 2017,
Vol. 26
2017,
Vol. 26



2. 远洋渔业协同创新中心, 上海 201306;
3. 国家远洋渔业工程技术研究中心, 上海 201306;
4. 大洋渔业资源可持续开发省部共建教育部重点实验室, 上海 201306
淀山湖的虾类资源以日本沼虾和秀丽白虾为主[1],日本沼虾(Macrobrachium nipponense)隶属于长臂虾科(Palaemonidae)、沼虾属(Macrobrachium),广泛分布于我国各地的江河、湖泊、水库、池塘及沟渠中,是我国淡水虾类中一个重要种类[2],也是可供出口的水产品[3],同时是多种名优鱼类的天然饵料[4]。秀丽白虾(Exopalaemon modestus)隶属于长臂虾科(Palaemonidae)、白虾属(Exopalaemon),主要分布于大型湖泊,如太湖、巢湖、白马湖、呼伦湖等[5],在水域生态系统中,秀丽白虾既是捕食者又是消费者,在食物链中发挥着重要作用[6]。所以对淀山湖日本沼虾和秀丽白虾的资源状况和时空变化进行研究具有重要意义。有关虾类的研究主要涉及到繁殖、生长和摄食等生物学特性方面[7-11],而有关资源动态的研究尚少,在国内仅温周瑞等[12]曾对太湖水域的虾类资源量动态进行过研究。目前,有关淀山湖的虾类研究,仅见刘仕鑫等[13]对秀丽白虾的通径分析。因此,本研究以2010年8月—2011年8月对日本沼虾和秀丽白虾的资源调查为基础,分析其资源量的周年动态变化,为合理管理和有效捕捞淀山湖虾类资源提供依据,也为渔业主管部门划定增殖放流区域提供参考。
1 材料与方法 1.1 调查时间和站点信息调查区域为淀山湖水域(31°06'N,120°58'E),地处江苏、浙江、上海两省一市交界地带,分属江苏昆山市和上海青浦区管辖,为苏州河和黄浦江的源头之一,湖泊面积为63.7 km2,其中上海境内47 km2,最大水深6.36 m,平均水深2.5 m,是上海最大的天然湖泊。在商榻与淀峰村河道之间画一条线,以此作为分界线,将淀山湖分成两段湖区,即上半段湖区(S1,S2,S3,S4,S5,S6)和下半段湖区(S7,S8,S9,S10)。根据前期的初步调查及当地渔民的访问,设置10个采样点,调查站点见图 1。调查时间为2010年8月— 2011年8月,每月一次虾拖网调查,调查时间涵盖了淀山湖的四个季节。虾拖网的网目为6 cm,网长3 m,网高1.8 m,网口宽2 m,平均拖速为3.3 km/h,平均拖网时间为30 min。调查人员将采集到的日本沼虾和秀丽白虾用保温箱带至实验室,采用游标卡尺测定头胸甲长,精确到0.01 mm,并记录每次作业的总渔获尾数和重量,重量精确到0.1 g。本次调查共采集到日本沼虾20 055尾,秀丽白虾17 538尾。

|
图 1 淀山湖虾拖网调查站点分布
Fig. 1 Sampling stations of trawl net in Dianshan Lake
|
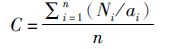 (1)
(1)
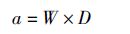 (2)
(2)
式中:Ni为第i网捕获数量和重量;ai为第i网拖过的面积;n为捕捞次数;C为单位面积捕获数量或重量;W表示拖网的有效宽度;D表示拖网的拖行距离。本文假设拖网的有效宽度为拖网宽度,每个站点的拖网时间和拖速恒定不变,取平均值。
1.3 现存量估算根据单位捕捞数量或重量,种群占据面积来估算种群的现存量[15-16]
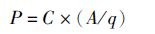 (3)
(3)
式中:P为现存量;A为种群占据面积;q为捕捞系数,即q=1-逃逸率;根据本次调查拖网网具特性,假设逃逸率为零[17],即q=1。
1.4 季节变化和空间变化分析方法 1.4.1 月变化分析对日本沼虾和秀丽白虾的数量、重量及头胸甲长进行统计分析,分别绘制出日本沼虾和秀丽白虾所有站点平均数量密度、平均重量密度、平均头胸甲长及头胸甲长组频率的月变化趋势图;以及不同站点日本沼虾和秀丽白虾的数量密度和重量密度的月变化趋势图。秀丽白虾5月份头胸甲长数据缺失,故没对这个月进行分析。
1.4.2 空间变化分析利用全年的平均数量密度和平均质量密度进行空间变化分析;通过分析发现资源密度按月变动情况不是十分明显,所以将时间尺度拉大,通过大尺度的分析来查看结果,故将2010年8月—2011年8月分为四个季节,即春季(3—5月)、夏季(6—8月)、秋季(9—11月)、冬季(12—2月),按季度分别对日本沼虾和秀丽白虾数量密度和重量密度进行空间变化分析。
1.4.3 方差显著性分析运用无重复双因素方差分析虾类资源密度在不同季节和不同站点中是否存在显著性差异,单因素方差分析两种虾类资源密度和头胸甲长月变化趋势是否存在显著性差异。为了使数据具有更好的方差齐性,所有用于统计分析的数据均进行平方根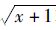
采用Excel 2013、SPSS 22.0和ArcGIS 10.2软件进行数据分析。
2 结果 2.1 淀山湖日本沼虾和秀丽白虾头胸甲长月变化根据采集的数据,通过单因素方差分析,发现日本沼虾和秀丽白虾头胸甲长的月变化趋势相似,两种虾头胸甲长月变化无显著性差异(P>0.05)。日本沼虾头胸甲长范围为7. 05~47. 07 mm,平均值为(22. 192 ± 1. 650)mm;秀丽白虾头胸甲长范围为9. 10~38. 7 mm,平均值为(21. 221 ± 1. 598)mm,日本沼虾5月达到最大值,秀丽白虾6月达到最大值,分别为25. 387 mm和23. 009 mm,之后呈递减趋势,至10月这两种虾资源密度都趋于平稳,这两种虾的头胸甲长均在冬季期间(12—2月)达到最小值(图 2)。

|
图 2 淀山湖日本沼虾和秀丽白虾头胸甲长月变化
Fig. 2 Monthly variation of the carapace length for M. nipponense and E. modestus in Dianshan Lake
|
淀山湖日本沼虾优势头胸甲长组为15.5~30.5 mm,占82.98 %。5—10月头胸甲长组(20.5~30.5 mm)的频率要比其余月份高,而其余月份头胸甲长组(15.5~20.5 mm)的频率要比5—10月份高;5—8月和12月采集到40.5~50.5 mm头胸甲长组的日本沼虾,其他月份均未采集到。11—3月期间,头胸甲长组20.5~25.5 mm的频率逐渐降低,而头胸甲长组15.5~20.5 mm频率逐渐增加,且头胸甲长组5.5~10.5 mm开始出现,另外8月也有头胸甲长组5.5~10.5 mm的小个体的出现(图 3)。

|
图 3 淀山湖日本沼虾头胸甲长组成频率月变化
Fig. 3 Monthly variation of frequency of carapace length for M. nipponense in Dianshan Lake
|
淀山湖秀丽白虾优势头胸甲长组15. 5 ~ 25. 5 mm,占80. 49 %。 6月份和9—12月头胸甲长组(20. 5~25. 5 mm)的频率要比其余月份高;而其余月份头胸甲长组(15. 5 ~ 20.5 mm)的频率要比6月和9—12月高;35. 5 ~ 40. 5 mm头胸甲长组的秀丽白虾仅仅在1月、4月、10月和11月出现,其他月份均未出现;5. 5 ~10. 5 mm头胸甲长组的秀丽白虾仅仅在1月、4月、12月出现,其他月份均未出现(图 4)。

|
图 4 淀山湖秀丽白虾头胸甲长组成频率月变化
Fig. 4 Monthly variation of frequency of the carapace length for E. modestus in Dianshan Lake
|
日本沼虾数量密度和质量密度的月变化趋势相似(图 5),无显著性差异(P>0. 05),9月份达到最大值,分别为0. 037尾/m2和0. 029 g/m2,随后迅速下降至11月,11—1月有回升趋势,1—3月开始下降取得最小值,分别为0.004尾/m2和0. 006 g/m2,之后又略微上升,至5月达到第二次高峰,其值分别为0. 011尾/m2和0.022 g/m2,随后再次下降。

|
图 5 淀山湖日本沼虾数量密度和质量密度月变化
Fig. 5 Monthly variation of the total quantity density and weight density of M. nipponense in Dianshan Lake
|
秀丽白虾数量密度和质量密度的月变化趋势相似,无显著性差异(P> 0. 05),8月秀丽白虾的数量密度和质量密度均达到最大值,分别为0.034 尾/m2和0. 032 g/m2,随后直线下降至12月,12—2月略微回升,2—3月下降,之后上升至4月第二次取得峰值,分别为0. 005尾/m2和 0. 010 g/m2,4—6月下降至最小值,分别为0. 001尾/m2和0. 001 g/m2,随后再次上升至8月(图 6)。

|
图 6 淀山湖秀丽白虾数量密度和质量密度月变化
Fig. 6 Monthly variation of the total quantity density and weight density of E.modestus in Dianshan Lake
|
日本沼虾数量密度和质量密度各站点月变化趋势相似,无显著性差异(P>0. 05),各站点日本沼虾的数量密度和重量密度在9月均出现峰值,5月第二次取得峰值,其他站点各月份数量密度和重量密度变化不大;9月3号站点数量密度达到最大值,为0. 042尾/m2;而重量密度也在9月3号站点达到最大值,为0. 058 g/m2(图 7)。

|
图 7 淀山湖日本沼虾各站点数量密度和质量密度的逐月变化
Fig. 7 Monthly variation of quantity density and weight density of M. nipponense in Dianshan Lake
|
秀丽白虾数量密度和质量密度各站点的月变化趋势相似,无显著性差异(P > 0. 05),各站点均在9月取得最大值,4月第二次取得峰值,其他月份变化趋势不明显(图 8)。

|
图 8 淀山湖秀丽白虾各站点数量密度和质量密度的逐月变化
Fig. 8 Monthly variation of quantity density and weight density of E.modestus in Dianshan Lake
|
日本沼虾全年数量密度和质量密度与站点呈极显著相关(P< 0.01),沿岸区域数量密度和质量密度普遍比敞水区高;北半段湖区的资源密度明显高于南半段湖区,且在北半段湖区东部沿岸区域资源密度较低,南半段湖区靠近淀峰村区域资源密度较低(表 1)。

|
表 1 淀山湖日本沼虾数量密度和质量密度的空间变化 Tab.1 Spatial variation of quantity density and weight density for M. nipponense in Dianshan Lake |
秀丽白虾数量密度和质量密度与站点呈极显著的相关性(P<0.01),与日本沼虾变化趋势相似(图 10),沿岸区域的数量密度和质量密度普遍高于敞水区;北半段湖区数量密度和质量密度略高于南半段湖区;同日本沼虾分布一样,北半段湖区东部沿岸区域密度较低。

|
表 2 淀山湖秀丽白虾数量密度和质量密度的空间变化 Tab.2 Spatial variation of quantity density and weight density for E. modestus in Dianshan Lake |
四个季节淀山湖日本沼虾和秀丽白虾数量密度南半段湖区小于北半段湖区(表 3和表 4);秋季和夏季沿岸区域资源密度高于敞水区,而冬季和春季有略微向敞水区移动的趋势。秀丽白虾北半段湖区与南段半湖区数量密度差别不大,仅略高于南半段湖区。另外,无论什么季节,上半段湖区东部的沿岸区域密度都较低,淀峰村附近区域也低于其余站点。

|
表 3 淀山湖日本沼虾各季度数量密度的空间变化 Tab.3 Temporal and spatial variation of quantity density of M. nipponensis in Dianshan Lake |

|
表 4 淀山湖秀丽白虾各季度数量密度的空间变化 Tab.4 Temporal and spatial variation of quantity density of E. modestus in Dianshan Lake |
这里用来估算日本沼虾和秀丽白虾资源量用的是当年渔获量的平均值,质量密度分别为0. 046 g/m2和0. 031 g/m2,数量密度分别为0. 032 尾/m2和0. 030尾/m2;该湖区的面积为63. 7 km2,根据公式(3)可以估算出淀山湖日本沼虾和秀丽白虾的资源量为2 930. 20 kg和197 4. 70 kg;估算尾数分别为2 038 400尾和1 911 000尾。由于各湖区日本沼虾和秀丽白虾分布不均和实际捕捞率不可能达到1,故估算出的资源量会偏低。
3 讨论 3.1 淀山湖秀丽白虾和日本沼虾月变化特征在冬季(12—2月)日本沼虾和秀丽白虾的质量密度、数量密度及头胸甲变化不大,但头胸甲长分别在5月和6月达到最大值,在11—3月和6—8月期间大个体(> 40. 5 mm)和小个体(< 10. 5 mm)同时出现,因而猜想这两种虾的繁殖期在这两个阶段,分别称其为秋季产卵群体和夏季产卵群体。温周瑞等[4, 14, 20]证明梅梁湾日本沼虾平均丰度和现存量冬季变化不大,与此同时,也证明太湖日本沼虾和秀丽白虾在11—2月种群生长基本处于停滞状态,3月开始生长,6月群体规格达到最大,7月因世代交替群体规格变小。而本研究头胸甲长在5月到达最大,可能是由于淀山湖与太湖的禁渔期不同所导致,还有可能是此时采集的样本大多属于产卵亲体。
虾类资源量与捕捞强度、环境因子、繁殖期、饵料资源、敌害生物等因子有关,冬季水温低,虾的活动能力弱,摄食不旺盛,生长缓慢,资源量相对稳定[18-19]。淀山湖拖网禁渔期是2月16日- 5月16日,随着禁渔期的结束,渔民便开始大规模和高强度的捕捞,使得淀山湖的资源不断下降,至冬季(11—2月),天气寒冷,风浪很大,渔民的作业受到很大的影响,所以淀山湖虾类受到的捕捞压力有所减缓,且夏季群体所产生的部分幼虾开始生长,从而导致冬季(11—2月)资源量密度有所提升。之后进入春季(3—5月),此时正是拖网禁渔期,水温回暖,虾类活动能力增强,摄食强度较冬季旺盛,刚好又是幼虾生长与发育的季节,因此资源量有所回升;随后进入夏季(5—7月)幼虾开始长大成熟,此时虾类头胸甲到达最大值,秋季产卵群体产卵之后便大量死亡,故资源量有所下降;到了秋季(8—10月)虾类幼体开始长大成熟,虽然个体不是最大的,但是随着虾类补充群体的加入,数量迅速增加至最大值,故此时资源量达到了最大值。
冬季产量低与饵料生物周年生长规律也有关,一般食物的出现率均是春夏秋季度出现率较高,冬季出现率较低[14],这也可能是导致冬季的产量降低的原因之一,另一原因可能是秋季产卵群体大量死亡所致。温周瑞等[21]也研究证明了冬季、春季、秋季日本沼虾的摄食高峰在黄昏前后,夏季的摄食高峰在上午和凌晨。冬季、春季和夏季秀丽白虾的摄食高峰基本在黄昏前后,秋季分别在14:00和凌晨。本次研究采样数据均来自早上的采样调查,采样可能与天气好坏有关,可能对本次研究结果产生影响,建议以后采样时间尽可能覆盖全天。
3.2 淀山湖秀丽白虾和日本沼虾空间变化特征不同湖区日本沼虾和秀丽白虾资源密度存在差异,反映其在空间分布上的特点。温周瑞和谢平[17]证明太湖日本沼虾和秀丽白虾空间变化特征与环境中食物丰富度有关,而李强等[22]对淀山湖浮游动物群落结构特征及其影响因子进行研究得出浮游生物多样性指数总体上上游高于下游的分布特征。康丽娟[23]证明淀山湖沉积物中营养物西北部高,东部低,进水口附近沉积物营养物浓度显著高于湖心及出水口。胡雪芹等[24]证明浮游植物叶绿素a浓度与浮游植物密度在水平分布上呈现北高南低的特点。吴阿娜等[25]证明淀山湖浮游植物叶绿素a浓度在进水口高(尤其是在S3站点),出水口低。这些都直接影响到水生生物的分布,然后间接影响到以这些水生生物为饵料的生物。另外,有研究表明日本沼虾与秀丽白虾的食性与食物碳源进行分析,得出这两种虾主要食物种类有有机碎屑、浮游植物、水生植物、原生动物、轮虫、枝角类、桡足类、底栖动物、鱼类等 [14, 26]。这些环境特点均可能是导致淀山湖北半段湖区资源量高于南半段湖区,而北半段湖区资源量主要集中在西北部,东北部偏低的空间分布特征的原因。南半段湖区、淀峰村和北半段湖区东部资源量比其他区域小,也可能是由于人类活动相对其他区域较频繁,人为因素所产生的污染物或者噪声导致这些区域的食物丰度降低,栖息环境受到破坏,致使这些区域的日本沼虾和秀丽白虾资源量较低。
严生良等[27]认为日本沼虾随季节变化有迁移现象。日本沼虾和秀丽白虾冬季、春季和夏季、秋季的空间变化与这两种虾的繁殖季节和生境类型密切有关[17, 28]。冬季和春季气温较低,摄食强度低下,活动性相对较弱,故潜伏至湖底水草丛中;夏季气温回升,虾类摄食强度较高,此时是虾类的繁殖季节,开始游向沿岸带进行繁殖,这便可能导致沿岸带资源明显高于敞水区,说明日本沼虾和秀丽白虾产卵亲体及当年繁殖的幼虾大量栖息于沿岸带。
| [1] | 俞存根, 宋海棠, 丁跃平, 等.浙江近海虾类资源量的初步评估[J]. 浙江水产学院学报, 1994, 13(3): 149–155. YU C G, SONG H T, DING Y P, et al.A primary estimation on shrimp resource of Zhejiang coastal area[J]. Journal of Zhejiang College of Fisheries, 1994, 13(3): 149–155. |
| [2] | 叶奕佐, 戈敏生, 王苹萍, 等. 淡水青虾养殖技术[M]. 武汉: 武汉出版社, 1997. YE Y Z, GE M S, WANG P P, et al. The farming technology of freshwater shrimp[M]. Wuhan: Wuhan Press, 1997. |
| [3] | 付辉云, 张燕萍, 方春林, 等.鄱阳湖日本沼虾捕捞种群生长与结构分析[J]. 江西水产科技, 2015: 17–20. FU H Y, ZHANG Y P, FANG C L, et al.Fishing population growth and structural analysis of freshwater shrimp: Macrobrachium nipponensis in Panyang Lake[J]. Jiangxi Fishery Science and Technology, 2015: 17–20. |
| [4] | 温周瑞, 谢平.太湖日本沼虾的生长特性[J]. 湖北农业科学, 2014, 53(18): 4374–4377. WEN Z R, XIE P.Growth characteristics of freshwater shrimp Macrobrachium nipponensis in Taihu Lake[J]. Hubei Agricultural Sciences, 2014, 53(18): 4374–4377. |
| [5] | 胡廷尖, 周志明, 黄鲜明, 等.秀丽白虾生物学特性及资源开发的初探[J]. 水利渔业, 2001, 21(2): 7–8. HU T J, ZHOU Z M, HUANG X M, et al.The primary research of biological characteristics and resource development of freshwater shrimp: Macrobrachium nipponensis[J]. Reservoir Fisheries, 2001, 21(2): 7–8. |
| [6] | XU J, ZHANG M, XIE P.Stable isotope changes in freshwater shrimps (Exopalaemon modestus and Macrobrachium nipponensis): trophic pattern implications[J]. Hydrobiology, 2008, 605(1): 45–54. DOI:10.1007/s10750-008-9299-y |
| [7] | OH C W, SUH H L, PARK K Y, et al.Growth and reproductive biology of the freshwater shrimp, Exopalaemon modestus (decapoda: palaemonidae) in a lake of Korea[J]. Journal of Crustacean Biology, 2002, 22(2): 357–366. DOI:10.1163/20021975-99990243 |
| [8] | 汪财生, 尹尚军, 钱国英, 等.日本沼虾染色体倍性检测技术的研究[J]. 海洋科学, 2006, 30(7): 5–8. WANG C S, YIN S J, QIAN G Y, et al.Detection of cellular DNA content in Macrobrachium nipponense[J]. Marine Sciences, 2006, 30(7): 5–8. |
| [9] | CHEN J, XIE P.Tissue distributions and seasonal dynamics of the hepatotoxic microcystins-LR and -RR in two freshwater shrimps, Palaemon modestus and Macrobrachium nipponensis, from a large shallow, eutrophic lake of the subtropical China[J]. Toxicon, 2005, 45(5): 615–625. DOI:10.1016/j.toxicon.2005.01.003 |
| [10] | 王志铮, 杨磊, 朱卫东, 等.日本鳗鲡(Anguilla japonica)对日本沼虾(Macrobrachium nipponense)的捕食效应[J]. 海洋与湖沼, 2011, 42(1): 107–113. WANG Z Z, YANG L, ZHU W D, et al.Study of predation habit of Anguilla japonica on Macrobrachium nipponense[J]. Oceanologia et Limnologia Sinica, 2011, 42(1): 107–113. |
| [11] | 刘凯, 段金荣, 徐东坡, 等.太湖秀丽白虾抱卵前后肌肉生化组成和能量密度变动[J]. 云南农业大学学报, 2010, 25(3): 377–382. LIU K, DUAN J R, XU D P, et al.Variation of biochemical components and energy density in muscle of Exopalaemon modestus in breeding Season in Taihu Lake[J]. Journal of Yunnan Agricultural University, 2010, 25(3): 377–382. |
| [12] | 温周瑞, 张大文, 谢平, 等.太湖日本沼虾、秀丽白虾现存量的周年动态研究—以梅梁湾、贡湖湾为例[J]. 长江流域资源与环境, 2011, 20(9): 1035–1039. WEN Z R, ZHANG D W, XIE P, et al.Seasonal dynamics in stock of the freshwater shrimps, Exopalaemon modestus and Macrobrachium nipponense in Meiliang and Gonghu bays, Lake Taihu[J]. Resources and Environment in the Yangtze Basin, 2011, 20(9): 1035–1039. |
| [13] | 刘仕鑫, 高春霞, 田思泉, 等.淀山湖秀丽白虾形态特征对体质量的影响效果[J]. 上海海洋大学学报, 2015, 24(6): 926–933. LIU S X, GAO C X, TIAN S Q, et al.Effects of morphometric traits on body weight for Exopalaemon modestus in Dianshan Lake[J]. Journal of Shanghai Ocean University, 2015, 24(6): 926–933. |
| [14] | 温周瑞, 谢平.太湖日本沼虾与秀丽白虾的食性与食物碳源分析[J]. 长江大学学报(自然科学版), 2013, 10(29): 36–42. WEN Z R, XIE P.Studies on the feeding habits and sources of dietary carbon of Macrobrachium nipponense and Exopalaemon modestus in Taihu Lake[J]. Journal of Yangtze University (Natural Science), 2013, 10(29): 36–42. |
| [15] | OH C W, HARTNOLL R G, NASHM R D M.Population dynamics of the common shrimp, Crangon crangon (L.), in Port Erin Bay, Isle of Man, Irish Sea[J]. ICES Journal of Marine Science, 1999, 56(5): 718–733. DOI:10.1006/jmsc.1999.0501 |
| [16] | 俞存根, 陈小庆, 宋海棠, 等.春季东海海域虾类群落结构及其多样性[J]. 生态学报, 2009, 29(7): 3593–3604. YU C G, CHEN X Q, SONG H T, et al.Analysis on community structure and diversity of shrimps in the East China Sea in spring[J]. Acta Ecologica Sinica, 2009, 29(7): 3593–3604. |
| [17] | 温周瑞, 谢平, 徐军.太湖贡湖湾虾类种类组成与时空分布特征[J]. 湖泊科学, 2011, 23(6): 961–966. DOI:10.18307/2011.0620 WEN Z R, XIE P, XU J.Spatial and temporal patterns of species composition and distribution of shrimp community in the Gonghu Bay, Lake Taihu[J]. Journal of Lake Sciences, 2011, 23(6): 961–966. DOI:10.18307/2011.0620 |
| [18] | 谈奇坤.安徽湖泊虾类的生物学和资源管理[J]. 安徽农业科学, 1991: 80–83. TAN Q K.Biology and the resources management of shrimps in the lakes of Anhui[J]. Journal of Anhui Agricultural Sciences, 1991: 80–83. |
| [19] | 孙建贻, 张道源, 谭德清, 等.洪湖日本沼虾种群生长的研究[J]. 湖泊科学, 1999, 11(2): 149–154. DOI:10.18307/1999.0211 SUN J Y, ZHANG D Y, TAN D Q, et al.Population growth of freshwater shrimp (Macrobrachium nipponensis) in Honghu Lake[J]. Journal of Lake Sciences, 1999, 11(2): 149–154. DOI:10.18307/1999.0211 |
| [20] | 温周瑞, 谢平.太湖秀丽白虾生长研究[J]. 水产科技情报, 2013, 40(3): 139–142. WEN Z R, XIE P.Research on growth of freshwater shrimp, Exopalaemon modestus in Taihu Lake[J]. Fisheries Science & Technology Information, 2013, 40(3): 139–142. |
| [21] | 温周瑞, 谢平.太湖日本沼虾与秀丽白虾摄食节律研究[J]. 水产科技情报, 2013, 40(1): 54–56. WEN Z R, XIE P.Research on Feeding rhythm of the freshwater shrimps, Macrobrachium nipponense and Exopalaemon modestus in Taihu Lake[J]. Fisheries Science & Technology Information, 2013, 40(1): 54–56. |
| [22] | 李强, 田华, 姜民, 等.淀山湖浮游动物群落结构特征及其影响因子[J]. 水生态学杂志, 2015, 36(4): 69–77. LI Q, TIAN H, JIANG M, et al.Community structure of zooplankton and influencing factors in Dianshan Lake of Shanghai[J]. Journal of Hydroecology, 2015, 36(4): 69–77. |
| [23] | 康丽娟.淀山湖沉积物碳、氮、磷分布特征与评价[J]. 长江流域资源与环境, 2012, 21(S1): 105–110. KANG L J.Characteristics and risk evaluation of carbon, nitrogen and phosphorus in sediments of the Dianshan Lake[J]. Resources and Environment in the Yangtze Basin, 2012, 21(S1): 105–110. |
| [24] | 胡雪芹, 王强, 马明睿, 等.淀山湖叶绿素a分布特征及其与浮游植物密度的相关性[J]. 华东师范大学学报(自然科学版), 2012: 149–156. HU X Q, WANG Q, MA M R, et al.Distribution characteristics of chlorophyll a and its relationship with phytoplankton density in Lake Dianshan[J]. Journal of East China Normal University (Natural Science), 2012: 149–156. |
| [25] | 吴阿娜, 朱梦杰, 汤琳, 等.淀山湖蓝藻水华高发期叶绿素a动态及相关环境因子分析[J]. 湖泊科学, 2011, 23(1): 67–72. DOI:10.18307/2011.0111 WU A N, ZHU M J, TANG L, et al.Dynamics of chlorophyll-a and analysis of environmental factors In Lake Dianshan during summer and autumn[J]. Journal of Lake Sciences, 2011, 23(1): 67–72. DOI:10.18307/2011.0111 |
| [26] | 张欢, 谢平, 吴功果, 等.日本沼虾与秀丽白虾的营养生态位[J]. 环境科学研究, 2013, 26(1): 22–26. ZHANG H, XIE P, WU G G, et al.Studies on trophic niches of Macrobrachium nipponensis and Exopalaemon modestus[J]. Research of Environmental Sciences, 2013, 26(1): 22–26. |
| [27] | 严生良, 金德沂.太湖的青虾[J]. 水产科技情报, 1981: 18–19. YAN S L, JIN D Y.The freshwater shrimps, Macrobrachium nipponense in Taihu Lake[J]. Fisheries Science & Technology Information, 1981: 18–19. |
| [28] | 温周瑞, 谢平, 徐军.太湖秀丽白虾繁殖生物学研究[J]. 长江大学学报(自然科学版), 2012, 9(4): 23–27. WEN Z R, XIE P, XU J.Reproductive biology of Exopalaemon modestus in Taihu Lake[J]. Journal of Yangtze University (Natural Science), 2012, 9(4): 23–27. |
2. Collaborative Innovation Center for Distant-water Fisheries, Shanghai 201306, China;
3. National Distant-water Fisheries Engineering Research Center, Shanghai Ocean University, Shanghai 201306, China;
4. The Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai Ocean University, Shanghai 201306, China