 2019,
Vol. 28
2019,
Vol. 28



2. 中国水产科学研究院东海水产研究所 渔业资源遥感信息技术重点开放实验室, 上海 200090
柔鱼(Ommastrephes bartramii),是一种具有重要经济价值的头足类渔业资源,广泛分布于全球三大洋的海域中,目前仅在西北太平洋海域具有较大规模性的商业性捕捞,作业渔场主要分布在38°N~46°N, 140°E~170°W范围内的海域,渔汛期为5月至11月中下旬。柔鱼一般栖息于200~300 m水层,晚上则游动至50 m以上的浅水层内[1-2],具有明显的昼夜垂直洄游习性,因而垂直水温结构在其渔场形成过程中具有重要影响作用,垂直的水层水温是柔鱼渔场形成和鱼群迁移的重要因子[3]。柔鱼资源种群变动对海洋环境变化较为敏感,导致种群的集群以及渔场形成会受到海洋环境变化的影响较大,从而使得可捕资源量处于较大波动状态。国内外学者[4-8]在西北太平洋柔鱼渔场环境方面进行了诸多的研究,主要集中在海表温度、叶绿素、海流等海洋环境方面。MTLRATA等[9]和YATSU等[10]研究表明柔鱼类资源与垂直水层温度有密切的相关关系,并且柔鱼的垂直活动与各个水层的温度也有关系,水温的各个剖面垂直结构在一定程度上反映了海流水系的消长情况[9-15]。鉴于渔场随着垂直水温变化而波动,本研究利用2007—2016年Argo温度剖面浮标资料反演的水层温度值,结合国内远洋渔业生产中的相关柔鱼渔业生产数据,利用广义加性模型等方法对柔鱼资源丰度与各个水层温度变量的季节性变化进行分析,以及对温跃层上下界的渔获量分布特征开展研究,以期为我国西北太平洋公海海域的渔情预报以及渔业生产提供科学依据和技术参考。
1 材料与方法 1.1 渔场海域西北太平洋柔鱼捕捞的主要渔场海域,主要集中在西北太平洋的公海海域,经纬度范围为140°E~180°E、38°N~46°N,主要作业区域如图 1所示。

|
图 1 西北太平洋柔鱼渔场作业海域 Fig. 1 Fishing grounds of neon flying squid in northwestern Pacific Ocean |
生产数据由中国远洋渔业协会鱿钓工作组提供,内容包括作业日期、作业经纬度、日产量(时间跨度为2007—2016年)。西北太平洋公海柔鱼渔场区域各个垂直水层结构(10、50、100、150、200、300 m)温度的数据来源Argo实时数据中心(http://www.argo.org.cn),数据提取的空间范围为38°N~45°N、150°E~160°W,网格为1°×1°,时间跨度为2007—2016年,与生产数据相符。
1.3 模型及数据处理方法利用单位捕捞努力量渔获量(catch per unit effort,CPUE)表示西北太平洋柔鱼的资源丰度[16]:
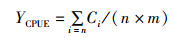 (1)
(1)
式中:YCPUE为单位捕捞努力量渔获量,t/(d·v);Ci为第i天的产量,t;n为总天数,d;m为总船次,v。
利用广义加性模型(generalized additive models,GAM)研究渔获量与各水层水温的相关关系[17]。利用一种非参数化的多元线性回归方法,非参数函数可通过数据平滑技术处理得到。建立以YCPUE为非独立变量、以主要的环境因子为解释变量的GAM模型[18],其表达式如下:
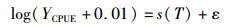 (2)
(2)
为了防止取对数时出现零值,先将YCPUE加上0.01,再进行对数化处理,其中s为自然立方样条平滑(natural cube spline smoother);T为海水温度;ε为误差项;将GAM模拟结果中上、下95%的置信区间虚线离实线最接近的区间定义为渔场最适环境范围。
采用Akima插值方法处理深度上分布不均匀的Argo浮标剖面温度资料,并计算温度剖面的梯度[19];根据温跃层定义标准,并采用温度垂直梯度法进行温跃层的判定,提取单点剖面处温跃层特征参数(温跃层上界深度、温度和温跃层下界深度、温度)[20]。
2 结果与分析 2.1 柔鱼资源丰度的年际变化2007—2016年西北太平洋公海柔鱼的年际产量显示,2007年产量最高,2012年产量最低。各月份平均CPUE显示:7月之前CPUE较低,渔汛未爆发;8月和9月是最高产的月份,为盛渔期;10月之后渔汛开始减弱。见图 2。

|
图 2 西北太平洋公海柔鱼渔获量和月平均CPUE的变化 Fig. 2 Changes in annual catch and monthly mean CPUE of neon flying squid in northwestern Pacific Ocean |
接近表层10 m水深水层的海水温度,5—6月集中在13~16 ℃, 7月集中在15~20 ℃,8月集中在17~23 ℃,9月集中在16~22 ℃,10月集中在14~18 ℃,11月集中在11~15 ℃,P检验显示显著(P<0.01),季节性的变化比较明显,见图 3。

|
图 3 表层水温(10 m)的季节性变化 Fig. 3 Seasonal changes of surface temperature (10 m) |
100 m水层的温度季节性变化中,5—6月海水温度集中在9~13 ℃,7月海水温度集中在7~14 ℃,8月海水温度集中在3~11 ℃,9月海水温度集中在2~11 ℃,10月海水温度集中在2~10 ℃,11月海水温度分布较散,集中在4~10 ℃,P检验显著(P<0.05),季节性变化明显,见图 4。

|
图 4 100 m水层水温的季节性变化 Fig. 4 Seasonal changes of 100 m layer water temperature |
200 m水层的温度季节性变化中,5—6月海水温度集中在4~11 ℃,7月海水温度集中在3~8 ℃,8月海水温度集中在3~8 ℃,9月海水温度集中在3~7 ℃,10月海水温度集中在3~6 ℃,11月海水温度分布较散,集中在3~5 ℃,P检验显著(P<0.05),季节性变化明显,见图 5。

|
图 5 200 m水层水温的季节性变化 Fig. 5 Seasonal changes of 200 m layer water temperature |
300 m水层的温度季节性变化中,5—6月海水温度集中在4~12 ℃,7月海水温度集中在4~10 ℃,8月海水温度集中在3~8 ℃,9月海水温度集中在3~7 ℃,10月海水温度集中在3~7 ℃,11月海水温度分布较散,集中在3~6 ℃,P检验显著(P<0.05),季节性变化明显,见图 6。

|
图 6 300 m水层水温的季节性变化 Fig. 6 Seasonal changes of 300 m layer water temperature |
渔汛初期5—6月各水层的水温差距较平稳,没有明显温差,其作业区域主要集中在160°E以东海域;7月开始0~50 m的水温温差逐步拉大,0~50 m的平均温差达到了5.17 ℃,0~100 m的温差为7.68 ℃;8—9月温差幅度最大,8、9月0~50 m的平均温差分别为9.25 ℃和9.89 ℃,0~100 m的平均温差分别为11.94 ℃和12.64 ℃;渔汛后期10—11月0~50 m、0~100 m的水层温差逐步减小。见图 7。

|
图 7 不同月份渔场的垂直水温变化 Fig. 7 Vertical water temperature changes in fishing grounds for each month |
西北太平洋公海中心渔场海域温跃层上界深度大致在20~50 m,其对应温跃层上界的海水温度范围在5~19 ℃,具有明显的季节性变化,渔场重心在11 ℃等温线附近波动,图 8。下界深度大致在150~230 m范围,具有一定的季节性变化,对应的温跃层下界的海水温度范围为3~12 ℃,渔场重心在7 ℃等温线附近波动,见图 9。同时从渔获量分布图可以得到,柔鱼资源的分布随着水层温度变化而变动,渔汛初期(5—7月中上旬)渔场分散,7月下旬到8、9、10月中心渔场逐渐集中聚拢,11月份渔场逐步分散。

|
图 8 渔获量与温跃层上界深度水层温度的叠加分布 Fig. 8 Distribution of catches and temperature of upper bounds of the thermocline |

|
图 9 渔获量与温跃层下界深度水层温度的叠加分布 Fig. 9 Distribution of catches and temperature of lower bounds of the thermocline |
对每月中0~50 m和0~100 m的水温差值变化与对应CPUE变化趋势进行对照比较,结果表明无论是0~50 m还是0~100 m水温差值变化越大,CPUE就越高,尤其是当ΔT0-50 m>8 ℃和ΔT0-100 m>10 ℃时,8月和9月CPUE值为最高,但有一个滞后交叉现象(图 10),根据检验ΔT0-50 m与CPUE相关性系数R2=0.739,P<0.05;ΔT0-100 m与CPUE相关性R2=0.838,P<0.05,相关性皆为显著。

|
图 10 各月份柔鱼CPUE与0~50 m及0~100 m的垂直温差的分布 Fig. 10 Vertical temperature distribution for ΔT0-50 m & ΔT0-100 m and CPUE of neon flying squid for each month |
为深入了解西北太平洋公海渔场的变化和波动趋势,研究柔鱼的昼夜垂直移动习性是重要前提。渔汛初期5—6月和7月中上旬渔场主要集中在160°E以东海域,渔船一般进行深水鱿钓,柔鱼栖息的水层在150~300 m,该渔场的柔鱼个体偏大,白天和晚上都可开展鱿钓捕捞生产,此时在160°E以西海域传统渔场(即150°E~160°E、39°N~45°N)并未到渔汛期。原因可能是:海表水温未上升至柔鱼的适宜温度,垂直水温在50 m到表层未形成明显的较大温差,温跃层未形成(图 9);或者是柔鱼还未进行索饵洄游[21];或者2个原因皆有。7月中下旬黑潮和亲潮锋区在西北太平洋40°N位置附近相遇处形成锋区海洋涡流,在锋区涡旋场,形成了温跃层中较大的等温面及与其相应的温度和密度水平差,为柔鱼渔场的形成与迁徙创造了条件[15]。7月中下旬随着传统渔场海域水温的上升,温跃层50 m水层内逐步形成大幅度温差,生产渔场重心逐步从东部往西部移动,西部传统渔场渔汛爆发,所以5、6、7月整个作业的渔场区域分布较广。8—9月,渔场表层至50 m水层的温差急剧下降,为所有月份中表层水温到50 m温差最大的2个月份,此时的CPUE最高;10—11月表层至50 m温差逐步缩小,渔获产量同步减小。7月中下旬之后在传统渔场作业都是夜间鱿钓,该时期夜间柔鱼的主要游泳水层为表层至50 m水层间,初步推测与柔鱼的饵料生物中某种浮游甲壳类动物和灯笼鱼等小型鱼类的分布有关[21]。
除了生物自身生理作用外,ΔT0-50 m和ΔT0-100 m较大温差可能是导致浮游动物中的甲壳类昼夜垂直移动为夜间上浮的驱动因子之一,而灯笼鱼等一些小型鱼类也进行昼夜垂直移动,夜间上浮至表层,温差驱动这些鱼类改变活动水层。因而,ΔT0-50 m和ΔT0-100 m越大,柔鱼活动越频繁。海洋生物包括柔鱼昼夜垂直移动现象不但与水温环境有密切有关,而且与光照强度有密切关联[22-23]。鱿钓作业先通过配备水上和水下集鱼灯进行灯光诱集,进而提高捕捞效率。这亦解释了海面上月光大的时候柔鱼资源容易分散,而阴霾天气渔获产量反而高的渔业生产现象。柔鱼类是中上层游泳动物,与多数鱼类一样,有昼夜垂直移动的习性,其栖息水层在很大程度上取决于水温的垂直结构[24]。温跃层像一道天然屏障,影响着鱼类的上下迁移。另外,利用垂直迁移和生活习性寻找中心渔场,海水温度随着地理位置、海区特性的不同,其垂直分布也会有所差异,所以对鱼群集群活动及渔场变化影响作用并不一致[25]。
3.2 渔场垂直水温结构与柔鱼集群的关系各水层水温的季节性变化分析结果表明,作业海域渔汛期表层水温在11~23 ℃,100 m水层水温在2~14 ℃,200 m水层温度在3~11 ℃,300 m水层温度在3~12 ℃,200~300 m水温变化微小。昼夜垂直移动与小型动物之间的关系十分密切,夜间的主要游泳层为表层到50 m内水层,这可能与其饵料生物中某种浮游甲壳动物和灯笼鱼等小型鱼类的趋光特性上浮分布有关系。可见西北太平洋公海柔鱼渔场在索饵场的昼夜垂直移动规律是夜间主要栖息于表层到50 m水层;白天主要栖息于200~300 m,这与陈峰等[13]的研究结果相同,但是区别在于本研究得出结果各水层温度相对更低。7月下旬开始渔场海域水深在20~50 m处出现显著的温跃层(图 8-9),柔鱼夜晚主要栖息层处在温跃层附近或在温跃层与海面之间,所以出现渔汛旺期。在水温急剧下降的水层,往往出现水温垂直梯度大的温跃层。温跃层的存在与浮游生物、鱼类活动的关系甚为密切,特别是中上层鱼类的分布和温跃层的形成与消长关系更为密切[26]。本研究发现:在160°E以东海域的温跃层未形成或者非常不明显,所以渔场分散渔获产量低;在150°E~160°E、39°N~45°N的西部传统渔场,不管是0~50 m还是0~100 m的水温差变化较大,当月柔鱼渔获量也相应高产,因而推断垂直变化的水温差驱动了鱼类的大面积移动和集群活动。
温跃层上下界深度对应的温度相差和ΔT0-50 m、ΔT0-100 m的水温季节性变化相似,都是在8、9月温差幅度最大。温跃层上界深度在20~50 m位置,等温线11 ℃可以作为中心渔场形成的指标;温跃层下界深度在150~230 m位置,等温线7 ℃可以作为该水层中心渔场形成的指标。因为柔鱼喜欢聚集的空间位置取决于最优温度区域,特别是从垂直结构来看,在夏季表层为高温,随着水深变大而温度变低,在0~100 m水温降幅较大,但是到了一定深度之后水温较稳定,温度变化幅度减小,海水表现出垂直稳定分层状态。由于海面上的风等外力作用[27],在交汇区上层海水混合交换作用下形成一个上均匀层,从而在垂直方向上形成垂直等温剖面与垂直分层剖面,这两种垂直结构对渔场的形成与变动都有驱动作用,所以渔场垂直水温结构的变化达到一定的特征是产生高产渔场的前提条件。海水温度的垂直结构的变化对柔鱼索饵洄游、栖息水层、趋光集群与散群等有着密切的关系;海水温度随时间的变化(日变化、月变化、季节变化和年变化等大小周期变化)会引起鱼类的垂直洄游及集群索饵:该类现象对渔业生产有明显的指示作用。因而,了解各水层水温分布特征有助于寻找其中心渔场,提高生产效率[28]。
4 结论利用2007—2016年西北太平洋柔鱼的渔获数据,结合同期Argo反演的各个水层的温度数据,分析了柔鱼渔场各垂直剖面水温的季节性变化,与渔汛期表层到300 m水层的海水温度,其中:8、9月0~50 m的平均温差为9.25 ℃和9.89 ℃,0~100 m的平均温差为11.94 ℃和12.64 ℃,温差越大对应的CPUE越高;其他月份ΔT0-50 m和ΔT0-100 m值较小,温差小对应渔场CPUE低。柔鱼渔场海域温跃层上界深度在20~50 m范围内,其对应温跃层上界的海水温度范围在5~19 ℃,渔场重心在11 ℃等温线上附近来回摆动。温跃层下界深度在150~230 m范围,得出对应的海水温度范围为3~12 ℃,渔场重心在7 ℃等温线上附近来回移动。利用Argo数据研究西北太平洋公海柔鱼渔场垂直水温结构的季节性变化特征,可以帮助判断柔鱼的中心渔场位置,并可以为远洋鱿钓渔业生产与管理提供参考意见。
| [1] |
村田守, 嶋津靖彦.北西太平洋海域にぉけるァカィカの资源特性值につぃて[R].北水研报告, 1982: 1-10. MURATA M, YASUHIKO Y J. According to the resource characteristic value of soft fish in the northwest Pacific Ocean[R]. North Water Research Report, 1982: 1-10. |
| [2] |
王尧耕, 陈新军. 世界大洋性经济柔鱼类资源及其渔业[M]. 北京: 海洋出版社, 2005: 124-156. WANG Y G, CHEN X J. Economy soft fish resources and fishing in the world ocean[M]. Beijing: Ocean Press, 2005: 124-148. |
| [3] |
YU W, CHEN X J, CHEN Y, et al. Effects of environmental variations on the abundance of western winter-spring cohort of neon flying squid (Ommastrephes bartramii) in the northwest Pacific Ocean[J]. Acta Oceanologica Sinica, 2015, 34(8): 43-51. DOI:10.1007/s13131-015-0707-7 |
| [4] |
MURATA M, HAYASE S. Life history and biological information on flying squid (Ommastrephes bartramii) in the north Pacific Ocean[J]. International north Pacific Fisheries Commission Bulletin, 1993, 53: 147-182. |
| [5] |
YU W, CHEN X J, YI Q. Interannual and seasonal variability of winter-spring cohort of neon flying squid abundance in the northwest Pacific ocean during 1995-2011[J]. Journal of Ocean University of China, 2016, 15(3): 480-488. DOI:10.1007/s11802-016-2884-1 |
| [6] |
田思泉, 陈新军, 冯波, 等. 西北太平洋柔鱼资源丰度与栖息环境的关系及其时空分布[J]. 上海海洋大学学报, 2009, 18(5): 586-592. TIAN S Q, CHEN X J, FENG B, et al. Spatio-temporal distribution of abundance index for Ommastrephes bartramii and its relationship with habitat environment in the northwest Pacific Ocean[J]. Journal of Shanghai Ocean University, 2009, 18(5): 586-592. |
| [7] |
沈新强, 樊伟, 崔雪森. 西北太平洋柔鱼渔场分布与水温关系的研究[J]. 海洋水产研究, 2004, 25(3): 10-14. SHEN X Q, FAN W, CUI X S. Study on the relationship of fishing ground distribution of Ommastrephes bartrami and water temperature in the northwest Pacific Ocean[J]. Marine Fisheries Research, 2004, 25(3): 10-14. |
| [8] |
陈新军, 田思泉. 西北太平洋海域柔鱼的产量分布及作业渔场与表温的关系研究[J]. 中国海洋大学学报, 2005, 35(1): 101-107. CHEN X J, TIAN S Q. Study on the catch distribution and relationship between fishing ground and surface temperature for Ommastrephes bartrami in the northwestern Pacific Ocean[J]. Periodical of Ocean University of China, 2005, 35(1): 101-107. |
| [9] |
MTLRATA M, NAKAMURA Y. Seasonal migration and diel vertical migration of the neon flying squid, Ommastrephes Bartramii, in the north Pacific[C]//International Symposium on Large Pelagic Squids, July 18-19, 1996, for JAMARC's, 25th Anniversary of its Foundation. Tokyo: Japan Marine Fishery Resources Research Center, 1998: 13-30.
|
| [10] |
YATSU A, WATANABE T. Interannual variability in neon flying squid abundance and oceanographic conditions in the Central north Pacific, 1982-1992[J]. Bulletin of the National Research Institute of Far Seas Fisheries, 1996, 33: 123-138. |
| [11] |
陈新军, 许柳雄. 北太平洋150°~165°E海域柔鱼渔场与表温及水温垂直结构的关系[J]. 海洋湖沼通报, 2004(2): 36-44. CHEN X J, XU L X. Analysis of relationship between fishing ground of Ommastrephe bartrami and surface water temperature and its vertical distribution from 150°E to 165°E in the northwestern Pacific[J]. Transactions of Oceanology and Limnology, 2004(2): 36-44. DOI:10.3969/j.issn.1003-6482.2004.02.006 |
| [12] |
范江涛, 陈新军, 曹杰, 等. 西北太平洋柔鱼渔场变化与黑潮的关系[J]. 上海海洋大学学报, 2010, 19(3): 378-384. FAN J T, CHEN X J, CAO J, et al. The variation of fishing ground of Ommastrephes bartramii in the northwest Pacific concerning with Kuroshio Current[J]. Journal of Shanghai Ocean University, 2010, 19(3): 378-384. |
| [13] |
陈峰, 陈新军, 刘必林, 等. 西北太平洋柔鱼渔场与水温垂直结构关系[J]. 上海海洋大学学报, 2010, 19(4): 495-504. CHEN F, CHEN X J, LIU B L, et al. Relationship between fishing ground of Ommastrephes bartramii and vertical temperature structure in the northwestern Pacific Ocean[J]. Journal of Shanghai Ocean University, 2010, 19(4): 495-504. |
| [14] |
陈新军, 陈峰, 高峰, 等. 基于水温垂直结构的西北太平洋柔鱼栖息地模型构建[J]. 中国海洋大学学报, 2012, 42(6): 52-60. CHEN X J, CHEN F, GAO F, et al. Modeling of habitat suitability of neon flying squid (Ommastrephes bartramii) based on vertical temperature structure in the northwestern Pacific Ocean[J]. Periodical of Ocean University of China, 2012, 42(6): 52-60. |
| [15] |
YATSU A, WATANABE T, MORI J, et al. Interannual variability in stock abundance of the neon flying squid, Ommastrephes bartramii, in the north Pacific Ocean during 1979-1998:impact of driftnet fishing and oceanographic conditions[J]. Fisheries Oceanography, 2000, 9(2): 163-170. DOI:10.1046/j.1365-2419.2000.00130.x |
| [16] |
化成君, 张衡, 伍玉梅, 等. 中东太平洋金枪鱼延绳钓中心渔场的时空变化[J]. 生态学杂志, 2014, 33(5): 1243-1247. HUA C J, ZHANG H, WU Y M, et al. Spatiotemporal patterns of fishing grounds of tuna longline fishery in the mid-eastern Pacific Ocean[J]. Chinese Journal of Ecology, 2014, 33(5): 1243-1247. |
| [17] |
HASTIE T J, TIBSHIRANI R J. Generalized additive models[M]. New York: Chapman and Hall, 1990.
|
| [18] |
唐峰华, 樊伟, 伍玉梅, 等. 北太平洋柔鱼渔场资源与海洋环境关系的季节性变化[J]. 农业资源与环境学报, 2015, 32(3): 242-249. TANG F H, FAN W, WU Y M, et al. Seasonal changes of relationship between marine environment and squid fishing resources in north Pacific Ocean[J]. Journal of Agricultural Resources and Environment, 2015, 32(3): 242-249. |
| [19] |
AKIMA H. A new method of interpolation and smooth curve fitting based on local procedures[J]. Journal of the ACM, 1970, 17(4): 589-602. DOI:10.1145/321607.321609 |
| [20] |
张胜茂, 杨胜龙. Argo剖面数据在远洋金枪鱼渔业中的应用[M]. 北京: 海洋出版社, 2014: 51-53. ZHANG S M, YANG S L. Argo profile data in the application of deep-sea tuna fishery[M]. Beijing: Ocean Press, 2014: 51-53. |
| [21] |
YATSU A, WATANABE T. Interannual variability in neon flying squid abundance and oceanographic conditions in the central north Pacific, 1982-1992[J]. Bulletin of the National Research Institute of Far Seas Fisheries, 1996, 33: 123-138. |
| [22] |
唐峰华, 靳少非, 张胜茂, 等. 北太平洋柔鱼渔场时空分布与海洋环境要素的研究[J]. 中国环境科学, 2014, 34(8): 2093-2100. TANG F H, JIN S F, ZHANG S M, et al. Study for marine environmental elements on spatio-temporal distribution of neon flying squid in the north Pacific fishing ground[J]. China Environmental Science, 2014, 34(8): 2093-2100. |
| [23] |
陈新军, 田思泉, 钱卫国. 月相对北太平洋海域柔鱼钓获率的影响[J]. 海洋渔业, 2006, 28(2): 136-140. CHEN X J, TIAN S Q, QIAN W G. Effect of moon phase on the jigging rate of Ommastrephes bartrami in the north Pacific[J]. Marine Fisheries, 2006, 28(2): 136-140. DOI:10.3969/j.issn.1004-2490.2006.02.009 |
| [24] |
晏磊, 张鹏, 杨炳忠, 等. 南海鸢乌贼产量与表温及水温垂直结构的关系[J]. 中国水产科学, 2016, 23(2): 469-477. YAN L, ZHANG P, YANG B Z, et al. Relationship between the catch of Symplectoteuthis oualaniensis and surface temperature and the vertical temperature structure in the South China Sea[J]. Journal of Fishery Sciences of China, 2016, 23(2): 469-477. |
| [25] |
杨胜龙, 张胜茂, 蒋兴伟, 等. 热带大西洋大眼金枪鱼和黄鳍金枪鱼渔场温跃层的时空变化特征[J]. 应用海洋学学报, 2013, 32(3): 349-357. YANG S L, ZHANG S M, JIANG X W, et al. Temporal and spatial variation characteristics of the thermocline in Thunnus obesus and Thunnus albacares fishing grounds in the tropical Atlantic Ocean[J]. Journal of Applied Oceanography, 2013, 32(3): 349-357. DOI:10.3969/J.ISSN.2095-4972.2013.03.007 |
| [26] |
YU W, CHEN X J, CHEN C S, et al. Impacts of oceanographic factors on interannual variability of the winter-spring cohort of neon flying squid abundance in the northwest Pacific Ocean[J]. Acta Oceanologica Sinica, 2017, 36(10): 48-59. DOI:10.1007/s13131-017-1069-0 |
| [27] |
王文宇, 周成虎, 邵全琴, 等. RS/GIS支持下的柔鱼中心渔场时空动态迁移研究[J]. 高技术通讯, 2003, 13(11): 90-93. WANG W Y, ZHOU C H, SHAO Q Q, et al. An application of rs/gis on study on the migration dynamics of Ommastrephes bartrami[J]. High Technology Letters, 2003, 13(11): 90-93. DOI:10.3321/j.issn:1002-0470.2003.11.021 |
| [28] |
CHEN X J, TIAN S Q, GUAN W J. Variations of oceanic fronts and their influence on the fishing grounds of Ommastrephes bartramii in the northwest Pacific[J]. Acta Oceanologica Sinica, 2014, 33(4): 45-54. DOI:10.1007/s13131-014-0452-3 |
2. Key Laboratory of Fishery Remote Sensing and Information, East China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Shanghai 200090, China