH N, FOY E A, et al. Embryonic development of the squid, Illexillecebrosus, and effect of temperature on development rates[J]. Journal of Northwest Atlantic Fisheries Science, 1982, 3: 41-45. DOI:10.2960/J.v3.a3 |
| [23] |
WATANABE K, SAKURAI Y, SEGAWA S, et al. Development of the ommastrephid squid Todarode spacificus from fertilized egg to the rhynchoteuthion paralarva[J]. American Malacological Bulletin, 1996, 13: 73-88. |
| [24] |
山本潤, 宮長幸, 福井信一, 等. スルメイカふ化幼生の遊泳行動に対する水温の影響[J]. 水産海洋研究, 2012, 76(1): 18-23. YAMAMOTO J, MIYANAGA S, FUKUI S, et al. Effect of temperature on swimming behavior of paralarvae of the Japanese common squid Todarodes pacificus[J]. Fisheries Ocean Research, 2012, 76(1): 18-23. |
| [25] |
董正之. 中国动物志软体动物门(头足纲)[M]. 北京: 科学出版社, 1988: 81-84. DONG Z Z. Chinese zoology mollusc (Cephalopoda)[M]. Beijing: Science Press, 1988: 81-84. |
| [26] |
SAKURAI Y, BOWER J R, NAKAMURA Y, et al. Effects of temperature on development and survival of Todarodes pacificus embryos and paralarvae[J]. American Malacological Bulletin, 1996, 13(1/2): 89-95. |
| [27] |
BOWER J R. A biological study egg masses and paralarvae of the squid Todarodes pacificus[D]. Hakodate, Japan: Hokkaido University, 1997.
|
| [28] |
YATSU A, WATANABE T, MORI J, et al. Interannual variability in stock abundance of the neon flying squid, Ommastrephes bartramii, in the north pacific ocean during 1979-1998:impact of driftnet fishing and oceanographic conditions[J]. Fisheries Oceanography, 2000, 9(2): 163-170. DOI:10.1046/j.1365-2419.2000.00130.x |
| [29] |
RODHOUSEP G. Managing and forecasting squid fisheries in variable environments[J]. Fisheries Research, 2001, 54(1): 3-8. |
| [30] |
AGNEW D J, BEDDINGTON J R, HILL S L. The potential use of environmental information to manage squid stocks[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2002, 59(12): 1851-1857. DOI:10.1139/f02-150 |
| [31] |
SCHNUTEJ. A general theory for analysis of catch and effort data[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1985, 42(3): 414-429. DOI:10.1139/f85-057 |
2. National Distant-water Fisheries Engineering Research Center, Shanghai 201306, China;
3. The key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai 201306, China;
4. Key Laboratory of Oceanic Fisheries Exploration, Ministry of Angriculture and Rural Affairs, Shanghai 201306, China