 2018,
Vol. 27
2018,
Vol. 27



2. 上海海洋大学 农业部淡水水产种质资源重点实验室, 上海 201306;
3. 上海海洋大学 水产动物遗传育种中心上海市协同创新中心, 上海 201306;
4. 上海城投原水有限公司青草沙水库管理分公司, 上海 201913
青草沙水库(31°25′N-31°29′N,121°34′E-121°42′E)位于上海市崇明县,紧邻长江入海口,水库总面积达66 km2,平均每年的径流总量是4 896×108 m3,一共可容纳的水量为5.24亿m3,每日可提供719万m3的水[1]。从2010年开始使用,目前水库的整体水生态系统已相对稳定。但为保证上海市民的饮水安全,对青草沙水库的水体质量进行监测和管理十分必要。
枝角类与桡足类是浮游甲壳动物的重要组成部分,普遍存在于淡水中,主要摄食浮游植物、细菌和碎屑,其本身也是鱼类和许多水生无脊椎动物的主要饵料,在食物网中起着连接作用[2-3]。通常水体的营养水平越高,浮游甲壳类的数量及生物量也越大[4]。季节间水温的不同使其群落结构有明显差异,在同一季节中水体理化因子、食物、竞争者和捕食者则是对其产生影响的重要因素[5]。杨桂军等指出水体的营养水平可被浮游甲壳类群落结构的变动所反映,对水体质量监测具有重要指示作用[6]。刘歆璞等[7]和孙陆宇等[8]对青草沙水库运行初期的浮游动物有所研究。王少坤等[9]对水库中的轮虫种群现状作了相应调查工作,同年严文逸等[10]研究了库内浮游植物的群落结构特征以及对环境因素的响应。该水库大型底栖动物的研究也已被发表[11]。但鲜有对浮游甲壳类单独的报道,研究青草沙水库浮游甲壳类群落结构,旨在为水库水质的调控提供生物学的理论基础。
1 材料与方法 1.1 站点设置与采样时间根据青草沙水库形态、地理状况及水文特征,同时结合站点设置规范[12]共设置11个站点(图 1)。综合水库上下游闸口位置和总体水体流动情况可将这些站点分为河流区站点(S1、S10和S11)、过渡区站点(S2、S3、S4、S8和S9)和湖泊区站点(S5、S6和S7)。采样频率为每月下旬1次。

|
图 1 青草沙水库站点分布
Fig. 1 Distribution of sampling sites in Qingcaosha Reservoir
|
采样办法参照《湖泊生态系统观测方法》[12]。利用2.5 L的不锈钢柱状采水器采集水样,水深小于等于3 m时,采表层水(0.5 m处)4次;水深大于3 m时,自表层0.5 m处开始每隔1 m采水两次,采至离水底0.5 m处,然后将采集的水样混合均匀后,用直径为112 μm的13号浮游生物网过滤,过滤后的样品收集于50 mL塑料瓶中,用5%的福尔马林溶液使生物固定,带回实验室观察鉴定。镜检时需要将全部水样计入,在10×10倍光学显微镜下使用1 mL浮游生物计数框进行种类鉴定,主要参考沈嘉瑞等所编著的书籍以及相关文献[13-15]。通过体质量-体长回归方程对枝角类和桡足类的生物量进行计算[16]。
样品采集前首先用Hondex声纳测深仪测定水深、萨氏盘测定透明度,同时利用哈希HQd系列便携式水质仪测定水温、电导率、溶解氧和pH。另外采集1 L水样于塑料瓶中不加任何药品,回到实验室后进行化学指标的测定,利用哈希DR3900分光光度计检测总氮、亚硝态氮、硝态氮、氨氮、总磷及高锰酸盐指数。
1.3 数据统计与分析优势度的计算公式为:
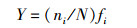 (1)
(1)
式中:ni为第i种的数量;N为共有数量;fi为第i种出现的频率,当Y > 0.02时,该种为优势种[17]。
多样性和丰富度采用Shannon-Wiene多样性指数H′和Margalef丰富度指数D来评估,公式分别为:
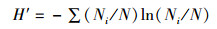 (2)
(2)
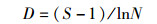 (3)
(3)
式中:Ni为样品第i种的个体数;N为样品中所有物种数之和;S为样品中浮游甲壳动物的种数。H′反映的是群落结构的复杂性及稳定程度,一般D值越大水质越好[18-19]。
均匀度指数:
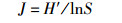 (4)
(4)
式中:H′为Shannon-Wiene多样性指数,S为样品中物种数。
Jaccard相似性指数:
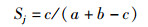 (5)
(5)
式中:a为一个区域物种种类总数;b为另一个区域种类数;c为两个区域共同的种类数。Sj < 0.25说明这两个区域物种组成极不相似,当0.25<Sj<0.5为中等不相似,Sj在0.5~0.75为中等相似,Sj > 0.75是极相似[20]。
采用Microsoft Excel进行图表制作,通过SPSS 18.0进行相关性分析,并采用CANOCO软件进行冗余分析(RDA),分析之前对数据进行筛选,选择物种相对丰度大于1%且出现频率大于25%的样品进行分析,最后得出物种与环境间的相关关系。
2 结果 2.1 理化数据指标两年间平均水深为以库尾湖泊区最大。透明度范围较广,从大到小依次是湖泊区[(86.9±22.2) cm]、过渡区[(60.5±20.7) cm]和河流区[(44.5±15.0) cm]。pH整体上呈弱碱性。比较两年理化因子的年均值可知:水温、溶氧、pH、电导率、总磷和高锰酸盐指数值在2016年相对升高,水深、透明度、总氮、硝态氮、亚硝态氮、铵态氮值有所下降,两年内各项指标的变化没有显著差异(P>0.05)。
2.2 浮游甲壳类种类组成特征及优势种两年内共鉴定出浮游甲壳类34种。枝角类(Cladocera)18种,隶属于5科10属,其中盘肠溞科(Chydoridae)和溞科(Daphniidae)种类较多,均有6种,占枝角类总种数的33.3%。桡足类(Copepoda)16种,隶属于7科12属,剑水蚤科(Cyclopidae)最多(7种),占桡足类总种数的43.8%。其中河流区发现26种浮游甲壳类,过渡区有34种,湖泊区有29种,各区之间种类数差别不大。河流区与湖泊区的浮游甲壳类种类为中等相似(Sj=0.72);河流区与过渡区为极相似(Sj=0.76);过渡区与湖泊区也为极相似(Sj=0.80)。季节不同其种类组成也略有差异,夏季最多(23种),春季次之(22种),秋季有20种,冬季最少(17种)。
2015年与2016年分别鉴定出浮游甲壳类22种、28种,2016年种类比2015年略有增加。两年间浮游甲壳类的种类为中等不相似(Sj=0.43)。

|
表 1 2015—2016年青草沙水库理化因子的年均值及其变动范围 Tab.1 Annual average and the range of main physicochemical factors in Qingcaosha Reservoir from 2015 to 2016 |

|
表 2 2015—2016年青草沙水库浮游甲壳动物种类的季节分布 Tab.2 Species distribution of crustacean zooplankton in Qingcaosha Reservoir from 2015 to 2016 |
2015和2016年均各有优势种13种,但仅长额象鼻溞和汤匙华哲水蚤为两年的全年优势种,其优势度一直处于较高水平。由Pearson相关性分析可知,浮游甲壳类总密度的高低主要由重要优势种长额象鼻溞和汤匙华哲水蚤所决定,总密度和两个优势种的密度之间有极显著的正相关性(r=0.745,P<0.01)。2015年秋季优势种最多,有8种,春季最少仅有4种,长额象鼻溞、脆弱象鼻溞和汤匙华哲水蚤为4季共有优势种。2016年则是夏季优势种数最多,共有12种,其中长额象鼻溞占据最大优势,汤匙华哲水蚤次之,春季最少有4种。2015年冬季简弧象鼻溞优势度超过长额象鼻溞,而2016年秋季中华窄腹剑水蚤具有较大优势。

|
表 3 2015—2016年青草沙水库浮游甲壳类优势种的季节变化 Tab.3 Dominance of dominant crustacean zooplankton in different seasons in Qingcaosha Reservoir from 2015 to 2016 |
2015年浮游甲壳类的生物密度和生物量平均为(3.1±2.7)个/L和(0.208 3±0.244 6)mg/L,其生物密度平均值波动于1.5~7.7个/L,生物量则在0.006 3~0.460 0 mg/L之间。2016年水库内浮游甲壳类的生物密度和生物量平均值各自为(8.7±9.1)个/L与(0.598 3±0.692 6)mg/L,其中生物密度的波动幅度为2.0~21.2个/L,生物量在0.199 5~1.539 1 mg/L之间起伏。如图 2所示,2015年其平均生物密度在6月为最大,平均值为(7.7±4.9)个/L,次高峰在4月,平均生物密度为(4.0±3.6)个/L,根据年均生物量曲线可以看出2015年浮游甲壳类的生物量呈明显的双峰型,在6月份达到其生物量的最大值为(0.460 0±0.342 5)mg/L。2016年其密度最大值也是在6月(21.2±13.8)个/L,其次为5月(18.3±12.2)个/L,生物量走势为双峰型,6月份的平均生物量最大为(1.539 1±0.973 5) mg/L,3月的平均生物量次之,为(1.279 1±0.658 8) mg/L。综上所述,2015—2016两年内的最大现存量均出现在6月。

|
图 2 2015—2016年青草沙水库浮游甲壳类生物密度和生物量的月变化
Fig. 2 Annual variation of density and biomass of crustacean zooplankton in Qingcaosha Reservoir from 2015 to 2016
|
2015年浮游甲壳类的平均生物密度在冬夏季高于春秋季,生物量则为春夏季高于秋冬季,夏季的平均生物密度与生物量最高,分别为(4.6±3.8)个/L和(0.276 0±0.262 3)mg/L,秋季最低为(2.1±1.4)个/L和(0.060 1±0.108 3)mg/L;2016年浮游甲壳类的现存量表现出春夏季高于秋冬季的规律,春季其生物密度与生物量最高分别是(13.3±9.3)个/L和(1.145 7±0.771 5)mg/L,冬季最低分别为(2.5±1.8)个/L和(0.275 7±0.215 7)mg/L,2015年秋季现存量为两年内季节分布的最低水平(图 3)。利用最小显著差异法(LSD)对浮游甲壳类现存量进行季节间差异性分析,得出在季节间并没有显性著差异(P>0.05)。

|
图 3 2015—2016青草沙水库浮游甲壳类生物密度与生物量的季节变化
Fig. 3 Seasonal change of density and biomass of crustacean zooplankton in Qingcaosha Reservoir from 2015 to 2016
|
图 4所示,浮游甲壳类的平均生物密度在6号点最大,为(9.2±8.1)个/L,7号点次之(9.0±9.3)个/L,生物量在7号点最大6号点次之,分别为(0.685 6±0.747 3)mg/L和(0.638 8±0.648 8)mg/L,10号点的生物密度和生物量最低,分别为(1.1±1.0)个/L和(0.073 0±0.090 2)mg/L。S1、S10和S11站点位于水库的河流区,水流相对较大,并受长江来水的影响,其生物密度与生物量均处于最低水平。

|
图 4 青草沙水库2015—2016年浮游甲壳类现存量的水平分布
Fig. 4 Horizontal distribution of density and biomass of crustacean zooplankton in Qingcaosha Reservoir from 2015 to 2016
|
图 5显示,从河流区到过渡区再到湖泊区,2015—2016浮游甲壳类的生物密度与生物量均依次递增,2015年河流区的生物密度和生物量分别为(1.1±1.2)个/L和(0.075 0±0.115 2)mg/L,过渡区与湖泊区的生物密度分别为(3.5±2.9)个/L和(4.6±2.4)个/L,生物量分别为(0.225 1±0.231 6)mg/L和(0.313 7±0.300 0)mg/L,2016年湖泊区现存量在两年内达最高水平,生物密度与生物量平均为(13.0±10.8)个/L和(0.907 6±0.811 7)mg/L。利用SPSS软件对2015年不同区域浮游甲壳类的生物密度、生物量进行比较,得出河流区与过渡区、河流区与湖泊区之间在生物密度及生物量上均有极显著差异(P<0.01),2016年其生物密度与生物量在该3个区域内均有极显著差异(F=35.446,P=0.000;F=34.584,P=0.000)。

|
图 5 2015—2016青草沙水库浮游甲壳类生物密度与生物量的区域变化
Fig. 5 Area variation of crustacean zooplankton's density and biomass in Qingcaosha Reservoir from 2015 to 2016
|
枝角类的Shannon-Wiener物种多样性指数H′年平均值为0.61±0.34,Pielou物种均匀度指数J平均值为0.62±0.29;Margalef物种丰富度指数D平均值为0.57±0.29。桡足类这3个指数的均值分别为0.87±0.37、0.67±0.23和1.66±4.95。结合图 6,发现这3个指数在不同季节有所不同,枝角类和桡足类该3个指数的曲线走势基本一致,在夏季处于较高水平,枝角类指数值在秋季和冬季月份较小,而桡足类则在春季月份最低。

|
图 6 2015—2016年青草沙水库浮游甲壳类多样性指数的季节变化
Fig. 6 Seasonal change of diversity indexes of crustacean zooplankton in Qingcaosha Reservoir from 2015 to 2016
|

|
图 7 2015—2016年青草沙水库浮游甲壳类物种与环境因子的RDA排序图
Fig. 7 RDA ordination diagram between crustacean zooplankton species and environment factors in Qingcaosha Reservoir from 2015 to 2016
|
在此选择物种相对丰度大于1%且出现频率大于25%的浮游甲壳种类进行去趋势对应分析(即DCA分析),发现4个梯度值中最大的是2.009(小于3),所以运用RDA进一步对两年内浮游甲壳类生物密度与相应理化指标之间的相互作用进行分析。第一轴和第二轴的特征值分别为0.321、0.131,物种跟各因子间的相关性系数达0.957和0.926,因此可以利用RDA排序图来分析各理化指标对浮游甲壳类群落特征的影响,其累积变异百分数分别为32.1%和45.3%,物种环境累积变异百分数为50.8%和71.6%。从表 4中可以看出,与第一排序轴具有最大正负相关性的是亚硝态氮与水深,其相关性系数分别为0.753 2与-0.555 9,在第二轴中则是电导率(r=0.619 0)和总氮(r=-0.436 4)。

|
表 4 青草沙水库浮游甲壳类物种与环境因子RDA分析中前两个排序轴相关系数 Tab.4 Correlation coefficients between crustacean zooplankton and environment factors and the first two ordination axes of Qingcaosha Reservoir |
RDA排序图显示,对浮游甲壳类具有较大影响的环境因素有水温、硝态氮、亚硝态氮、高锰酸盐指数,青草沙水库中大多数浮游甲壳类与水温呈正相关关系,其中长额象鼻溞的相关性最大。多数种类与水深呈负相关,指状许水蚤受电导率的影响显著,pH、溶解氧、透明度和铵态氮对物种丰度的影响不显著,模式有爪猛水蚤、窄肢湖角猛水蚤和大尾真剑水蚤受各环境因素的影响甚小。主要优势种长额象鼻溞与水深、溶解氧、透明度有显著负相关关系,与其他各因子均具有显著性正相关关系;而汤匙华哲水蚤与总氮、总磷及水温具有负相关关系,与其他各因素有正相关关系。
3 讨论 3.1 青草沙水库浮游甲壳类种类组成特征两年共鉴定出枝角类18种,与广东省大沙河水库[21]、宁波横山水库[22]、湖北省三道河水库[23]相比物种相对丰富。青草沙水库有比较多的广温性枝角类种类,如低额溞属(Simocephalus)、尖额溞属(Alona)、盘肠溞属(Chydorus)的种类等。近几年每年的12月至下一年的4月份就会投放一些鱼类、环棱螺等对水环境进行控制与改善;滤食性的鲢(Hypophthalmichthys molitrix)、鳙(Aristichthys nobilis)鱼占主要组成部分,而浮游甲壳类是它们的主要食物,其有选择地捕食对枝角类的体型大小和现存量有明显影响[24],因而导致水库内枝角类的小型化。青草沙水库内枝角类的年均密度为(2.7±4.6)个/L,低于上海世博园后滩湿地的年均值[25],且远远低于富营养型湖泊淀山湖的枝角类平均生物密度(71个/L)[26],说明青草沙水库水体的营养水平较低。
两年内共鉴定出桡足类16种,与湖北金沙河水库[27]、安徽省太平湖水库[28]相比物种丰富,其中指状许水蚤、汤匙华哲水蚤、大尾真剑水蚤等属于广温性种类。哲水蚤目(Calanoida)往往构成寡营养水体中的最大优势种[29-31],而本研究显示汤匙华哲水蚤为两年中全年优势种且优势度较大,由此也可以说明青草沙水库营养水平较低。桡足幼体与无节幼体可长年观测到,并且其数量众多,这可能是由于水库内的滤食性鱼类先捕食大个体动物导致的,因为幼体透明度较高,不易被鱼类所发现[32]。
3.2 影响浮游甲壳类的环境因子浮游甲壳类的群落结构特征不仅受到其自身生长发育规律的影响,而且会受其生存环境中各因素的制约,经冗余分析显示:水温、硝态氮、亚硝态氮、高锰酸盐指数对浮游甲壳类影响较明显。本研究中青草沙水库的浮游甲壳类现存量表现出春夏季较高,由此说明,使群落结构改变的主要原因是温度的升降。有研究指出,当温度低于10 ℃时很少或基本没有广布中剑水蚤,当温度大于20 ℃时,广布中剑水蚤会快速繁殖,成为水体中的优势种类[33],这与本研究中广布中剑水蚤主要在夏秋季成为优势种的结果相吻合。浮游动物种类组成、生物密度及生物量的变动与水体的营养状态也有关[34],长额象鼻溞与总氮、总磷等营养盐参数呈正相关关系,长额象鼻溞在寡污到富营养型水体中均能成为优势种[35],在该水库中是两年内的全年优势种。
相关研究表明,水的流动速度是水体中浮游性生物生存与分布的关键因子,当流动速度很大时就不利于其生存[36-37],在水平分布上,浮游甲壳类的生物密度与生物量均呈现河流区<过渡区<湖泊区的规律,这与河流区(上游进水口)水流较大,深度较小,水体受搅动较大,使得浮游甲壳类的生存状况不佳,依次到过渡区与湖泊区,水位不断增加,水流速度减缓,并且水体不与外界进行交换,受人为影响小,使其在水中的滞留时间延长,导致其生物密度与生物量均依次递增。综上所述,青草沙水库浮游甲壳类群落主要受到水体温度、营养水平及流速的影响。
| [1] |
周超, 高乃云, 赵世嘏, 等. 上海青草沙水库水质调查与评价[J]. 同济大学学报(自然科学版), 2012, 40(6): 894-899. ZHOU C, GAO N Y, ZHAO S G, et al. Evaluation of water quality in Qingcaosha reservoir of Shanghai[J]. Journal of Tongji University (Natural Science), 2012, 40(6): 894-899. DOI:10.3969/j.issn.0253-374x.2012.06.016 |
| [2] |
LAMPERT W. Zooplankton research: the contribution of limnology to general ecological paradigms[J]. Aquatic Ecology, 1997, 31(1): 19-27. DOI:10.1023/A:1009943402621 |
| [3] |
YANG W, DENG D G, ZHANG S, et al. Seasonal dynamics of crustacean zooplankton community structure in Erhai Lake, a plateau lake, with reference to phytoplankton and environmental factors[J]. Chinese Journal of Oceanology and Limnology, 2014, 32(5): 1074-1082. DOI:10.1007/s00343-014-3204-5 |
| [4] |
AUER B, ELZER U, ARNDT H. Comparison of pelagic food webs in lakes along a trophic gradient and with seasonal aspects: influence of resource and predation[J]. Journal of Plankton Research, 2004, 26(6): 697-709. DOI:10.1093/plankt/fbh058 |
| [5] |
冯坤, 万成炎, 彭建华, 等. 三峡库区26条支流浮游甲壳动物的群落结构[J]. 水生态学杂志, 2012, 33(4): 46-48. FENG K, WAN C Y, PENG J H, et al. The crustacean zooplankton community structure in 26 tributaries of Three Gorges reservoir area[J]. Journal of Hydroecology, 2012, 33(4): 46-48. |
| [6] |
杨桂军, 潘宏凯, 刘正文, 等. 太湖不同湖区浮游甲壳动物季节变化的比较[J]. 中国环境科学, 2008, 28(1): 27-32. YANG G J, PAN H K, LIU Z W, et al. A comparative study on seasonal variations of crustaceans in the different lake areas in Lake Taihu[J]. China Environmental Science, 2008, 28(1): 27-32. DOI:10.3321/j.issn:1000-6923.2008.01.007 |
| [7] |
刘歆璞, 王丽卿, 张宁, 等. 青草沙水库后生浮游动物群落结构及其与环境因子的关系[J]. 生态学杂志, 2013, 32(5): 1238-1248. LIU X P, WANG L Q, ZAHNG N, et al. Community structure of metazoan zooplankton and its relationships with environmental factors in Qingcaosha Reservoir of Shanghai, East China[J]. Chinese Journal of Ecology, 2013, 32(5): 1238-1248. |
| [8] |
孙陆宇, 温晓蔓, 禹娜, 等. 上海青草沙水库后生浮游动物群落结构[J]. 复旦学报(自然科学版), 2013, 52(1): 119-128. SUN L Y, WEN X M, YU N, et al. Community structure of metazoan zooplankton in Qingcaosha Reservoir, Shanghai[J]. Journal of Fudan University (Natural Science), 2013, 52(1): 119-128. |
| [9] |
王少坤, 王绍祥, 顾静, 等. 上海饮用水源地青草沙水库轮虫群落结构的特征及其环境影响因子[J]. 动物学杂志, 2017, 52(6): 1023-1036. WANG S K, WANG S X, GU J, et al. The characteristics of community structure of rotifer and its environmental factors in Qingcaosha Reservoir, Shanghai[J]. Chinese Journal of Zoology, 2017, 52(6): 1023-1036. |
| [10] |
严文逸, 王绍祥, 顾静, 等. 青草沙水库浮游藻类群落组成及其与环境因子的关系[J]. 上海海洋大学学报, 2017, 26(1): 75-84. YAN W Y, WANG S X, GU J, et al. Composition of planktonic algae community and its relationship with environmental factors in Qingcaosha Reservoir[J]. Journal of Shanghai Ocean University, 2017, 26(1): 75-84. |
| [11] |
汤勇, 王绍祥, 陈立婧. 青草沙水库大型底栖动物群落结构及水质生物学评价[J]. 上海海洋大学学报, 2016, 25(6): 873-883. TANG Y, WANG S X, CHEN L J. Macrozoobenthic community structure and bioassessment for water quality of Qingcaosha Reservoir[J]. Journal of Shanghai Ocean University, 2016, 25(6): 873-883. |
| [12] |
陈伟民, 黄祥飞, 周万平. 湖泊生态系统观测方法[M]. 北京: 中国环境科学出版社, 2005: 23-35. CHEN W M, HUANG X F, ZHOU W P. Lake ecosystem observation method[M]. Beijing: China Environmental Science Press, 2005: 23-35. |
| [13] |
沈嘉瑞. 中国动物志-节肢动物门甲壳纲淡水桡足类[M]. 北京: 科学出版社, 1979. SHEN J R. Chinese zoology-arthropoda crustacea freshwater copepoda[M]. Beijing: Science Press, 1979. |
| [14] |
蒋燮治, 堵南山. 中国动物志-节肢动物门甲壳纲淡水枝角类[M]. 北京: 科学出版社, 1979. JIANG X Z, DU N S. Chinese zoology-arthropoda crustacea freshwater cladocera[M]. Beijing: Science Press, 1979. |
| [15] |
《浙江省主要觉见淡水浮游动物图集:饮用水水源》编委会. 浙江省主要常见淡水浮游动物图集(饮用水水源)[M]. 北京: 中国环境出版社, 2013: 8. Zhejiang Province is Mainly Aware of Atlas of Freshwater Zooplankton: Editorial Board of Drinking Water Sources. The main common freshwater zooplankton in Zhejiang Province[M]. Beijing: China Environmental Science Press, 2013: 8. |
| [16] |
黄祥飞, 胡春英.淡水常见枝角类体长-体重回归方程式[M]//甲壳动物学论文集编委会编.甲壳动物学论文集, 北京: 科学出版社, 1986: 147-157. HUANG X F, HU C Y. Freshwater common Cladocera body length-weight regression equation[M]//Editorial Committee of Crustaceans. Crustacean Papers. Beijing: Science Press, 1986: 147-157. |
| [17] |
陈亚瞿, 徐兆礼, 王云龙, 等. 长江口河口锋区浮游动物生态研究Ⅰ生物量及优势种的平面分布[J]. 中国水产科学, 1995, 2(1): 49-58. CHEN Y Q, XU Z L, WANG Y L, et al. An ecological study on zooplankton in plume front zone of changjiang (Yangtze) river estuarine area Ⅰ biomass distribution of dominant species[J]. Journal of Fishery Sciences of China, 1995, 2(1): 49-58. DOI:10.3321/j.issn:1005-8737.1995.01.003 |
| [18] |
SHANNON C E, WEAVER W. The mathematical theory of communication[M]. Urbana: University of Illinois Press, 1949: 623-656.
|
| [19] |
MARGALEF R. Information theory in ecology[J]. General Systems, 1958, 3: 36-71. |
| [20] |
李共国, 虞左明. 千岛湖轮虫群落结构及水质生态学评价[J]. 湖泊科学, 2003, 15(2): 169-176. LI G G, YU Z M. Community structure of rotifera and ecological assessment of water quality in Qiandao Lake[J]. Journal of Lake Sciences, 2003, 15(2): 169-176. DOI:10.3321/j.issn:1003-5427.2003.02.011 |
| [21] |
郭菲菲, 李慧明, 顾杨亮, 等. 大沙河水库浮游甲壳类动物群落特征[J]. 生态科学, 2015, 34(5): 8-15. GUO F F, LI H M, GU Y L, et al. Community structure of planktonic crustaceans in large, tropical, eutrophic Dashahe reservoir, South China[J]. Ecological Science, 2015, 34(5): 8-15. |
| [22] |
杨亮杰, 吕光汉, 竺俊全, 等. 横山水库浮游动物群落结构特征及水质评价[J]. 水生生物学报, 2014, 38(4): 720-728. YANG L J, LÜ G H, ZHU J Q, et al. Characteristics of zooplankton community in Hengshan Reservoir and water quality assessment[J]. Acta Hydrobiologica Sinica, 2014, 38(4): 720-728. |
| [23] |
冯坤, 彭建华, 万成炎, 等. 三道河水库浮游甲壳动物群落结构特征的初步研究[J]. 水生态学杂志, 2010, 3(3): 18-22. FENG K, PENG J H, WAN C Y, et al. Studies on the community structure characteristics of the microcrustaceans in Sandaohe Reservoir[J]. Journal of Hydroecology, 2010, 3(3): 18-22. |
| [24] |
MACIEJ GLIWICZ Z. On the different nature of top-down and bottom-up effects in pelagic food webs[J]. Freshwater Biology, 2002, 47(12): 2296-2312. DOI:10.1046/j.1365-2427.2002.00990.x |
| [25] |
陈立婧, 吴艳芳, 景钰湘, 等. 上海世博园后滩湿地枝角类群落结构的周年动态[J]. 应用生态学报, 2012, 23(10): 2863-2870. CHEN L J, WU Y F, JING Y X, et al. Annual dynamics of cladocera community structure in backshore wetland of Expo Garden, Shanghai[J]. Chinese Journal of Applied Ecology, 2012, 23(10): 2863-2870. |
| [26] |
郑小燕, 王丽卿, 盖建军, 等. 淀山湖浮游动物的群落结构及动态[J]. 动物学杂志, 2009, 44(5): 78-85. ZHENG X Y, WANG L Q, GAI J J, et al. Dynamic and community structure of zooplankton in Dianshan Lake[J]. Chinese Journal of Zoology, 2009, 44(5): 78-85. |
| [27] |
郭飞飞, 张云, 赵广, 等. 金沙河水库浮游动物群落结构及其与环境因子的关系[J]. 生态学杂志, 2016, 35(8): 2208-2116. GUO F F, ZHANG Y, ZHAO G, et al. Community structure of zooplankton in the Jinshahe Reservoir and its relationship with environmental factors[J]. Chinese Journal of Ecology, 2016, 35(8): 2208-2116. |
| [28] |
吴明姝.安徽省太平湖水库浮游动物群落结构及水质评价[D].上海: 上海师范大学, 2015. WU M S. Zooplankton community structure and water quality analysis of Taiping lake in Anhui Province[D]. Shanghai: Shanghai Normal University, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10270-1015371660.htm |
| [29] |
林秋奇, 胡韧, 段舜山, 等. 广东省大中型供水水库营养现状及浮游生物的响应[J]. 生态学报, 2003, 23(6): 1101-1108. LIN Q Q, HU R, DUAN S S, et al. Reservoir trophic states and the response of plankton in Guangdong Province[J]. Acta Ecologica Sinica, 2003, 23(6): 1101-1108. DOI:10.3321/j.issn:1000-0933.2003.06.010 |
| [30] |
REYNOLDS C S. What factors influence the species composition of phytoplankton in lakes of different trophic status[J]. Hydrobiologia, 1998, 369-370: 11-26. DOI:10.1023/A:1017062213207 |
| [31] |
RAWSON D S. Algal indicators of Trophic Lake types[J]. Limnology and Oceanography, 1956, 1(1): 18-25. DOI:10.4319/lo.1956.1.1.0018 |
| [32] |
YANG Y F, HUANG X F, LIU J K, et al. Effects of fish stocking on the zooplankton community structure in a shallow lake in China[J]. Fisheries Management and Ecology, 2005, 12(2): 81-89. DOI:10.1111/fme.2005.12.issue-2 |
| [33] |
陈受忠. 武昌东湖桡足类数量的周年资料[J]. 水生生物学集刊, 1965, 5(2): 202-219. CHEN S Z. Anniversary information on the number of copepods in East Lake of Wuchang[J]. Aquatic Biology Journal, 1965, 5(2): 202-219. |
| [34] |
侯来利.淮南市高塘湖水域浮游动物群落特征研究[D].淮南: 安徽理工大学, 2007. HOU L L. Study on the community characteristics of zooplankton of Gaotang Lake in Huainan[D]. Huainan: Anhui University of Science & Technology, 2007. http://cdmd.cnki.com.cn/Article/CDMD-10361-2007127872.htm |
| [35] |
陈绵润, 赵帅营, 林秋奇, 等. 广东省水库枝角类组成特征的初步研究[J]. 湖泊科学, 2007, 19(1): 77-86. CHEN M R, ZHAO S Y, LIN Q Q, et al. Cladoceran distribution in reservoirs of Guangdong Province, South China[J]. Journal of Lake Sciences, 2007, 19(1): 77-86. DOI:10.3321/j.issn:1003-5427.2007.01.012 |
| [36] |
陈受忠. 四川沱江口浮游甲壳动物记述[J]. 海洋湖沼通报, 1990(3): 86-91. CHEN S Z. Microcrustaceans at the mouth of Tuojiang River[J]. Transactions of Oceanology and Limnology, 1990(3): 86-91. |
| [37] |
向贤芬, 陈受忠, 曹文宣. 汉江中下游春季浮游甲壳动物群落结构的空间差异[J]. 长江流域资源与环境, 2004, 13(2): 187-192. XIANG X F, CHEN S Z, CAO W X. Spatial variations of planktonic crustacea communities in the middle and lower Hanjiang River[J]. Resources and Environment in the Yangtze Basin, 2004, 13(2): 187-192. DOI:10.3969/j.issn.1004-8227.2004.02.018 |
2. Key Laboratory ofFreshwater Aquatic Genetic Resources, Ministry ofAgriculture, Shanghai Ocean University, Shanghai 201306, China;
3. Shanghai Collaborative Innovation for Aquatic Animal Genetics and Breeding, Shanghai Ocean University, Shanghai 201306, China;
4. Qingcaosha Reservoir Management Branch, Shanghai Smi Raw Water Co. Ltd., Shanghai 201913, China