 2020,
Vol. 29
2020,
Vol. 29

2. 上海海洋大学 上海水产养殖工程技术研究中心, 上海 201306;
3. 上海海洋大学 农业农村部鱼类营养与环境生态研究中心, 上海 201306;
4. 江苏省滆湖渔业管理委员会办公室, 江苏 常州 213161
富营养化导致水生态系统藻类大量增殖,淡水生态系统发生蓝藻水华、海洋生态系统发生赤潮。国内外常采用经典生物操纵和非经典生物操纵技术控制藻类生物量的异常增殖。对淡水生态系统而言,以鲢鳙作为生态修复工具种的非经典生物操纵技术既有大量成功案例,也有一些不能成功控制藻类生物量的报道[1]。有学者[1]认为,鲢鳙非经典生物操纵容易导致藻类小型化,可能导致藻类生物量不降反增,这也是非经典生物操纵引起争议的一个方面。因此,人们常结合鱼蚌(贝)开展小尺度控藻和鱼贝混养试验及野外大尺度生态修复实验[2-4],以便同时对大型和小型藻类产生摄食压力,贝类也单独用于湖库水质的调控[5]。
摄食率是反映滤食性贝类生理活动和生态功能的重要指标[6-7],贝类摄食率的测定可以为定量评估其在生态系统中的作用提供必要参数[8]。国内外对贝类摄食率开展了较多的研究,但在国内主要的研究对象集中在海洋经济贝类[9-11],对淡水贝类的研究较为少见[12-13]。国内学者[8, 12-14]主要开展了淡水贝类摄食率种间比较、对不同藻类的选择性及温度、饵料(藻类)密度、规格、pH等对其摄食率影响等方面的研究,并就其对浮游植物及水质影响进行了报道[8, 15-19]。对海洋贝类摄食率的主要研究内容亦如此,但因为环境的差异,学者就盐度对海洋贝类摄食率影响的研究较多[20]。尽管存在贝类摄食率影响因素的研究,但对它们的交互作用还少有报道。河蚬(Corbicula fluminea)和背角无齿蚌(Anodonta woodiana)为我国重要的淡水经济贝类,是湖泊等淡水生态系统中重要的滤食性底栖生物。本文研究这两种贝类对3种藻类的摄食率及贝类规格和放养密度对摄食率的影响,并探讨这些因素对它们摄食率的交互作用,从而为利用贝类控制湖库藻类提供基础数据和理论依据。
1 材料与方法 1.1 实验材料背角无齿蚌来自江苏滆湖,河蚬取自上海市青浦区淀山湖。实验贝取回后,挑选活力强,无损伤个体暂养于水箱内,水箱内为提前曝气24 h的自来水,水温维持在(20±1)℃。贝类暂养期间,每天换水1次,根据摄食实验安排投喂相应藻类以驯化适应投喂环境,并连续充气。实验前1天停止投喂。
随机选择健康的背角无齿蚌及河蚬。选用的藻类为铜绿微囊藻(Microcystis aeruginosa)、小球藻(Chlorella pyrenoidosa)和斜生栅藻(Scenedesmus obliquus),均购于中国科学院水生生物研究所,用BG-11培养基培养于5 L锥形瓶中,锥形瓶放入恒温光照培养箱,培养箱温度为(26±0.5) ℃,光照条件为12L:12D。
1.2 实验设计及方法背角无齿蚌摄食实验在水箱(长380 mm、宽275 mm、高140 mm)中进行,实验前倒入6 000 mL的曝气自来水,水深57 mm。密度设置3个水平,即每个水箱分别放入2、4和6个背角无齿蚌,每个密度设置2种贝类规格(表 1)。河蚬摄食实验在1 L的烧杯(直径105 mm、高150 mm)中进行,实验前倒入750 mL曝气自来水,水深87 mm。河蚬密度设置4个水平,即5、20、35、50 g(湿质量)/750 mL,每个密度设置2种河蚬规格(表 1)。河蚬和背角无齿蚌的实验水温均控制在(20±1)℃,分别投喂铜绿微囊藻、小球藻、斜生栅藻,藻类投放密度均为5×105 cells/mL。因此对每种贝类而言,每个处理投喂1种藻类、贝类放置1种密度和1种规格,每个处理设置3个重复,同时将装有同样体积的无贝藻液作为对照。实验结束,采集1.5 mL水样于2 mL离心管中,加鲁戈氏液固定待镜检,采用血球计数板于40×显微镜下计数。水样采集前,采用溶氧仪(YSI-550A)测定实验容器水体中溶解氧含量。实验结束后,去除双壳后将软体组织放入烘箱60 ℃烘干至恒重并称量,湿质量精确到0.01 g、软组织干质量精确到0.001 g,贝类壳长、壳宽、壳高测定精确到0.01 mm,两种贝类规格的形态学参数见表 1。

|
表 1 背角无齿蚌和河蚬两种规格的形态学参数 Tab.1 The morphological parameters of two shellfish species |
贝类摄食率(Feeding rate)[10]计算公式:
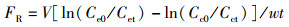 (1)
(1)
式中:FR为摄食率;V为实验溶液体积,mL;w为实验用贝的组织干质量,g;t为实验时间,h;Ce0为实验开始时的藻类细胞含量,cells/mL;Cet为t时间的藻类细胞含量,cells/mL;Cc0为实验开始时空白对照组的藻类细胞含量,cells/mL;Cct为t时间空白对照组的藻类细胞含量,cells/mL。
1.4 数据处理将藻种作为协变量,采用单因素协方差分析比较河蚬和背角无齿蚌摄食率的差异性。采用三因素(藻种、密度和规格)方差分析,探讨藻种、密度和规格对两种贝类摄食率的影响,多重比较采用Duncan氏检验。采用SPSS 19.0软件进行分析,P < 0.05表示存在显著差异,P < 0.01表示存在极显著差异。
2 结果 2.1 两种贝类摄食率比较单因素协方差分析表明(藻类作为协变量),投喂藻类对两者的摄食率无影响(F=1.416,P=0.236),河蚬的摄食率(92.7±2.3) mL/(g·h)显著高于背角无齿蚌(40.8±1.9) mL/(g·h)(F=274.825,P < 0.001)。
2.2 三因素对背角无齿蚌和河蚬摄食率的影响 2.2.1 三因素对背角无齿蚌摄食率的影响三因素方差分析表明,背角无齿蚌对藻类摄食率无显著种间差异(表 2),背角无齿蚌对铜绿微囊藻、小球藻和斜生栅藻的摄食率分别为(42.0±3.6)、(40.4±2.8)和(39.9±3.5)mL/(g·h)。但贝规格和贝密度显著影响背角无齿蚌的摄食率,且3个因素对背角无齿蚌的摄食率无交互作用(表 2)。小规格蚌的摄食率(50.8±1.9)mL/(g·h)显著大于大规格(30.7±1.7)mL/(g·h)。低密度蚌的摄食率显著高于高密度,而中密度与低密度和高密度间均无显著差异。见图 1。

|
表 2 藻种、贝规格和密度对背角无齿蚌摄食率影响的三因素方差分析 Tab.2 Three-way ANOVA of algae species, shellfish size and density on filter-feeding rate of Anodonta woodiana |

|
不同小写字母说明两者间差异显著(P<0.05);LD代表低密度;MD代表中密度;HD代表高密度 Different lowercase letters in the table indicate a significant difference between the two treatment groups (P<0.05); LD.low density of shellfish, MD.middle density of shellfish, HD.high density of shellfish 图 1 蚌密度对背角无齿蚌摄食率的影响 Fig. 1 Effect of shellfish density on the feeding rate of Anodonta woodiana |
三因素方差分析表明,藻种、贝规格和贝密度对河蚬的摄食率均有显著影响,贝规格和密度对摄食率有交互作用(表 3)。Duncan氏多重比较显示,河蚬对斜生栅藻[97.3±3.5 mL/(g·h)]和小球藻[93.0±4.4 mL/(g·h)]的摄食率均显著高于其对铜绿微囊藻的摄食率[87.8±4.0 mL/(g·h)]。

|
表 3 藻种、贝规格和密度对河蚬摄食率影响的3因素方差分析 Tab.3 Three-way ANOVA of algae species, shellfish size and density on filter-feeding rate of Corbicula fluminea |
单因素方差分析表明,贝密度对小规格河蚬摄食率有显著影响(F=61.680,P < 0.001),摄食率各密度间均存在显著差异(Duncan氏多重比较,图 2)。贝密度对大规格河蚬的摄食率也有显著影响(F=19.784,P < 0.001),密度1显著高于密度2和密度3,后两者又显著高于密度4,密度2和密度3之间摄食率无显著差异(Duncan氏多重比较,图 2)。t-检验表明,在密度1(t=3.050,P=0.008)和密度2(t=2.798,P=0.013)情况下,均是小规格河蚬摄食率显著大于大规格(图 2)。在密度3(t=2.034,P=0.059)和密度4(t=0.966,P=0.348)情况下,规格对摄食率均无显著影响。

|
图中不同大写字母表示同一密度条件下贝类的摄食率在规格间存在显著差异(P < 0.05),不同小写字母表示同一规格贝类摄食率在密度间存在显著差异(P < 0.05);D1代表最低密度;D2代表第二低密度;D3代表第二高密度;D4代表最高密度 Different uppercase letters in the figure indicate significant differences in feeding rate between shellfish size at each density (P < 0.05), and different lowercase letters indicate significant differences in feeding rates between shellfish densities at each shellfish size (P < 0.05); D1:the lowest density; D2:the second low density; D3:the second high density; D4:the highest density 图 2 贝密度和规格对河蚬摄食率的影响 Fig. 2 Effect of shellfish density and size on the feeding rate of Corbicula fluminea |
藻种对水体溶解氧均无显著影响(所有F < 2.901,P>0.067),且贝类密度和规格均显著影响水体溶解氧(所有F>9.992,P < 0.01)。见表 4~6。藻种和背角无齿蚌规格(F=12.812,P < 0.001)及河蚬规格和密度对溶解氧产生交互作用(F=7.167,P < 0.001),其他因素两两间无交互作用(P>0.05)。

|
表 4 两种贝处于不同藻类水环境中的溶解氧含量
Tab.4 Dissolved oxygen content of different algal water environment of two shellfish species |

|
表 5 不同规格两种贝所处水环境的溶解氧含量
Tab.5 Dissolved oxygen content of the water environment of different body sizes of two shellfish species |

|
表 6 放置不同密度背角无齿蚌和河蚬容器中的水体溶解氧含量
Tab.6 Dissolved oxygen content of the waters in the container stocked with different densities of Anodonta woodiana and Corbicula fluminea |
背角无齿蚌(r=0.426,P=0.001,图 3a)和河蚬(r=0.715,P < 0.001,图 3b)摄食率与溶解氧含量显著正相关。

|
图 3 背角无齿蚌(a)和河蚬(b)摄食率与溶解氧的关系 Fig. 3 Relationship between feeding rate and dissolved oxygen for Anodonta woodiana (a) and Corbicula fluminea (b) |
彭建华等[21]报道的背角无齿蚌摄食率约为本文的1.47倍;张许峰等[13]报道背角无齿蚌的摄食率是本文的3.48倍,报道河蚬的摄食率是本文的2.91倍。张许峰等[13]研究表明:不同贝类物种的摄食率差异很大,甚至同一物种不同学者的研究结果也存在很大变异。许多因素会造成不同学者对同一贝类物种摄食率的差异,如温度、盐度、受试贝类的规格和密度(或生物量)、投喂藻类的情况(密度及种类)以及贝类试验水体的尺度等,如喂以混合藻类的褶纹冠蚌、背角无齿蚌和河蚬的摄食率均分别大于喂以单一藻类的[13],贝类最高藻类摄食率出现在某一温度或盐度条件[11, 13, 22]。同时,实验中饵料充足与否也会影响摄食率,为了真实反映某一饵料含量下的贝类摄食率,建议将实验结束时的饵料含量控制在实验开始时含量的30%~40%[11]。另外,不同文献报道的贝类摄食率单位的不同也降低了物种内和物种间差异的可比较性,如mg(或mL)/(g·h)、mg(或mL)/(ind·h)等,单位中的体质量数据有的文献使用的是组织干质量、有的文献使用的是湿质量。因此,建议学者在开展此类研究时,能够统一单位,笔者认为使用mg/(g·h)比较好,其中体质量建议使用软体组织干质量。同时,由于摄食率受到众多因素如体质量的影响,建议在方法中尽量交代清楚实验条件,如实验动物的壳长和体质量数据。
贝类对藻类是否有选择性以及对不同单胞藻摄食率是否存在显著差异,不同学者研究结果不尽一致。BAKER等[23]实验提出,3种淡水北美蚌类均对微囊藻有偏好性;费志良等[24]认为,三角帆蚌对微囊藻的摄食选择性很高;谢磊等[25]认为,一些藻类可能产生某种生物活性物质引发贝类的生理应激,从而对贝类滤食产生影响,但这种所谓的活性物质并未成功分离,研究有待继续。与此不同的是,在单一藻类投喂条件下,褶纹冠蚌、河蚬和背角无齿蚌3种藻类的摄食率无显著差异[13]。本实验背角无齿蚌对3种藻类的摄食率无显著差异,与张许峰等[13]报道的结果一致,可能与它们对藻类无选择性有关[13-14],如三角帆蚌对滇池试验区和附近池塘水体中藻类无显著偏好[14]。河蚬对铜绿微囊藻的摄食率显著低于对其他两种绿藻,与张许峰等[13]报道的结果有差异,可能与微囊藻对不同体型大小的贝类摄食率影响存在差异有关,微囊藻毒素没有抑制圆顶珠蚌的摄食,但对河蚬的摄食有抑制作用[26]。
身体大小(规格)具有重大的生态学意义,通常小个体比大个体具有更高的生理速率(如新陈代谢速率)。不同贝类物种的摄食率(单位体质量单位时间摄食率)也如此,如褶纹冠蚌的最大、背角无齿蚌次之、河蚬最小[13],较大规格三角帆蚌的摄食率小于较小规格背角无齿蚌[21]。同种物种不同规格之间亦如此,如背角无齿蚌[8, 14]、三角帆蚌[21]、尖紫蛤[10]、栉孔扇贝[9]。本文研究结果与上述学者的研究结果类似,种间比较而言,体型较大的背角无齿蚌的摄食率显著小于体型较小的河蚬,大规格背角无齿蚌的摄食率显著小于小规格,在2种较低密度下,大规格河蚬的摄食率亦显著低于小规格。
贝类对藻类的摄食率及去除效果与密度有关,有些研究表明,藻类的去除量随着贝类密度的增加而显著增加[17]。另外一些研究则显示,贝类对藻类的去除效果受密度制约[17],如太湖围隔中的蚌类(主要指褶纹冠蚌)在中等生物量时对藻类、叶绿素a和悬浮物的去除效果最好,因为生物量过大导致了单位消除量的下降[15],这可能与贝类的单位体质量摄食率随着密度或生物量的增加而下降有关[9]。本文结果与此一致,背角无齿蚌和河蚬的摄食率随着贝类密度的增加而显著下降,且密度和规格对背角无齿蚌摄食率无交互作用,而它们对河蚬具有交互作用。推测可能与溶解氧有关,尽管不同密度背角无齿蚌水体溶解氧差异显著,但高密度下,溶解氧仍高于5 mg/L,而2种高密度河蚬实验环境条件下,溶解氧低于2.3 mg/L。摄食率与溶解氧的相关性河蚬明显高于背角无齿蚌,也能说明河蚬实验水体溶解氧的急剧下降导致了密度和规格对两种贝类摄食率交互作用的差异。在2种较低密度下,小规格河蚬个体的摄食率分别显著高于大规格个体;而在2种较高密度环境下,两者均无显著差异,这可能与小规格河蚬个体对胁迫因子如低氧的耐受能力低于大规格个体有关,如小规格尖紫蛤对海水盐度和pH的敏感性比大规格个体更强[10]。
| [1] |
ZHANG X, XIE P, HUANG X P. A review of nontraditional biomanipulation[J]. The Scientific World Journal, 2008, 8: 1184-1196. DOI:10.1100/tsw.2008.144 |
| [2] |
孟顺龙, 陈家长, 胡庚东, 等. 滤食性动物放流对西五里湖的生态修复作用初探[J]. 中国农学通报, 2009, 25(16): 225-230. MENG S L, CHEN J Z, HU G D, et al. Preliminary study on eco-restoration effect of releasing suspension-feeding animal on West Wuli Lake[J]. Chinese Agricultural Science Bulletin, 2009, 25(16): 225-230. |
| [3] |
周小玉, 张根芳, 刘其根, 等. 鲢、鳙对三角帆蚌池塘藻类影响的围隔实验[J]. 水产学报, 2011, 35(5): 729-737. ZHOU X Y, ZHANG G F, LIU Q G, et al. Effects of Hyriopsis cumingii and Aristichthys nobilis on the enclosures phytoplankton community of Hypophthalmichthys molitrix pond[J]. Journal of Fisheries of China, 2011, 35(5): 729-737. |
| [4] |
唐金玉, 王岩, 戴杨鑫. 在淡水鱼类混养系统中吊养三角帆蚌对养殖产量和水质的影响[J]. 水产学报, 2014, 38(2): 208-217. TANG J Y, WANG Y, DAI Y X. Effects of co-culturing freshwater pearl mussel Hyriopsis cumingii in a fish polycultural system on fish yield and water quality[J]. Journal of Fisheries of China, 2014, 38(2): 208-217. |
| [5] |
彭建华, 陈文祥, 刘家寿, 等. 三角帆蚌对水库水污染的水质调控模型及作用[J]. 环境科学与技术, 2009, 32(2): 46-48. PENG J H, CHEN W X, LIU J S, et al. Water quality model of Hyriopsis cumingii and effect in reservoir pollution control[J]. Environmental Science & Technology, 2009, 32(2): 46-48. DOI:10.3969/j.issn.1003-6504.2009.02.013 |
| [6] |
边学森, 刘洪波, 甘居利, 等. 太湖背角无齿蚌中多氯联苯的残留[J]. 农业环境科学学报, 2008, 27(2): 767-772. BIAN X S, LIU H B, GAN J L, et al. Residues of PCBs in Anodonta woodiana from the Taihu Lake, China[J]. Journal of Agro-Environment Science, 2008, 27(2): 767-772. DOI:10.3321/j.issn:1672-2043.2008.02.067 |
| [7] |
NADDAFI R, PETTERSSON K, EKLÖV P. The effect of seasonal variation in selective feeding by zebra mussels(Dreissena polymorpha)on phytoplankton community composition[J]. Freshwater Biology, 2007, 52(5): 823-842. DOI:10.1111/j.1365-2427.2007.01732.x |
| [8] |
丁涛, 李林, 彭亮, 等. 背角无齿蚌摄食率及对水中叶绿素α清除能力的研究[J]. 水生生物学报, 2010, 34(4): 779-786. DING T, LI L, PENG L, et al. Studies on elimination of chlorophyll-a and ingestion rate in eutrophic water by Anodonta woodiana elliptica[J]. Acta Hydrobiologica Sinica, 2010, 34(4): 779-786. |
| [9] |
匡世焕, 方建光, 孙惠玲, 等. 桑沟湾栉孔扇贝不同季节滤水率和同化率的比较[J]. 海洋与湖沼, 1996, 27(2): 194-199. KUANG S H, FANG J G, SUN H L, et al. Seasonal variation of filtration rate and assimilation efficiency of scallop Chlamys farreri in Sanggou Bay[J]. Oceanologia et Limnologia Sinica, 1996, 27(2): 194-199. DOI:10.3321/j.issn:0029-814X.1996.02.013 |
| [10] |
黄洋, 黄海立, 邓乐平, 等. 盐度、pH和规格对尖紫蛤滤水率、摄食率、吸收率的影响[J]. 广东海洋大学学报, 2014, 34(1): 42-47. HUANG Y, HUANG H L, DENG L P, et al. Effects of salinity, pH and sizes on clearance rate, ingestion rate and absorption efficiency of Soletellina acuta[J]. Journal of Guangdong Ocean University, 2014, 34(1): 42-47. |
| [11] |
吕昊泽, 刘健, 陈锦辉, 等. 盐度对长江口3种滤食性贝类滤水率、摄食率、同化率的影响[J]. 海洋科学, 2016, 40(8): 10-17. LYU H Z, LIU J, CHEN J H, et al. Effects of salinity on filtration, ingestion, and assimilation rates of three filter-feeding bivalves in the Yangtze River estuary[J]. Marine Sciences, 2016, 40(8): 10-17. |
| [12] |
朱爱民, 陈文祥, 栾建国, 等. 三角帆蚌食性及摄食率的初步研究[J]. 水生生物学报, 2006, 30(2): 244-246. ZHU A M, CHEN W X, LUAN J G, et al. A study on feeding habits and ingestion rate of Hyriopsis cumingii[J]. Acta Hydrobiologica Sinica, 2006, 30(2): 244-246. DOI:10.3321/j.issn:1000-3207.2006.02.021 |
| [13] |
张许峰, 吴小平, 欧阳珊, 等. 四种淡水贝的摄食率和耗氧率的实验研究[J]. 海洋湖沼学报, 2008(3): 144-154. ZHANG X F, WU X P, OU Y S, et al. Experiment study on feeding rates and oxygen consumption rates of four species of freshwater mollusks[J]. Transactions of Oceanology and Limnology, 2008(3): 144-154. |
| [14] |
吴庆龙, 陈宇炜, 刘正文. 背角无齿蚌对浮游藻类的滤食选择性与滤水率研究[J]. 应用生态学报, 2005, 16(12): 2423-2427. WU Q L, CHEN Y W, LIU Z W. Filtering capacity of Anodonta woodiana and its feeding selectivity on phytoplankton[J]. Chinese Journal of Applied Ecology, 2005, 16(12): 2423-2427. DOI:10.3321/j.issn:1001-9332.2005.12.040 |
| [15] |
潘建林, 徐在宽, 唐建清, 等. 湖泊大型贝类控藻与净化水质的研究[J]. 海洋湖沼通报, 2007(2): 69-79. PAN J L, XU Z K, TANG J Q, et al. Study on the effects of large mollusks on alge control and water quality at meiliang gulf in Taihou lake[J]. Transactions of Oceanology and Limnology, 2007(2): 69-79. DOI:10.3969/j.issn.1003-6482.2007.02.012 |
| [16] |
杨东妹, 陈宇炜, 刘正文, 等. 背角无齿蚌滤食对营养盐和浮游藻类结构影响的模拟[J]. 湖泊科学, 2008, 20(2): 228-234. YANG D M, CHEN Y W, LIU Z W, et al. Top-down effects of Anodonta woodiana on nutrient concentration & phytoplankton community composition in a microcosm ecosystem[J]. Journal of Lake Sciences, 2008, 20(2): 228-234. DOI:10.3321/j.issn:1003-5427.2008.02.015 |
| [17] |
徐海军, 凌去非, 杨彩根, 等. 3种淡水贝类对藻类消除作用的初步研究[J]. 水生态学杂志, 2010, 3(1): 72-75. XU H J, LING Q F, YANG C G, et al. Preliminary studies on the elimination effect of algae by three species of freshwater bivalve[J]. Journal of Hydroecology, 2010, 3(1): 72-75. |
| [18] |
林军, 邓明星, 章守宇, 等. 贻贝养殖海域表层水温季节变化及其对紫贻贝生长的影响[J]. 上海海洋大学学报, 2016, 25(6): 918-926. LIN J, DENG M X, ZHANG S Y, et al. Seasonal variation of surface water temperature and its ecological impacts in a mussel aquaculture farm[J]. Journal of Shanghai Ocean University, 2016, 25(6): 918-926. |
| [19] |
王旭, 赵旭, 章守宇, 等. 枸杞岛贻贝养殖水域碳氮磷分布格局[J]. 水产学报, 2015, 39(11): 1650-1664. WANG X, ZHAO X, ZHANG S Y, et al. Distribution pattern of dissolved carbon, nitrogen and phosphorus in mussel culture areas of Gouqi Island[J]. Journal of Fisheries of China, 2015, 39(11): 1650-1664. |
| [20] |
黄洋, 黄海立, 杨土新, 等. 体质量和温度对尖紫蛤滤水率、摄食率、吸收率的影响[J]. 浙江海洋学院学报(自然科学版), 2013, 32(2): 109-114. HUANG Y, HUANG H L, YANG S X, et al. Effects of body mass and temperature on clearance rate, ingestion rate absorption efficiency of Soletellina acuta[J]. Journal of Zhejiang Ocean University (Natural Science), 2013, 32(2): 109-114. DOI:10.3969/j.issn.1008-830X.2013.02.004 |
| [21] |
邱哲文, 刘婧, 赵开彬, 等. 长江口北支滩涂贝类放流对潮下带大型底栖动物群落的影响[J]. 上海海洋大学学报, 2019, 28(2): 277-282. QIU Z W, LIU J, ZHAO K B, et al. Effects of shellfish release on macrobenthic communities in the subtidal zone in the northern branch of the Yangtze River Estuary[J]. Journal of Shanghai Ocean University, 2019, 28(2): 277-282. |
| [22] |
徐钢春, 顾若波, 闻海波, 等. 温度、体重和饵料密度对河蚬滤食率与同化率的影响[J]. 上海水产大学学报, 2007, 16(2): 151-156. XU G C, GU R B, WEN H B, et al. Effect of tempreature, body weight, food density on filter feeding rate and assimilation efficiency of Corbicula fluminea[J]. Journal of Shanghai Fisheries University, 2007, 16(2): 151-156. |
| [23] |
BAKER S M, LEVINTON J S. Selective feeding by three native North American freshwater mussels implies food competition with zebra mussels[J]. Hydrobiologia, 2003, 505(1/3): 97-105. |
| [24] |
费志良, 廖馨, 赵沐子, 等. 三角帆蚌(Hyriopsis cumingii Lea)对富营养化水体悬浮物消除的时效模型[J]. 南京大学学报(自然科学版), 2005, 41(6): 634-638. FEI Z L, LIAO X, ZHAO M Z, et al. Time-effect model for Hyriopsis cumingii Lea's elimination of suspended matter in eutrophic water[J]. Journal of Nanjing University (Natural Sciences), 2005, 41(6): 634-638. DOI:10.3321/j.issn:0469-5097.2005.06.007 |
| [25] |
谢磊, 徐晓娇, 江天久. 抑食金球藻对翡翠贻贝抗氧化酶系统的影响[J]. 水产学报, 2018, 42(6): 846-853. XIE L, XU X J, JIANG T J. Effect of Aureococcus anophagefferens on the antioxidant enzyme system in Pernaviridis[J]. Journal of Fisheries of China, 2018, 42(6): 846-853. |
| [26] |
刘莹.铜绿微囊藻对枝角类及双壳类毒性影响的实验研究[D].南昌: 南昌大学, 2006: 8. LIU Y. Experimental studies on the toxic effects of Microcystis aeruginosa on cladocerans and bivalves[D]. Nanchang: Nanchang University, 2006: 8. http://cdmd.cnki.com.cn/Article/CDMD-11902-2006121803.htm |
 ,
ZHANG Mingxing1,2,3,
CHEN Liping1,2,3,
JIA Xixi1,2,3,
KONG Youjia4,
HU Zhongjun1,2,3
,
ZHANG Mingxing1,2,3,
CHEN Liping1,2,3,
JIA Xixi1,2,3,
KONG Youjia4,
HU Zhongjun1,2,3
2. Shanghai Engineering Research Center of Aquaculture, Shanghai Ocean University, Shanghai 201306, China;
3. Centre for Research on Environmental Ecology and Fish Nutrion, Ministry of Agriculture and Rural Affairs, Shanghai Ocean University, Shanghai 201306, China;
4. Fisheries Management Commission of Lake Gehu, Changzhou 213161, Jiangsu, China