 2019,
Vol. 28
2019,
Vol. 28



2. 上海海洋大学 港航生态安全研究中心, 上海 201306
浮游植物是指生活史的一部分或全部在海洋、湖泊、池塘或河流中漂浮生活的光合自养生物[1],是河口生态系统的初级生产者,处于食物链营养级的基础,直接或间接地为河口水域和沉积物中的浮游动物、鱼虾类及其幼体提供主要的食物来源。浮游植物的物种组成、群落结构特征及分布不仅可以直接影响水域生态系统,而且可以改善水体的物质循环、能量流动和自我净化效率,以及能够调节水域生态系统的平衡[2-5]。另外,水体环境条件的变化也会引起浮游植物群落特征的变化,例如水温、溶氧量和pH等非生物因素变化会使浮游植物的种类和数量升高或降低,一些生物因素如鱼的捕食等也会引起浮游植物的改变[6-7]。
长江口地处中国东部沿海地区,是我国最大的入海河口。长江口周边的发展以及受径流、潮汐、沿岸流和降水等多方面环境因子的影响,给该区域环境造成巨大的压力,富营养化和赤潮问题严重[8-10]。长江口南支水域流量大,受长江径流影响较为明显,一般显示出明显的淡水特性,北支水域受长江径流影响较小,主要受潮流的影响,含盐度显著高于南支,显示出一定的海水特征,这就形成了南北支水域中浮游植物结构组成的差异[9-12]。关于长江口及其附近海域的浮游植物群落结构与时空分布等有较多的研究[13-18],但目前的研究一般集中于长江口的南支水域[2, 16]、北支水域[19-20]以及邻近海域[2, 21],没有对南、北分支及分支前主河道的浮游植物进行较为全面的比较。笔者通过对长江口南、北分支和分支前主河道的浮游植物进行调查,分析在长江口复杂水文条件下不同水域浮游植物群落结构特征,探讨影响浮游植物分布的影响因子,以期丰富对河口浮游植物与环境关系的认识。
1 材料与方法 1.1 样本的采集和处理依据《海洋监测规范》[22]相关内容,于2010年8月至2010年9月在长江河口口门内水域潮下带开展水环境因子的监测和浮游植物的采样。根据当地的水动力学和泥沙沉积情况等环境特征,共设置7个采样断面18个采样点,主要集中在长江口口门段内的潮下带水域,包括9个南支采样点、6个北支采样点以及3个主河道采样点,采样点具体分布如图 1所示。

|
图 1 长江口浮游植物采样断面分布示意图 Fig. 1 The sampling stations in the Yangtze River Estuary |
水温、盐度、电导率、pH、溶解氧等指标使用YSI(85-25)手持式野外水质检测仪进行现场测定。取水样装入1 L聚乙烯瓶中常温保存,用于叶绿素a和悬浮物测定。叶绿素a按《海湾生态系统观测方法》测定。悬浮物采用重量法测定。浮游植物定量样品采用浮游植物采水器采集表层水1 L,采集后立即滴加15 mL Lugol’s碘液进行固定,并带回实验室,静置沉淀48 h弃上清,浓缩至50 mL后滴加福尔马林保存,显微镜镜检,进行浮游植物的物种鉴定和计数[23-25]。
1.2 数据分析物种优势种用优势度指数表示, 公式为
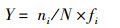 (1)
(1)
式中:ni为第i种物种个体数,N为总个体数,fi为第i种物种在总站点中出现频率。当Y>0. 02时,表示该物种为优势种。
物种多样性指数(H′)的计算采用香农-威纳指数(Shannon-Wiener index),公式为
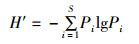 (2)
(2)
物种丰富度指数(D)采用Margalef的计算公式:
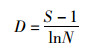 (3)
(3)
物种均匀度指数(J)采用Pielou的计算公式:
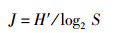 (4)
(4)
式中:S为样品中的种类总数,N为样品中所有种类的总个体数,Pi为第i个物种的个体数与样品中的总个体数的比值。
通过数据分析软件PASW Statistics 18.0对长江口南支、北支以及主河道浮游植物细胞密度进行One-way ANOVA分析和浮游植物细胞密度与环境因子Pearson相关性分析。
采用国际标准通用软件Canoco for Windows 4.5软件,对长江口浮游植物数据和环境因子进行典范应对分析(canonical correspondence analysis,CCA)。排序之前,对物种数据进行筛选,用于排序的物种,其密度占总密度比例≥5%,筛选过后用于CCA分析的浮游植物物种编号见表 1。用于分析的环境因子有溶氧(DO)、温度(Tm)、电导率(Conductivity)、盐度(salinity)、pH、叶绿素a(Chl.a)和悬浮物(TSS)。

|
表 1 长江口CCA分析中的浮游植物编号 Tab.1 Codes of phytoplankton species for CCA |
18个采样点采集的浮游植物样品,共鉴定到浮游植物8门,71属,157种。其中:硅藻门种类最多达61种,占总种类数的39%;其次为绿藻门41种,占26%;蓝藻门35种,占22%;裸藻门7种,占4%;甲藻门6种,占4%;黄藻门4种,占3%;金藻门2种,占1%;隐藻门1种,占1%。对各样点的种类数进行统计(图 2),平均物种数为32种,其中:南支水域的S1样点的物种数最多,为57种; 北支水域的S13样点的物种数最少,仅有19种。结果显示南支水域的浮游植物种类数较多,北支水域和主河道水域的种类数相对较少,可达极显著差异(P<0.01)。

|
图 2 长江口各样点浮游植物种类数变化 Fig. 2 Variation in the number of phytoplankton species at the stations in the Yangtze River estuary |
对长江口各取样点的优势种进行分析(表 2),发现南支和主河道水域的优势种都来自蓝藻门,而北支的优势种来自硅藻门。蓝藻门的湖泊鞘丝藻(Lyngbya limnetica)是全年优势种中优势度最高的,常见种为蓝藻门的阿氏颤藻(Oseillatoria agardhii),两栖颤藻(amphibia),小颤藻(Oseillatoria tenuis),绿藻门的小球藻(Chlorella vulgaris),以及硅藻门Oseillatoria的中肋骨条藻(Skeletonema costatum)。

|
表 2 长江口浮游植物各样点优势种、平均细胞密度及优势度 Tab.2 The dominant species, average abundance and dominance of phytoplankton in the Yangtze River Estuary |
2010年长江口环岛采样个体数最多的样点为S6(图 3),其个体数量达到1.50×106个/L,个体数最少的样点为S8,其个体数量为3.27×104个/L,18个样点的平均个体数为3.31×105个/L。在植物细胞密度方面,南北支水域以及主河道水域没有差别。

|
图 3 长江口各样点浮游植物细胞密度变化 Fig. 3 The variations of cells density of phytoplankton species at the stations in the Yangtze River Estuary |
分析Pielou均匀度指数、Margalef丰富度指数和Shannon-Wiener多样性指数结果见图 4。各样点的均匀度指数变化比较平稳,仅在S6样点偏低,达0.06,其余各样点均在0.30~0.79之间波动,S8样点的均匀度最高,达到了0.79;丰富度指数变化较大,在1.45~4.16之间:南支的丰富度指数偏高,平均为3.38,最高点出现在S1样点,为4.16;北支和主河道的丰富度指数较低,分别为1.82和1.93,最低点为S14样点的1.453。Shannon-Wiener多样性指数变化和丰富度指数变化比较一致:南支的多样性指数除S6样点外,整体较高(H′>2),平均为2.15;北支和主河道的多样性指数偏低,平均为1.57和1.84。

|
图 4 长江口浮游植物多样性指数变化 Fig. 4 The diversity index variation of phytoplankton species at the stations in the Yangtze River Estuary |
在长江口河口口门内水域潮下带18个采样点所检测到的环境因子中,各样点溶氧量浓度的变化范围为3.60~6.62 mg/L,平均值为5.06 mg/L;各样点电导率的变化范围为8.31~3 042.33 ms,平均值为566.48 ms;各样点水温的变化范围为25.50~32.37 ℃,平均值为29.39 ℃,各样点之间水温的差异较小;各样点盐度的变化范围为0.20~8.31,南北支盐度差异较大,南支和主河道的盐度均为0.20,北支的盐度平均为5.76;pH的变化范围为8.17~8.78,平均值为8.50,变幅较小;各样点叶绿素a浓度的变化范围为0.56~2.67 μg/L,平均值为1.59 μg/L。
利用PASW Statistics 18.0分析软件对各个样点的溶氧量、温度、电导率、盐度、pH、叶绿素a的六项相关环境因子与物种数、Margalef丰富度指数、多样性指数、均匀度指数、蓝藻细胞密度、硅藻细胞密度共13项进行Pearson相关性分析(表 3),结果如下:物种数和丰富度指数分别与温度、叶绿素a显著正相关,与盐度显著负相关;多样性指数和均匀度指数分别与温度、叶绿素a显著正相关,与电导率显著负相关;蓝藻细胞密度与温度显著正相关,与溶氧量、盐度显著负相关;硅藻细胞密度与电导率显著正相关,与温度显著负相关。

|
表 3 长江口浮游植物群落与环境因子Pearson相关性分析 Tab.3 The Pearson correlation analysis between phytoplankton abundance and environmental factors |
用CCA对13种浮游植物(表 1)和7种环境因子的关系进行分析,前两个排序轴的特征值为0.768和0.481,浮游植物和环境因子相关系数分别为0.970和0.758,说明排序图能够很好地反映物种与环境的关系。前两个排序轴所能解释的浮游植物种类与环境之间关系的解释量为76.4%。对物种与环境因子排序,盐度(r=0.613 7)、电导率(r=0.583 1)、溶氧量(r=0.624 8)与轴1表现出较高的正相关性,温度(r=-0.897 3)、叶绿素a(r=-0.669 2)与轴一表现出较高的负相关性。因此对调查水域浮游植物种类和密度的分布影响较为显著的环境因子为:盐度、电导率、溶氧量、温度和叶绿素a。
浮游植物物种分布在环境因子综合作用下呈分化现象,位置相近物种对环境条件的适应性比较一致。如图 5所示,南支水域的主要优势种湖泊鞘丝藻(Lyngbya limnetica)、小颤藻(Oseillatoria tenuis)、极大螺旋藻(Spirulina maxima)、两栖颤藻(Oseillatoria amphibia)、小席藻(Phormidium tenu)集中分布,都受到温度的正相关影响,与盐度、电导率呈负相关;北支水域的主要优势种小环藻(Cyclotella sp.)、阿氏颤藻(Oseillatoria agardhii)、中肋骨条藻(Skeletonema costatum)受电导率、盐度的正相关影响。具缘微囊藻(Microcystis marginata)受悬浮物的正相关影响。

|
图 5 浮游植物物种和环境因子的CCA排序图 Fig. 5 CCA biplot of phytoplankton species and environmental variables in the Yangtze River Estuary |
由于环境特征的不同,长江口的不同水域的浮游植物组成差异较为明显:长江口南支水域,夏季降水丰富,入海径流多,河流淡水稀释作用强,使盐度降低,该区域的优势种主要为淡水藻,如蓝藻门的湖泊鞘丝藻,由于夏季水温也较高,也适合蓝藻的生长[26];长江口北支水域的径流量明显少于南支,受潮汐的影响大,形成了独特的高盐水域环境,在这种河口淡水与海水交界区域,出现了与长江口邻近海域相同的优势种中肋骨条藻[27-28],中肋骨条藻是广温广盐的典型代表种,适宜在该盐度多变的环境中生存;在南北支分支前的主河道水域,由于盐度与南支水域相似,其优势种也是蓝藻门的湖泊鞘丝藻。
长江口在长江径流和倒灌海水等水流的共同作用下,水环境极其复杂。南北支的径流量和盐度等差异造成浮游植物群落空间分布的差异。对夏季长江口浮游植物的Margalef丰富度指数和Shannon-Wiener多样性指数分析,各样点物种多样性和丰富度变化趋势基本相同,南支水域的物种数和丰富度显著高于北支水域和主河道水域。
3.2 浮游植物群落特征和环境因子的关系另外,浮游植物的群落特征及变化也受温度、叶绿素a等多个环境因子综合作用的影响[29-30]。以细胞密度为物种变量,研究了浮游植物与环境因子之间的相关性,结果显示细胞密度分别与温度和叶绿素a显著正相关。在CCA分析中,温度也是影响浮游植物的显著性环境因子,主要是因为温度变化会影响浮游植物细胞新陈代谢的速率,也会对生物体内酶的活性和细胞分裂周期等造成不同程度的影响,特别是在水体环境中,温度变化幅度比陆地温度变化相对较小,导致水生生物缺乏相应的温度补偿机制,因此浮游植物的细胞密度与温度有一定的相关性[31-32]。蓝藻细胞密度与温度呈正相关,而硅藻细胞密度与温度呈负相关,这与之前的对长江口破碎带硅藻细胞与温度呈负相关的研究结果一致[33]。水域的环境因子可以影响水体叶绿素a的分布,同时叶绿素a也是反映该水域生态系统现状的重要指标。有研究表明在长江口丰水期,最高叶绿素a含量与浮游植物最大数量同时出现于长江口冲淡水区[32]。物种数、总细胞丰富度、多样性指数和均匀度指数均与叶绿素a显著正相关。盐度效应主要是由于长江口受陆地径流和外海高盐水共同影响,造成南北盐度差异较大,从而对浮游植物分布和种类组成产生重要影响。在调查水域,盐度较低的南支和主河道水域的优势种都来自蓝藻门的淡水藻,而盐度较高的北支的优势种则来自硅藻门的广温广盐藻。CCA分析结果表明盐度对不同的藻类的生长影响不同,因此,浮游植物群落与它们生长的水体环境息息相关,不同的浮游植物对于同一环境反应不同,不同环境因子的排列组合也会影响浮游植物的生长。
| [1] |
钱树本, 陈国蔚. 浮游植物生态[J]. 山东海洋学院学报, 1986(2): 26-55, 85-86. QIAN S B, CHEN G W. Phytoplankton ecology[J]. Journal of Ocean University of Shandong, 1986(2): 26-55, 85-86. |
| [2] |
林峰竹, 吴玉霖, 于海成, 等. 2004年长江口浮游植物群落结构特征分析[J]. 海洋与湖沼, 2008, 39(4): 401-410. LIN F Z, WU Y L, YU H C, et al. Phytoplankton community structure in the Changjiang estuary and its adjacent waters in 2004[J]. Oceanologia et Limnologia Sinica, 2008, 39(4): 401-410. DOI:10.3321/j.issn:0029-814X.2008.04.016 |
| [3] |
MALLIN M A, CAHOON L B, MCIVER M R, et al. Alternation of factors limiting phytoplankton production in the Cape Fear River Estuary[J]. Estuaries, 1999, 22(4): 825-836. DOI:10.2307/1353064 |
| [4] |
FLEMING-LEHTINEN V, LAAMANEN M, KUOSA H, et al. Long-term development of inorganic nutrients and chlorophyll α in the open northern Baltic Sea[J]. Ambio:A Journal of the Human Environment, 2008, 37(2): 86-92. DOI:10.1579/0044-7447(2008)37[86:LDOINA]2.0.CO;2 |
| [5] |
FRONEMAN P W. Feeding studies on selected zooplankton in a temperate estuary, South Africa[J]. Estuarine, Coastal and Shelf Science, 2000, 51(5): 543-552. DOI:10.1006/ecss.2000.0700 |
| [6] |
周绪申, 李慧峰, 潘曼曼, 等. 海河蓝藻暴发期间浮游植物群落结构分析[J]. 人民黄河, 2013, 35(1): 63-65. ZHOU X S, LI H F, PAN M M, et al. Study on phytoplankton community during the cyanobacteria bloom in Haihe river[J]. Yellow River, 2013, 35(1): 63-65. DOI:10.3969/j.issn.1000-1379.2013.01.020 |
| [7] |
孙晓庆, 董树刚, 汤志宏. 营养盐和光照对浮游植物群落结构的影响[J]. 南方水产, 2008, 4(1): 1-9. SUN X Q, DONG S G, TANG Z H. Influences of nutrients and illuminance on phytoplankton community structure[J]. South China Fisheries Science, 2008, 4(1): 1-9. DOI:10.3969/j.issn.2095-0780.2008.01.001 |
| [8] |
刘录三, 李子成, 周娟, 等. 长江口及其邻近海域赤潮时空分布研究[J]. 环境科学, 2011, 32(9): 2497-2504. LIU L S, LI Z C, ZHOU J, et al. Temporal and spatial distribution of red tide in Yangtze River estuary and adjacent waters[J]. Environmental Science, 2011, 32(9): 2497-2504. |
| [9] |
张桂成.长江口及其邻近海域溶解有机氮的生物可利用性及其在赤潮爆发过程中的作用研究[D].青岛: 中国海洋大学, 2015. ZHANG G C. Bioavailability of dissolved organic nitrogen and its role during the outbreak of harmful algal blooms in Changjiang River Estuary and its adjacent sea[D]. Qingdao: Ocean University of China, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10423-1015712679.htm |
| [10] |
亢振军, 于仁成, 孔凡洲, 等. 长江口南部赤潮区春季表层沉积物中色素组成、含量与分布状况[J]. 海洋科学, 2014, 38(10): 30-39. KANG Z J, YU R C, KONG F Z, et al. Pigment composition, content and distribution in surface sediment of the red-tide zone south to the Changjiang River estuary in spring[J]. Marine Sciences, 2014, 38(10): 30-39. DOI:10.11759/hykx20130423001 |
| [11] |
曹民雄, 高正荣, 胡金义. 长江口北支水道水沙特性分析[J]. 人民长江, 2003, 34(12): 34-36. CAO M X, GAO Z R, HU J Y. Analysis on flow and sediment characteristics of north branch of the Yangtze estuary[J]. Yangtze River, 2003, 34(12): 34-36. DOI:10.3969/j.issn.1001-4179.2003.12.015 |
| [12] |
殷晓龙, 徐兆礼. 长江口南支、北支、北港及口外水域浮游动物群聚相似性[J]. 生态学报, 2016, 36(18): 5621-5631. YIN X L, XU Z L. Similarity analysis of zooplankton assemblages among different zones in the Yangtze River estuary[J]. Acta Ecologica Sinica, 2016, 36(18): 5621-5631. |
| [13] |
GAO X L, SONG J M. Phytoplankton distributions and their relationship with the environment in the Changjiang Estuary, China[J]. Marine Pollution Bulletin, 2005, 50(3): 327-335. DOI:10.1016/j.marpolbul.2004.11.004 |
| [14] |
JIANG Z B, LIU J J, CHEN J F, et al. Responses of summer phytoplankton community to drastic environmental changes in the Changjiang (Yangtze River) estuary during the past 50 years[J]. Water Research, 2014, 54: 1-11. DOI:10.1016/j.watres.2014.01.032 |
| [15] |
江志兵, 曾江宁, 李宏亮, 等. 长江口及其邻近陆架区夏季网采浮游植物及其影响因素[J]. 海洋学报, 2014, 36(6): 112-123. JIANG Z B, ZENG J N, LI H L, et al. Distribution of net-phytoplankton community in summer and its influence factors in the Changjiang Estuary and adjacent continental shelf[J]. Acta Oceanologica Sinica, 2014, 36(6): 112-123. DOI:10.3969/j.issn.0253-4193.2014.06.014 |
| [16] |
赵冉, 孙军, 宋书群. 2006年春季长江口及其邻近水域浮游植物[J]. 海洋通报, 2013, 32(4): 421-428. ZHAO R, SUN J, SONG S Q. Phytoplankton in the Yangtze River estuary and its adjacent waters in spring 2006[J]. Marine Science Bulletin, 2013, 32(4): 421-428. |
| [17] |
何青, 孙军, 栾青杉, 等. 冬季长江口及其邻近水域的浮游植物[J]. 海洋环境科学, 2009, 28(4): 360-365. HE Q, SUN J, LUAN Q S, et al. Phytoplankton in Changjiang estuary and adjacent waters in winter[J]. Marine Environmental Science, 2009, 28(4): 360-365. DOI:10.3969/j.issn.1007-6336.2009.04.004 |
| [18] |
王云龙, 袁骐, 沈新强. 长江口及邻近水域春季浮游植物的生态特征[J]. 中国水产科学, 2005, 12(3): 300-306. WANG Y L, YUAN Q, SHEN X Q. Ecological character of phytoplankton in spring in the Yangtze River estuary and adjacent waters[J]. Journal of Fishery Sciences of China, 2005, 12(3): 300-306. DOI:10.3321/j.issn:1005-8737.2005.03.011 |
| [19] |
刘笑, 薛俊增, 吴惠仙. 长江口北支浮游植物群落结构周年变化特征[J]. 生物学杂志, 2014, 31(3): 20-24, 28. LIU X, XUE J Z, WU H X. Annual variation of phytoplankton community in the north branch of the Yangtze River estuary[J]. Journal of Biology, 2014, 31(3): 20-24, 28. DOI:10.3969/j.issn.2095-1736.2014.03.020 |
| [20] |
高月鑫, 江志兵, 曾江宁, 等. 春季长江口北支邻近海域浮游植物群落及其影响因子[J]. 海洋通报, 2018, 37(4): 430-439. GAO Y X, JIANG Z B, ZENG J N, et al. Phytoplankton community in relation to environment factors off the north branch of the Yangtze River Estuary[J]. Marine Science Bulletin, 2018, 37(4): 430-439. |
| [21] |
李俊龙, 郑丙辉, 刘录三, 等. 长江口浮游植物群落特征及其与环境的响应关系[J]. 环境科学研究, 2013, 26(4): 403-409. LI J L, ZHENG B H, LIU L S, et al. Phytoplankton community structure in the Yangtze River estuary and its relation to environmental factors[J]. Research of Environmental Sciences, 2013, 26(4): 403-409. |
| [22] |
国家质量技术监督局. GB 17378-1998海洋监测规范[S].北京: 中国标准出版社, 2004. The Quality and Technology Supervision Bureau. GB 17378-1998 The specification for marine monitoring[S]. Beijing: Standards Press of China, 2004. |
| [23] |
程兆第, 高亚辉. 中国海藻志第五卷, 第二册, Ⅰ-硅藻门, 羽纹纲, 等片藻目曲壳藻目褐指藻目短缝藻目[M]. 北京: 科学出版社, 2012. CHENG Z D, GAO Y H. Chinese seaweed[M]. Beijing: Science Press, 2012. |
| [24] |
胡鸿钧, 魏印心. 中国淡水藻类:系统、分类及生态[M]. 北京: 科学出版社, 2006. HU H J, WEI Y X. The freshwater algae of China:systematics, taxonomy and ecology[M]. Beijing: Science Press, 2006. |
| [25] |
曾呈奎. 中国经济海藻志[M]. 北京: 科学出版社, 1962. ZENG C K. Chinese economic seaweed[M]. Beijing: Science Press, 1962. |
| [26] |
谭啸, 孔繁翔, 于洋, 等. 升温过程对藻类复苏和群落演替的影响[J]. 中国环境科学, 2009, 29(6): 578-582. TAN X, KONG F X, YU Y, et al. Effects of enhanced temperature on algae recruitment and phytoplankton community succession[J]. China Environmental Science, 2009, 29(6): 578-582. DOI:10.3321/j.issn:1000-6923.2009.06.004 |
| [27] |
李俊龙, 郑丙辉, 刘录三, 等. 长江口浮游植物群落特征及其与环境的响应关系[J]. 环境科学研究, 2013, 26(4): 403-409. LI J L, ZHENG B H, LIU L S, et al. Phytoplankton community structure in the Yangtze River estuary and its relation to environmental factors[J]. Research of Environmental Sciences, 2013, 26(4): 403-409. |
| [28] |
陈立婧, 蒋琰, 龚小玲. 长江口九段沙湿地近岸水域浮游植物群落结构的特征[J]. 上海海洋大学学报, 2013, 22(1): 74-80. CHEN L J, JIANG Y, GONG X L. Characteristics of community structures of phytoplankton in the Jiuduansha inshore waters of the Yangtze River estuary[J]. Journal of Shanghai Ocean University, 2013, 22(1): 74-80. |
| [29] |
辛明.长江口海域关键环境因子的长期变化及其生态效应[D].青岛: 中国海洋大学, 2014. XIN M. Long-term variations of the key environmental factors and their ecological effects in the Changjiang Estuary[D]. Qingdao: Ocean University of China, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10423-1014369032.htm |
| [30] |
韩秀荣.长江口及邻近海域浮游植物生长的多环境效应因子影响解析研究[D].青岛: 中国海洋大学, 2009. HAN X R. Analytical study on multi-environment factors that influence the phytoplankton growth in the Changjiang Estuary and its adjacent area[D]. Qingdao: Ocean University of China, 2009. http://cdmd.cnki.com.cn/Article/CDMD-10423-2009160402.htm |
| [31] |
刘东艳.胶州湾浮游植物与沉积物中硅藻群落结构演替的研究[D].青岛: 中国海洋大学, 2004. LIU D Y. Community structure succession study of phytoplankton and sediment diatom in Jiaozhou Bay[D]. Qingdao: Ocean University of China, 2004. http://cdmd.cnki.com.cn/Article/CDMD-10423-2004131357.htm |
| [32] |
李照.长江口邻近海域浮游植物群落特征及其碳沉降研究[D].青岛: 中国科学院大学(中国科学院海洋研究所), 2018. LI Z. Phytoplankton community and its related carbon sinking in the Changjiang (Yangtze River) estuary and adjacent waters[D]. Qingdao: University of Chinese Academy of Sciences (The Institute of Oceanology, Chinese Academy of Sciences), 2018. http://cdmd.cnki.com.cn/Article/CDMD-80068-1018138301.htm |
| [33] |
张松艳, 王琼, 薛俊增, 等. 长江口碎波带浮游植物群落结构特征[J]. 生物学杂志, 2014, 31(3): 15-19. ZHANG S Y, WANG Q, XUE J Z, et al. Ecological features of phytoplankton community in the surf zones of the Yangtze River estuary[J]. Journal of Biology, 2014, 31(3): 15-19. DOI:10.3969/j.issn.2095-1736.2014.03.015 |
2. Centre for Research on the Ecological Security of Ports and Shipping, Shanghai 201306, China