 2018,
Vol. 27
2018,
Vol. 27



有明银鱼(Salanx ariakensis)隶属胡瓜鱼目(Osmeriformes),胡瓜鱼科(Osmeridae),银鱼属(Salanx)。为沿海港湾及河口半咸淡水域的小型鱼类,栖息于沿海河口附近的中上层水域,分布于中国沿海、也见于朝鲜半岛和日本九州西部沿岸[1]。现阶段,国内已有的研究主要集中在有明银鱼的基础生物学特征[2-3]、耳石结构[4]、遗传分析[5]、仔鱼资源分布[6]等方面,国外则多见遗传检测[7]、基础形态学[8-9]等报道。国内外关于有明银鱼骨骼系统发育的研究,仅MIZUTAN[10]描述了仔稚鱼的附肢骨骼的发育出现时间及其完成时间与支鳍骨数目,缺乏有明银鱼骨骼系统在仔稚鱼阶段的形态学研究。本研究在探明杭州湾北部有明银鱼时空分布的基础上,利用硬骨-软骨双染色技术,对有明银鱼早期发育过程中脊柱与附肢骨骼的发育及其变化进行了观察和描述,旨在为有明银鱼仔稚鱼的分类与鉴定提供基础的科学依据。
1 材料与方法 1.1 调查方法2014年6月至2015年5月,在杭州湾北部水域设置10个采样点(St.1-St.10, 图 1)。每月大潮前后使用大型仔稚鱼网(口径1.3 m,网目0.5 mm)进行表层水平拖网。拖速为2海里/h。使用网口流量计(KC Denmark A/S Model 23.090)记录拖网流量值。采集到的标本用5%甲醛溶液固定。在所有样本中筛选出有明银鱼,参照KENDALL[11]的仔稚鱼发育分期标准划分发育阶段,计数并计算密度(尾/m3)。前弯曲期和弯曲期仔鱼测量脊索长(notochord length, NL),后弯曲期仔鱼测量标准体长(standard length, SL)。

|
图 1 调查站位图
Fig. 1 Location of sampling stations
|
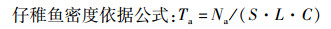 (1)
(1)
式中: Ta为单位体积海水中仔稚鱼个体数(尾/m3);Na为每网采集到的仔稚鱼个体数(尾);S为网口面积(m2);L为流量计转数;C为流量计校正值(C=0.3)[12]。
1.2 双染色透明样本处理参考DINGERKUS和UHLER[13]的软骨-硬骨双染色透明技术进行研究,(1)清洗:使用蒸馏水清洗用5%甲醛溶液固定的有明银鱼仔稚鱼标本,浸泡2~3 d,中间多次更换蒸馏水;(2)软骨染色:将仔稚鱼标本浸泡于软骨染色液中24~36 h,直至标本鳍条基部出现明显的蓝色;(3)梯度漂洗:先将标本取出并移入95%乙醇溶液中浸泡2~3 h,随后依次移入呈递减浓度(75%、40%、15%)的乙醇中直至标本完全沉底后再换入下一浓度梯度;(4)组织的消化与透明:将标本浸于0.5%胰蛋白酶消化液中,直至标本在消化液中完全软化且透明;(5)硬骨染色:将消化透明后的标本移入硬骨染色液中24~48 h,直至硬骨染成红紫色为止;(6)脱色:经过染色处理的样品放入清水中,并加入1~2滴H2O2,放置于阳光下褪色;(7)保存:将标本依次移入体积比为3:1、1:1、1:3的0.5%KOH-甘油混合试剂中,每一梯度至标本完全沉底转移入下一梯度,最终将标本转移入纯甘油中密封保存,并加入几粒百里酚密封保存。在Olympus SZX7解剖镜下,对有明银鱼的骨骼形态进行连续观察并绘制发育图。并使用Photoshop和Adobe Illustrator软件对骨骼发育图进行编辑处理。
2 结果 2.1 时空分布2014年6月至2015年5月共采集到鱼类仔稚鱼96 118尾,其中有明银鱼仔稚鱼1 972尾,占总渔获量的2.04%。有明银鱼仔稚鱼仅出现于2014年10月至2015年1月,平均密度呈先增加后下降的趋势,11月有明银鱼数量最多,平均密度为6.06尾/100 m3(表 1)。各站位点均有有明银鱼分布,St.3平均密度最高(2.59尾/100 m3),St.7平均密度最低(0.52尾/100 m3)。前弯曲期仔鱼主要分布在靠近岸边的St.9和St.10,弯曲期仔鱼主要分布在靠近岸边的St.8和St.9,后弯曲期仔鱼主要分布在远离岸边的St.6,稚鱼主要集中于东海大桥两侧的St.3和St.4(图 2)。从仔稚鱼的发育阶段来看,10月以前弯曲期仔鱼为主(63.42%),11月以弯曲期仔鱼为主(47.07%)、后弯曲期仔鱼和稚鱼所占比例上升。12月和1月以稚鱼为主(图 3)。

|
表 1 有明银鱼仔稚鱼个体数与平均密度 Tab.1 Quantity and mean density of S. ariakensis larvae and juveniles |

|
图 2 杭州湾北部水域有明银鱼仔稚鱼的密度分布
Fig. 2 Density distribution of S. ariakensis larvae and juveniles in the north of Hangzhou Bay
|

|
图 3 各月有明银鱼仔稚鱼发育阶段组成
Fig. 3 Development stages composition of S. ariakensis larvae and juveniles in different months
|
有明银鱼前弯曲期仔鱼体长范围为4.55~12.56 mm。弯曲期仔鱼体长范围为11.47~19.28 mm,后弯曲期仔鱼体长范围为18.20~24.14 mm,稚鱼体长范围为24.13~49.70 mm。优势体长组是10~20 mm,占有明银鱼总捕获数的47.76%。
2.3 脊柱发育体长(下略)9.7 mm个体的脊索粗大呈管状(图 4a)。13.77 mm个体脊索末端已开始向上弯曲,脊索仍呈管状(图 4b)。13.94 mm,16.08 mm,15.99 mm,14.88 mm个体脊索无明显变化(图 4c-f)。22.81 mm个体出现18个脉弓与19个髓弓(图 4g)。脉弓起点位于臀鳍的第3~4枚支鳍骨间上方,后至脊索末端。髓弓与脉弓的位置相对,尾杆骨的上边缘开始骨化。28.93 mm个体出现扁平的脉棘和髓棘,尤其是前者几近薄片状,髓弓71个贯穿整个主轴骨骼,脉弓21个,脉弓起点已超过臀鳍前缘。整个尾杆骨呈红色,已骨化完全。除尾杆骨之外的脊柱部分即脊索仍未钙化,在髓弓和脉弓的基础上,髓棘和脉棘以软骨形式开始出现,但未见到背肋、腹肋(图 4h)。34.2 mm个体椎体仍不明显,髓弓和脉弓、髓棘和脉棘完全形成,髓弓73个,脉弓22个(图 4i)。髓弓、脉弓首先以软骨组织形式出现,并整体由脊椎尾端向头部发生,整个发生过程中髓弓与脉弓几乎同时出现,生长速度相对平行。脊柱从头至尾、从先至后也均未出现分节的硬骨环,椎弓不明显。

|
图 4 有明银鱼主轴和附肢骨骼发育示意图
Fig. 4 Development of the vertebral column and the appendicular skeleton of S. ariakensis
(a)体长9.7 mm; (b)体长13.77 mm; (c)体长13.94 mm; (d)体长16.08 mm; (e)体长15.99 mm; (f)体长14.88 mm; (g)体长22.81 mm; (h)体长28.93 mm; (i)体长34.2 mm; AnPt.臀鳍支鳍骨; DoPt.背鳍支鳍骨; EP.尾上骨; HA.脉弓; HS.脉棘; HY.尾下骨; NA.髓弓; NC.脊索; NS.髓棘; PH.侧尾下骨; UR.尾杆骨; UrN.尾椎髓体; 粗点区域表示软骨; 细点区域表示硬骨(尾杆骨); 空白区域表示膜质骨(脊索)、硬骨(支鳍骨、尾鳍鳍条)
(a)9.7 mm NL; (b)13.77 mm NL; (c)13.94 mm NL; (d)16.08 mm NL; (e)15.99 mm NL; (f)14.88 mm NL; (g)22.81 mm SL; (h)28.93 mm SL; (i)34.2 mm SL; AnPt. pterygiophore of anal; DoPt. pterygiophore of dorsal; EP. epural; HA. haemal arch; HS. haemal spine; HY. hypural; NA. neural arch; NC. notochord; NS. neural spine; PH. parhypural; UR. urostyle; UrN. uroneural; big point areas were cartilage; small point areas were bone in urostyle; blank areas were notochord, bone in pterygiophore |
9.7 mm个体尾鳍以鳍褶形式出现,未见软骨组织(图 5a)。13.77 mm个体最早出现2枚尾下骨,即尾下骨2-3(图 5b)。13.94 mm个体有4枚尾下骨,即尾下骨1-4(图 5c)。16.08 mm个体有4枚尾下骨,同时尾索下部鳍皱演变成放射丝,着生在尾下骨上,尾索上部仍然是鳍褶(图 5d)。14.88mm和15.99 mm个体出现侧尾下骨1枚、尾下骨5枚(图 5e-f)。22.81 mm个体尾杆骨上边缘变红开始钙化,仍保持1枚侧尾下骨、5枚尾下骨,尾下骨1的中部轻微钙化。出现较为明显的上、下叶之分。其中,下叶由侧尾下骨和尾下骨1-2组成,上叶由3-5组成(图 5g)。28.93 mm个体的尾杆骨基本与脊索垂直且除基部外均已钙化完全。出现6枚尾下骨,侧尾下骨、第1尾下骨与第2尾下骨三者末端呈现愈合趋势,第1尾下骨与第2尾下骨的基部完全愈合。6枚尾下骨的中间大部分硬骨化,尾杆骨上方出现1枚软骨质的尾上骨(图 5h)。34.2 mm个体尾鳍骨骼基本成型,第3尾下骨与第4尾下骨的末端也呈现出愈合趋势。第5和6尾下骨游离。尾上骨仍为1枚。尾杆骨完全硬化,其上方出现1枚尾椎髓体(uroneural)。下叶的侧尾下骨和尾下骨1-2基部与末端还未骨化,中间大部分已骨化;上叶的尾下骨3-5除末端外都已骨化。尾下骨6已骨化完全(图 5i)。

|
图 5 有明银鱼尾鳍支鳍骨发育
Fig. 5 Development of caudal fin in S. ariakensis
(a)体长9.7 mm; (b)体长13.77 mm; (c)体长13.94 mm; (d)体长16.08 mm; (e)体长15.99 mm; (f)体长14.88 mm; (g)体长22.81 mm; (h)体长28.93 mm; (i)体长34.2 mm; EP.尾上骨; HY.尾下骨; PH.侧尾下骨; UR.尾杆骨; UrN.尾椎髓体; 粗点区域表示软骨; 细点区域表示硬骨(尾杆骨); 空白区域表示膜质骨(脊索)、硬骨(支鳍骨)
(a)9.7 mm NL; (b)13.77 mm NL; (c)13.94 mm NL; (d)16.08 mm NL; (e)15.99 mm NL; (f)14.88 mm NL; (g)22.81 mm SL; (h)28.93 mm SL; (i)34.2 mm SL; EP. epural; HY. hypural; PH. parhypural; UR. urostyle; UrN. uroneural; big point areas were cartilage; small point areas were bone in urostyle; blank areas were notochord, bone in pterygiophore |
9.7 mm个体背鳍以鳍褶形式出现(图 4a)。16.08 mm个体脊索中部出现6枚的背鳍支鳍骨(图 4d)。随着时间的推移,背鳍支鳍骨向前移动、数量增多、形态加粗,15.99 mm个体出现11枚支鳍骨,此时背鳍仍是鳍褶状(图 4e)。14.88 mm个体支鳍骨为11枚,但鳍褶已演变成放射丝(图 4f)。22.81 mm SL个体开始出现担鳍骨,此时背鳍支鳍骨、担鳍骨数增至13枚;并形成软骨质鳍条(图 4g)。28.93 mm个体背鳍支鳍骨、担鳍骨数增至13枚,仍为软骨质(图 4h)。34.2 mm个体时背鳍支鳍骨、担鳍骨数稳定在13枚(图 4i)。
2.4.3 臀鳍13.77 mm个体在肛门后端出现7枚软骨质的臀鳍支鳍骨(图 4b)。13.94 mm个体出现11枚臀鳍支鳍骨(图 4c),16.08 mm个体具15枚(图 4d),15.99 mm个体16枚(图 4e),14.88 mm个体19枚(图 4f)。支鳍骨均为软骨,其末端发育出放射性的鳍丝。22.81 mm个体出现担鳍骨,此时臀鳍的支鳍骨和担鳍骨均为软骨,数目为26枚,支鳍骨末端发育出1节鳍条,其前缘部分开始硬骨骨化。臀鳍支鳍骨的第一枚与最后一枚,其上均生发有2枚鳍条。共有26枚支鳍骨,28枚鳍条(图 4g)。28.93 mm个体臀鳍担鳍骨开始硬骨化,支鳍骨没有变化。臀鳍的支鳍骨和担鳍骨数仍为26枚(图 4h)。34.2 mm个体臀鳍担鳍骨与鳍条已完全硬骨化,支鳍骨仍保持软骨。支鳍骨和担鳍骨数保持稳定为27枚(图 4i)。
2.4.4 胸鳍胸鳍是有明银鱼最早出现的鳍。9.7 mm个体胸鳍有软骨质的匙骨、乌喙骨和支鳍骨原基,匙骨呈长条状(图 6a)。16.08 mm个体匙骨上下两端延长,从中部开始骨化,出现上匙骨和后颞骨(图 6b)。22.81 mm个体匙骨与上匙骨已经骨化。支鳍骨原基出现3个裂缝,形成两个分支,无后匙骨(图 6c)。28.93 mm个体乌喙骨上出现一个孔,后颞骨开始骨化(图 6d)。34.2 mm个体乌喙骨上出现第二个孔,后颞骨已经骨化,支鳍骨原基出现5个裂缝,形成3个分支,胸鳍鳍条尚未开始分化(图 6e)。36.77 mm个体胸鳍鳍条开始分化。

|
图 6 有明银鱼肩带发育
Fig. 6 Development of pectoral girdle in S. ariakensis
(a)体长9.7 mm; (b)体长16.08 mm; (c)体长22.81 mm; (d)体长28.93 mm; (e)体长34.2 mm; Cl.匙骨; Fp.支鳍骨原基; Co.乌喙骨; Suc.上匙骨; Pot.后颞骨.细点区域表示硬骨; 空白区域表示软骨 (a)9.7 mm NL; (b)16.08 mm NL; (c)22.81 mm SL; (d)28.93 mm SL; (e)34.2 mm SL; Cl. cleithrum; Co. coracoid; Fp. fin plate; Pot. posttemporal; Suc. supracleithrum; small point areas were bone; blank areas were cartilage |
22.81 mm个体未发现腹鳍原基。28.93 mm个体在肛门前方可见软骨质腹鳍支鳍骨,呈棒状,末端具5枚软骨质鳍条。34.2 mm个体腹鳍支鳍骨仍为软骨,软骨质鳍条数目增加为6。
杭州湾有明银鱼各鳍发育顺序依次为:胸鳍最早出现,尾鳍与臀鳍几乎同时出现,背鳍随后出现,腹鳍出现较晚,49.7 mm个体出现脂鳍。
3 讨论 3.1 与日本有明海产有明银鱼比较杭州湾北部13.77 mm个体出现2枚尾下骨;而日本有明海[10]产12.8 mm个体未出现尾下骨,15.2 mm个体出现尾下骨。杭州湾北部的有明银鱼尾下骨可能比有明海产的有明银鱼发育更早。杭州湾北部有明银鱼尾鳍条组成为上叶10枚鳍条、下叶9枚鳍条,与有明海产有明银鱼相同。杭州湾北部的13.77 mm个体在肛门后端出现7枚软骨质的臀鳍支鳍骨,34.2 mm个体臀鳍发育完全,鳍条数量达到恒定的27。有明海产12.8 mm个体臀鳍支鳍骨出现;23.5 mm个体臀鳍发育完全,鳍条数量为29。鳍条数目的不同可能受鳍条统计方法不同有关。杭州湾产16.08 mm个体出现6枚软骨质的背鳍支鳍骨,22.81 mm个体开始出现担鳍骨,此时背鳍支鳍骨、担鳍骨数增至13枚。有明海产14.5 mm个体形成背鳍原基,23.5 mm个体鳍条数成为恒定的13枚。本研究所用的杭州湾有明银鱼的胸鳍和腹鳍鳍条未发育完全。而有明海产38.9 mm个体可明显观察到胸鳍鳍条,44.1 mm个体胸鳍鳍条数量达到9枚。腹鳍在后弯曲期(23.5 mm)的第28至29个肌节间出现原基,38.9 mm个体可以明显观察到,44.1 mm个体腹鳍鳍条数目达到恒定的7枚,出现脂鳍。
3.2 与其他种的比较据ROBERTS[14]报道,大银鱼(Protosalanx chinensis)、居氏银鱼(Salanx cuvieri)、石川日本新银鱼(Neosalangichthys ishikawae)、乔氏新银鱼(Neosalanx jordani)第1-2髓弓末端愈合,脉弓和髓弓尤其是前者几近薄片状。有明银鱼的第1与第2髓弓间距相较于其他髓弓则较近,末端并未愈合(图 4i),脉弓和髓弓呈薄片状,尤其是脉弓十分平薄(图 4g~i)。林信伟和熊全沫[15]通过组织学方法发现寡齿新银鱼(Neosalanx oligodontis)也是在椎体位置仅存在一条脊索,呈节状。每节脊椎通过其外包围有3块软骨的脊索构成。本实验过程中的有明银鱼脊索位置并未观察到3块软骨的存在。但是考虑到ROBERTS、林信伟等所用样本都为成鱼,关于有明银鱼脊椎之后是否会发生变化需要进一步研究阐明。
ROBERTS[14]还指出银鱼的尾杆骨复合体由3个椎体、尾椎髓体1或2枚组成。但在有明银鱼中未见到3个已分化的椎体,仅在尾杆骨上方见到1枚已骨化的尾椎髓体(图 5i)。此外有明银鱼尾鳍的侧尾下骨、第1尾下骨与第2尾下骨三者的末端呈现愈合趋势,第1尾下骨与第2尾下骨的基部完全愈合。此外,第3尾下骨与第4尾下骨的末端也呈现出愈合趋势。第5和6尾下骨游离。然而,同一科中的小齿日本银鱼(Salangichthys microdon)尾鳍的6枚尾下骨则相互分离。杭州湾银鱼类除有明银鱼外,还有大银鱼[16]。大银鱼尾下骨1-2、尾下骨5-6游离,但是尾下骨3-4的末端愈合。在新银鱼属、银鱼属中,其尾下骨的愈合现象更多。其中,银鱼属的侧尾下骨与尾下骨1-2末端靠近呈愈合趋势;尾下骨1-2和3-4基本完全愈合,只在基部剩下一个椭圆形的空洞表明还未全部愈合。可见银鱼科鱼类的尾下骨愈合现象比较复杂,可能与其复杂、多样的生存环境有关。但是,从大多数已描述的银鱼类的尾下骨数目上来看,基本上都保持在6个尾下骨,这加强了这一事实:尾下骨数目在该物种中可能是高度保守的。
3.3 有明银鱼骨骼发育的特殊性在许多鱼类仔稚鱼骨骼系统的发育中,其形态结构都会出现显著的变化。其中脊椎的躯椎部分分化出圆柱形椎体,以椎体两端的关节顺次衔接。椎体两端向中部凹入,椎体两侧向上发出二片髓弓,其间形成髓管,上端合并为髓棘。髓弓前缘有前髓关节突,后方有后髓关节突;椎体两侧出现倾斜的长突起即椎体横突,与肋骨上端相关节[17]。然而,本研究通过硬骨-软骨双染色技术,观察有明银鱼仔稚鱼脊柱的形态结构始终呈现粗细相同的棒状结构;其染色效果则既非显著的红色,也非深蓝色,几乎透明无色略泛蓝。说明有明银鱼的脊柱在本研究个体中并未分化出椎体,而是本应完全退化或者部分残余但仍完全保留下来的脊索。推测由于外围的脊索鞘由纤维组织和弹性组织所构成,故对阿辛蓝和茜苏红都不呈显色反应。而且臀鳍、背鳍和尾鳍已基本发育完全时,脊柱上的脉弓与髓弓表现出蓝色的半月牙形,此后不再变化。说明同源性的脊柱以及其上的脉弓与髓弓在后弯曲期出现了差异:脊柱停滞在膜质期,形成仅具有膜质结构的脊索;脉弓与髓弓的发育则先后经历了膜质期与软骨期,至此具有明显的软骨结构。本研究结果与1925年,JORDAN在发现银鱼科鱼类的骨骼几乎没有骨化;1940年,BERG指出的银鱼类似乎是幼态持续的鱼类;以及1965年,伍献文在实际观察中发现的银鱼科鱼类在性成熟时期尚滞留在个体发育较前期或幼龄的形态,即“幼态持续”的结果相同[18]。
| [1] |
赵盛龙, 徐汉祥, 钟俊生, 等. 浙江海洋鱼类志[M]. 杭州: 浙江科学技术出版社, 2016: 255-256. ZHAO S L, XU H X, ZHONG J S, et al. Fauna of Marine Fishes in Zhejiang[M]. Hangzhou: Zhejiang Science and Technology Press, 2016: 255-256. |
| [2] |
孙帼英. 长江口及其邻近海域有明银鱼的生物学[J]. 海洋湖沼通报, 1990(1): 41-46. SUN G Y. Salanx ariakensis in Yangtze River Estuary and the Neighbouring Marine Water[J]. Transactions of Oceanology and Limnology, 1990(1): 41-46. |
| [3] |
唐作鹏, 解涵, 李勃, 等. 鸭绿江有明银鱼的生物学研究[J]. 大连海洋大学学报, 2000, 15(2): 113-118. TANG Z P, XIE H, LI B, et al. The biological aspects of Ariake icefish (Salanx ariakensis Kishinouye) in Yalujiang River[J]. Journal of Dalian Fisheries University, 2000, 15(2): 113-118. DOI:10.3969/j.issn.1000-9957.2000.02.007 |
| [4] |
解玉浩, 唐作鹏, 解涵, 等. 有明银鱼耳石显微结构和微化学研究[J]. 动物学报(Current Zoology), 2001, 47(2): 215-220. XIE Y H, TANG ZP, XIE H, et al. Microstructure And Microchemistry In Otolith Of Ariake Icefish (Salanx ariakensis)[J]. Acta Zoologica Sinica, 2001, 47(2): 215-220. |
| [5] |
张际峰, 郝培应, 聂刘旺, 等. 2种银鱼线粒体COⅡ及侧翼tRNA基因的测定分析及其亲缘关系研究[J]. 中国海洋大学学报(自然科学版), 2008, 38(3): 424-428. ZHANG J F, HAO P Y, NIE L W, et al. Studies on the Analysis of Sequences of CO Ⅱ and its Connected tRNA Genes of Mitochondrial Genome from 2 Species of Salangidae and Their Phylogenetic Relationship[J]. Periodical of Ocean University of China, 2008, 38(3): 424-428. |
| [6] |
宋超, 刘媛媛, 吕杨, 等. 长江口有明银鱼仔鱼的分布及其与环境因子的关系[J]. 海洋渔业, 2015, 37(4): 318-324. SONG C, LIU Y Y, LV Y, et al. Distribution of Salanx ariakensis larvae in the Yangtze Estuary and its relationship with environmental factors[J]. Marine Fisheries, 2015, 37(4): 318-324. DOI:10.3969/j.issn.1004-2490.2015.04.004 |
| [7] |
JIN K K, KAI Y, NAKABO T. Genetic diversity of Salanx ariakensis, (Salangidae) from Korea and Japan inferred from AFLP[J]. Ichthyological Research, 2007, 54(4): 416-419. DOI:10.1007/s10228-007-0418-y |
| [8] |
HIBINO M, KINOSHITA I, OHTA T, et al. Morphology of Ariake icefish Salanx ariakensis larvae in the Chikugo estuary[J]. Jpn J Ichthyol, 2002, 49: 103-108. |
| [9] |
JIN K K, DOIUCHI R, NAKABO T. Molecular and morphological differences between two geographic populations of Salanx ariakensis, (Salangidae) from Korea and Japan[J]. Ichthyological Research, 2006, 53(1): 52-62. DOI:10.1007/s10228-005-0315-1 |
| [10] |
MIZUTANI H, MATSUI S, TAKESHITA N. Embryonic, larval and juvenile development of Salanx ariakensis (Pisces:Salangidae)[J]. Suisanzoshoku, 2010, 48: 497-502. |
| [11] |
KENDALL JR A W. Early life history stages of fishes and their characters[M]//MOSER H G, RICHARDS W J, COHEN D M, et al. Ontogeny & Systematics of Fishes. The American Society of Ichthyologists Herpetologists. Lawrence: Allen Press Inc, 1984: 11-22
|
| [12] |
侯世挺, 钟俊生, 王晓东, 等. 长江口南支仔稚鱼数量随潮汐变化的研究[J]. 上海海洋大学学报, 2018, 27(2): 250-258. HOU S T, ZHONG J S, WANG X D, et al. The study on the amount changes of fish larvae during flood and ebb in the south branch of Yangtze River Estuary[J]. Journal of Shanghai Ocean University, 2018, 27(2): 250-258. |
| [13] |
DINGERKUS G, UHLER I D. Enzyme clearing of alcian blue stained whole small vertebrates for demonstration of cartilage[J]. Stain Technology, 1977, 52(4): 229-232. DOI:10.3109/10520297709116780 |
| [14] |
ROBERTS T R. Skeletal anatomy and classification of the neotenic Asian salmoniform superfamily Salangoidea (icefishes or noodlefishes)[J]. Proceedings of the California Academy of Sciences, 1984, 43(13): 179-220. |
| [15] |
林信伟, 熊全沫. 寡齿新银鱼骨骼系统的形态和组织学特征[J]. 动物学报(Current Zoology), 1994(2): 198-199. LIN X W, XIONG Q M. Morphological and Histological Characteristics of Skeletal System of Neosalanx oligodontis Chen[J]. Acta zoologica Sinica, 1994(2): 198-199. |
| [16] |
胡敏, 钟俊生, 唐子涵, 等. 杭州湾北部水域仔稚鱼组成的月度及年间比较[J]. 上海海洋大学学报, 2017, 26(1): 103-112. HU M, ZHONG J S, TANG Z H, et al. Monthly and annual comparison of fish larvae and juveniles composition in the north of Hangzhou Bay[J]. Journal of Shanghai Ocean University, 2017, 26(1): 103-112. |
| [17] |
孟庆闻, 苏锦祥, 李婉端. 鱼类比较解剖[M]. 科学出版社, 1987: 99-102. MENG Q W, SU J X, LI W R. Comparative anatomy of fishes[M]. Science Press, 1987: 99-102. |
| [18] |
胡传林, 刘家寿, 彭建华, 等. 我国银鱼研究概况及其移植的生态管理准则[J]. 湖泊科学, 2001, 13(3): 204-210. HU C L, LIU J S, PENG J H, et al. Salangids in China and Their Ecological Management Principles for Transplantation[J]. Journal of Lake Sciences, 2001, 13(3): 204-210. DOI:10.3321/j.issn:1003-5427.2001.03.002 |