 2017,
Vol. 26
2017,
Vol. 26



2. 江苏省海洋水产研究所, 江苏 南通 226007;
3. 上海市浦东新区农业委员会执法大队渔政中队, 上海 201308
杭州湾北邻长江口,西连钱塘江,是典型的喇叭形强潮海湾[1-2],与河口物质交换频繁,拥有丰富的生物资源[3]。杭州湾海域是棘头梅童鱼(Collichthys lucidus)、凤鲚(Coilia mystus)等多种鱼类产卵、索饵场所,群落结构具有典型的季节性特征[4]。随着杭州湾东海大桥、洋山深水港和跨海大桥等工程以及周边人类活动的增多,生境发生变化,势必影响鱼类资源分布、群落结构和多样性变化等[5]。把握鱼类资源的变动是评价渔业资源动态的重要手段,仔稚鱼群落分布除了物种自有属性外,还受到环境因子的影响[6]。因此,研究仔稚鱼分布特征,对掌握了解渔业资源具有重要意义。
目前关于杭州湾鱼类资源的研究多局限于季节性的比较[7-11],缺乏年度间的比较成果。陈渊戈等[12]比较分析了长江口南支和杭州湾北部沿岸碎波带仔稚鱼群落特征,结果显示杭州湾北部碎波带仔稚鱼具有更高的物种丰富度。 闫欣等[8]分析了杭州湾北部水域周年仔稚鱼种类的组成及其生物多样性,发现杭州湾北部水域是多种经济鱼类仔稚鱼的保育场。本研究在进一步开展杭州湾北部水域周年调查的基础上,与闫欣等2013年的调查结果作比较,分析该水域仔稚鱼种类组成和群落结构的年度变化,旨在为杭州湾北部水域鱼类资源保护与利用进一步提供科学依据。
1 材料与方法 1.1 采样区域及方法本研究于2014年6月至2015年5月,在杭州湾北部水域设置10个站点(St.1~St.10,图 1),以渔船(沪捕渔No.49627,功率120马力)于每月大潮前后在各站点采用大型仔稚鱼网(口径1.3 m,网目0.5 mm)进行表层水平拖网。拖网速度控制在2 nmile/h,拖网时间为10 min。使用网口流量计(KC Denmark A/S Model 23.090)记录拖网流量值;同时记录各站位的表层水温、盐度等环境因子数据。所有采集样本均在现场用5%的甲醛溶液固定。

|
图 1 调查站位
Fig. 1 Sampling stations
|
在实验室内将采集到的仔稚鱼在Olympus SZX7解剖镜下鉴定到科、属、种,按KENDALL等[13]的仔稚鱼发育分期标准划分发育阶段,计数并计算密度(ind./m3),并用目测微尺或游标卡尺测量仔稚鱼的体长(SL)。科名按NELSON等[14],学名和中文名按照伍汉霖等[15],同属的种名按英文字母进行排序,并确定生态类型。
1.3 数据分析数据使用SPSS 19.0校验分析两次调查环境因素和仔稚鱼密度的相关显著性。使用PRIMER 5.0软件计算两年仔稚鱼Shannon-Wiener指数(H),均匀度指数(J)以及Margalef丰富度指数(D)。使用Shannon-Wiener指数(H)对仔稚鱼群落多样性进行简单评价。使用Arcgis 10.2软件绘制数量密度分布图。
仔稚鱼密度依据公式:
 (1)
(1)
优势种依据公式:
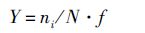 (2)
(2)
式中: Ga 为单位体积海水中仔稚鱼个体数(m3); Na为全网仔稚鱼个体数(尾);S为网口面积(m2 );L为流量计转数;C为流量计校正值(C=0.3)。ni为第i种仔稚鱼物种的物种数;N为调查航次物种总数;f为该物种在该调查航次出现的频率。
2 结果 2.1 水温和盐度 2.1.1 水温水温范围为6.4~27.4 ℃,2015年2月平均水温最低(6.4±0.3 ℃),2014年7月平均水温最高(27.4±0.9 ℃),平均值为16.9±0.6 ℃。本次调查平均水温低于2013年调查的平均水温(图 2)。SPSS差异性统计分析表明两年水温差异性不显著(P>0.05)。

|
图 2 杭州湾北部水域月平均水温变化
Fig. 2 Monthly changes of mean water temperature in the north of Hangzhou Bay
|
平均盐度为14.2±1.6,2015年2月盐度最高为19.9±1.0,8月份最低为4.6±1.3。2014年7—9月梅雨季节时间长,降水量大,导致采样点海水表面盐度下降,7—9月的平均值仅为8.4±1.2,呈现出与2013年相反的下降趋势。杭州湾北部水域盐度变化幅度大,无明显的变化规律(图 3)。SPSS差异性统计分析表明两年盐度差异性不显著(P>0.05)。

|
图 3 杭州湾北部水域的月平均盐度变化
Fig. 3 Monthly changes of mean salinity in the north of Hangzhou Bay
|
2014年6月至2015年5月共拖网120网次,采集仔稚鱼96 118尾,平均密度0.79ind./m3,隶属于23科55种。个体数、种类数和平均密度均小于2013年采集到的仔稚鱼。两年的共有种类有46种,且两年调查仔稚鱼物种种类组成差异不大(表 1)。

|
表 1 2014年6月-2015年5月杭州湾北部水域采集的仔稚鱼种类组成 Tab.1 Species compositions of fish larvae and juveniles collected in the north of Hangzhou Bay |
本次调查所采集的髭缟虾虎鱼(Tridentiger barbatus)数量最多,共39 524尾(41.12%),其次为棘头梅童鱼(19.80%)、拉氏狼牙虾虎鱼(Odontamblyopus lacepedii,14.14%)和凤鲚(9.79%)。而2013年占百分比前四种的仔稚鱼为:髭缟虾虎鱼(64.55%)、棘头梅童鱼(13.09%)、凤鲚(9.24%)和斑尾刺虾虎鱼(Acanthogobius ommaturus,2.72%)。本次调查中拉氏狼牙虾虎鱼的数量显著增多,2013年没有采集到此次百分比排在第八位的前鳞(Liza affinis)。从表 2可以看出,两年间前10位种类基本类同。

|
表 2 渔获仔稚鱼主要种类比较 Tab.2 Comparison of dominant species of fish larvae and juveniles between two years |
按照仔稚鱼生态类型的不同,可将杭州湾北部水域仔稚鱼划分为河口性鱼类(Es)、海洋性鱼类(Ma)、洄游性鱼类(Mi)和淡水性鱼类(Fr)四大类。两年采集到的仔稚鱼均以河口性种类和海洋性种类为主,且河口性鱼类所占比例均多于海洋性种类(表 3)。

|
表 3 杭州湾北部水域仔稚鱼生态类型组成 Tab.3 Ecological types of fish larvae and juveniles distribution in the north of Hangzhou Bay |
2013年1月、2月和2015年2月调查未采集到仔稚鱼。相对于2015年3月采集到的均为卵黄囊期仔鱼,2013年3月卵黄囊期仔鱼也达到92.98%。2014—2015前弯曲期仔鱼则集中出现在4—11月,所占比例均大于2013年同期,其中4月达到最大值(95.99%)。弯曲期仔鱼出现在2014年6月至12月,最大值出现在11月为45.52%,与2013年有相同趋势。后弯曲期仔鱼均出现在5—12月,2013年最大值出现在10月为44.44%,而2014—2015年出现在11月为29.42%。稚鱼在4—12月出现,以12月采集量最多。由此可见,两年仔稚鱼发育阶段组成趋势基本一致,前弯曲期仔鱼占优势,其次为弯曲期仔鱼(图 4)。

|
图 4 各月仔稚鱼发育阶段组成
Fig. 4 Development stages distribution of fish larvae and juveniles
|
仔稚鱼的水平分布可以显示站位间的差异。本次调查中远岸站位密度高于近岸站位,其中St.4密度最高(1.29 ind./m3),St.9密度最低(0.23 ind./m3)。2013年调查则显示仔稚鱼分布比较离散,St.8密度最高(3.66 ind./m3),St.10密度最低(0.44 ind./m3)。SPSS配对样本t校验差异性统计分析表明,两年各站位仔稚鱼的平均密度差异显著(P<0.05)。

|
图 5 杭州湾北部水域仔稚鱼平均密度分布
Fig. 5 Average density of fish larvae and juveniles of each station in the north of Hangzhou Bay
|
总体来看两年仔稚鱼密度月变化无显著差异(P>0.05),具有相同的变动趋势,均以6月份仔稚鱼数量最多,呈明显峰值,这是由于6月份髭缟虾虎鱼和棘头梅童鱼大量出现所致。也正是因2014年6月髭缟虾虎鱼的数量远比2013年6月要少,导致其仔稚鱼的密度(4.82 ind./m3)远小于2013年6月(13.4 ind./m3)(图 6)。

|
图 6 杭州湾北部水域仔稚鱼月份变化
Fig. 6 Monthly changes of abundance and density of fish larvae and juveniles in the north of Hangzhou Bay
|
选取Y值大于0.02为优势种,本次调查中优势种为髭缟虾虎鱼、棘头梅童鱼和凤鲚。髭缟虾虎鱼为最优势种,优势度为0.12,与2013年的调查相吻合,出现频率、数量均低于2013年;棘头梅童鱼优势度较2013年有所增高;优势种凤鲚在两年调查的百分比、出现频率和优势度趋于相近(表 4)。SPSS配对样本t检验差异性统计分析表明,两年调查的优势种分布差异不显著(P>0.05)。

|
表 4 杭州湾北部水域仔稚鱼优势种的比较 Tab.4 Comparison of dominant species of fish larvae and juveniles in the north of Hangzhou Bay |
对仔稚鱼种类数、密度与水温、盐度分别对应进行双变量相关性分析,结果显示:两年种类数与水温极显著相关。
2013年和2014—2015年仔稚鱼种类数与水温极显著相关,密度与水温不显著相关。2014—2015年仔稚鱼种类数与盐度呈显著负相关(P<0.05),相关系数R为-0.665,密度与盐度无显著相关关系;2013年种类数、密度与盐度不显著相关(表 5)[8]。

|
表 5 种类数、密度与环境因子的关系 Tab.5 Pearson correlations between environmental factors and species number,density |
从月间变化来看,仔稚鱼群落Shannon-Wiener指数(H′)和Pielou均匀度指数(J′)变化趋势一致,后者变化幅度相对较小。本研究由于2月无渔获物,因此3个指数均为0。Margalef丰度指数(D)变化较大,1月物种数很少,丰富度指数极低,3—6月呈持续上升趋势,6—8月丰富度指数维持较高水平,9—12月呈急剧下降趋势(图 7)。比较两年的Margalef丰富度指数(D),2014年8月仔稚鱼种类数较多(26种),使得D值维持在较高水平;而2013年8月仔稚鱼种类数仅有17种,导致D值急剧下降。2013年Margalef丰富度指数(D)的最大值出现在7月。
2013年Shannon-Wiener指数(H′)和Pielou均匀度指数(J′)最大值分别出现在10月和12月,二者在6—8月均较低;而本次调查Shannon-Wiener指数(H’)和Pielou均匀度指数(J’)最大值分别出现在7月和9月,二者在6—8月均较高。

|
图 7 杭州湾北部水域仔稚鱼群落多样性指数变化
Fig. 7 Variation of diversity index of fish larvae and juveniles community in the north Hangzhou Bay
|
综上所述,多样性分析表明两年多样性指数季节性变化明显,均在夏季较高、冬季较低。本次调查夏季Margalef丰富度指数(D)、Shannon-Wiener指数(H′)和Pielou均匀度指数(J′)均高于2013年夏季。
3 讨论 3.1 环境因子对仔稚鱼分布的影响温度是鱼类繁殖及生长发育的直接控制因子之一[16]。鱼类产卵活动和生长与水温息息相关,不同鱼类适应温度和感觉温差的能力各不相同,海水鱼类感觉温差的能力达到0.1 ℃,鱼类会选择或避开一定的温度范围[17]。RAMOS等[18]对Lima河口鱼类的分析表明温度的季节变动对仔稚鱼群落时间变化起主导作用。本研究对仔稚鱼种类数、密度(表 4)与水温进行双变量相关性分析结果也表明了温度对仔稚鱼种类数有一定的影响。
潮汐强弱、降水等直接影响海湾水域的盐度变化[19],从而对仔稚鱼空间分布特征产生影响[20]。SUSANA等[21]对葡萄牙海岸的研究发现盐度影响仔稚鱼群落在河口不同盐度区域的分布;李建生等[6]对长江口水域仔稚鱼研究也指出盐度影响着仔稚鱼群落的分布。本研究调查期间虽因降雨引起表层盐度变化,但并不影响仔稚鱼的水平分布(P>0.05),这可能是调查水域的选取划分,并不足以突破盐度影响阈值,达到显性状态。本次调查种类数与盐度显著相关(P<0.05),表明盐度在某种程度上可以影响仔稚鱼群落的组成。
3.2 种类组成与多样性HALDORSON等[22]对Alaskan海湾仔稚鱼研究表明河口、海湾种群结构较单一,主要优势种优势地位特别突出,在种群结构中起到主导地位。本研究杭州湾各种鱼类丰度变化较大,优势种对总丰度的贡献最大,2013年和2014-2015年优势种占总捕获量的86.88%和70.71%,优势种中尤其以髭缟虾虎鱼的贡献最大,为该水域的代表种。
两年调查期间杭州湾北部水域6月、8月共采集到42种仔稚鱼,多于宋秀凯等[23]在莱州湾6月、8月的调查结果;黄凤鹏等[24]在胶州湾进行周年调查仅采集到10种仔稚鱼。三者的优势种差别明显,莱州湾的优势种为凤鲚、(Liza haematocheila)和斑(Konosirus punctatus);胶州湾的优势种为刺虾虎鱼(Acanthogobius sp.)、斑和玉筋鱼(Ammodytes personatus)。由此可见,海湾鱼类仔稚鱼的种类组成、多样性与其地理位置、类型、大小有一定的关系。而且,使用网具的不同也对研究结果的差异起到一定的作用。
蒋玫等2003—2005年对洋山深水港区(与本研究站位接近)调查发现优势种为中华侧带小公鱼(Stolephorus chinensis)、和虾虎鱼科属Gobiidae spp.,且仔稚鱼数量明显低于施工前的2001年,优势种也有较大差别[25];周永东等2008年对浙江中北部沿岸水域(其Ⅰ区站位与本研究站位接近)调查发现凤鲚、棘头梅童鱼主要在Ⅰ区杭州湾区域,并且为优势种[26];本研究与周永东等调查结果优势种一致。杭州湾北部水域由于港口码头、跨海大桥等施工影响导致2003—2005年仔稚鱼群落结构发生了很大的改变,一些无法适应环境改变的鱼类亲本远离或者生殖能力下降,使得优势种群体改变,但是随着时间的推移,这种影响逐渐减小[25]。本研究比较2013年的结果,发现棘头梅童鱼优势度有所增高,凤鲚的优势度比较稳定。说明近年随着环境保护力度的加强,局部生态结构趋于稳定。
沈新强等杭州湾水域近20年间生物多样性研究表明,杭州湾水域生物多样性变化可归纳为两个阶段,第一阶段是上世纪90年代末至本世纪初,杭州湾生物群落多样性指标呈普遍下降态势;第二阶段是2000年至今,随着政府对水体生态环境保护力度加大和更多保护方案的实施,杭州湾生物多样性水平得到不同程度水平上的恢复[27]。比较两年的研究结果,仔稚鱼群落的种类组成、发育阶段、月份间的差异并不显著,仅各站位的仔稚鱼分布差异显著。多样性分析发现两年多样性指数季节性变化明显,均在夏季较高、冬季较低,而且本次调查夏季丰富度指数(D)、Shannon-Wiener指数(H′)和Pielou均匀度指数(J′)均高于2013年夏季,说明近两年杭州湾北部水域仔稚鱼群落仍处于一个稳定发展的阶段。
| [1] | 中国海湾志编纂委员会. 中国海湾志[M]. 北京: 海洋出版社, 1998. Editorial Board of China Bay Survey. Survey of China bays[M]. Beijing: China Ocean Press, 1998. |
| [2] | 陈吉余. 中国河口海岸研究与实践[M]. 北京: 高等教育出版社, 2007. CHEN J Y. The review and prospect of estuarine and coastal of China[M]. Beijing: Higher Education Press, 2007. |
| [3] | 贾海波, 邵君波, 曹柳燕.杭州湾海域生态环境的变化及其发展趋势分析[J]. 环境污染与防治, 2014, 36(3): 14–19. JIA H B, SHAO J B, CAO L Y.Analysis of the changes and development trend of ecological environment in Hangzhou Bay[J]. Environmental Pollution and Control, 2014, 36(3): 14–19. |
| [4] | 王淼, 洪波, 张玉平, 等.夏、冬季杭州湾北部游泳动物群落结构[J]. 广东海洋大学学报, 2015, 35(3): 56–62. WANG M, HONG B, ZHANG Y P, et al.Community structure of nektons in northern Hangzhou Bay in summer and winter[J]. Journal of Guangdong Ocean University, 2015, 35(3): 56–62. |
| [5] | 周燕, 龙华, 余骏.应用大型底栖动物污染指数评价杭州湾潮间带环境质量[J]. 海洋环境科学, 2009, 28(5): 473–477. ZHOU Y, LONG H, YU J.Assessment on environmental quality in tideland of Hangzhou Bay by macrozoobenthos pollution index[J]. Marine Environmental Science, 2009, 28(5): 473–477. |
| [6] | 李建生, 胡芬, 程家骅.长江口水域春季鱼卵仔鱼分布及其与温度盐度的关系[J]. 海洋科学, 2007, 31(4): 16–21. LI J S, HU F, CHENG J H.Distributions of fish eggs and juveniles and relations between surface layer water temperature and salinity of the Yangtze estuary water area in spring[J]. Marine Science, 2007, 31(4): 16–21. |
| [7] | 魏永杰, 王晓波, 张海波, 等.2004~2010年夏季杭州湾鱼类浮游生物种类组成与数量分布[J]. 台湾海峡, 2012, 31(4): 501–508. WEI Y J, WANG X B, ZHANG H B, et al.Ichthyoplankton species composition and distribution in Hangzhou Bay in summer during 2004-2010[J]. Journal of Oceanography in Taiwan Strait, 2012, 31(4): 501–508. |
| [8] | 闫欣, 钟俊生, 王明星, 等.杭州湾北部水域仔稚鱼种类组成和多样性研究[J]. 上海海洋大学学报, 2014, 23(5): 765–773. YAN X, ZHONG J S, WANG M X, et al.Study on the species composition and diversity of fish larvae and juveniles in the north of Hangzhou Bay[J]. Journal of Shanghai Ocean University, 2014, 23(5): 765–773. |
| [9] | 谢旭, 俞存根, 周青松, 等.杭州湾海域春、 秋季鱼类种类组成和数量分布[J]. 海洋与湖沼, 2013, 44(3): 656–663. XIE X, YU C G, ZHOU Q S, et al.Species composition and quantitative distribution of fishes in the Hangzhou Bay during spring and autumn[J]. Oceanologia et Limnologia Sinica, 2013, 44(3): 656–663. |
| [10] | 范明生, 王海明, 蔡如星, 等.杭州湾期间带生态学研究─Ⅰ. 种类组成与分布[J]. 海洋学研究, 1996, 4(4): 1–11. FAN M S, WANG H M, CAI R X, et al.Studies on interdital zone ecology of Hangzhou Bay─Ⅰ. composition and distribution of species[J]. Journal of Marine Science, 1996, 4(4): 1–11. |
| [11] | 周永东, 薛利建, 徐开达.舟山近海凤鲚Coilia mystus(Linnaeus)的生物学特性研究[J]. 现代渔业信息, 2004, 19(8): 19–21. ZHOU Y D, XUE L J, XU K D.A study on biological characteristics of Coilia mystus(Linnaeus) offshore of Zhoushan[J]. Modern Fisheries Information, 2004, 19(8): 19–21. |
| [12] | 陈渊戈, 张宇, 钟俊生, 等.长江口南支和杭州湾北岸碎波带水域仔稚鱼群聚的比较[J]. 上海海洋大学学报, 2011, 20(5): 688–696. CHEN Y G, ZHANG Y, ZHONG J S, et al.Comparison in fish larvae and juvenile assemblages between the surf zones of south branch of Yangtze River estuary and north coast of Hangzhou Bay[J]. Journal of Shanghai Ocean University, 2011, 20(5): 688–696. |
| [13] | KENDALL A W Jr. Early life history stages of fishes and their characters[M]//MOSER H G, RICHARDS W J, COHEN D M, et al. Ontogeny and Systematics of Fishes. Lawrence: The American Society of Ichthyologists Herpetologists, Spec Publ, 1984. |
| [14] | NELSON J S. Fishes of the World[M]. 4th ed. New York: John Wiley & Sons, 2006. |
| [15] | 伍汉霖, 邵广昭, 賴春福. 拉汉世界鱼类系统名典[M]. 基隆: 水产出版社, 1999. WU H L, SHAO G Z, LAI C F. Latin-Chinese dictionary of fish names by classification system[M]. Keelung: The Sueichan Press, 1999. |
| [16] | 刘栋, 朱鑫华, 沙学绅.长江口春季鱼类浮游生物群落结构与环境因子的关系[J]. 海洋科学集刊, 2002: 169–179. LIU D, ZHU X, SHA X S.Relationship between community structure of ichthyoplankton and environmental factors in the estuary of Changjiang River in spring[J]. Studia Marina Sinica, 2002: 169–179. |
| [17] | 林昭进, 詹海刚.大亚湾核电站温排水对邻近水域鱼卵、 仔鱼的影响[J]. 热带海洋, 2000, 19(1): 44–51. LIN Z J, ZHAN H G.Effects of thermal effluent on fish eggs and larvae in waters near Daya Bay Nuclear Plant[J]. Tropic Oceanology, 2000, 19(1): 44–51. |
| [18] | RAMOS S, COWEN R K, RÉ R K, et al.Temporal and spatial distributions of larval fish assemblages in the Lima estuary (Portugal)[J]. Estuarine, Coastal and Shelf Science, 2006, 66(1/2): 303–314. |
| [19] | 孔亚珍, 贺松林, 丁平兴, 等.长江口盐度的时空变化特征及其指示意义[J]. 海洋学报, 2004, 26(4): 9–18. SUN Y Z, HE S L, DING X P, et al.Characteristics of temporal and spatial variation of salinity and their indicating significance in the Changjiang Estuary[J]. Acta Oceanologica Sinica, 2004, 26(4): 9–18. |
| [20] | ISLAM M S, HIBINO M, TANAKA M.Distribution and diets of larval and juvenile fishes: influence of salinity gradient and turbidity maximum in a temperate estuary in upper Ariake Bay, Japan[J]. Estuarine, Coastal and Shelf Science, 2006, 68(1/2): 62–74. |
| [21] | FRANÇA S, COSTA M J, CABRAL H N.Inter-and intra-estuarine fish assemblage variability patterns along the Portuguese coast[J]. Estuarine, Coastal and Shelf Science, 2011, 91(2): 262–271. DOI:10.1016/j.ecss.2010.10.035 |
| [22] | HALDORSON L, PRITCHETT M, STERRITT D, et al.Abundance patterns of marine fish larvae during spring in a southeastern Alaskan bay[J]. Fishery Bulletin, 1993, 91: 36–44. |
| [23] | 宋秀凯, 刘爱英, 杨艳艳, 等.莱州湾鱼卵、仔稚鱼数量分布及其与环境因子相关关系研究[J]. 海洋与湖沼, 2010, 41(3): 378–385. SONG X K, LIU A Y, YANG Y Y, et al.Distributing of ichthyoplankton and the correlation with environmental factors in Laizhou Bay[J]. Oceanologia et Limnologia Sinica, 2010, 41(3): 378–385. |
| [24] | 黄凤鹏, 黄景洲, 杨玉玲, 等.胶州湾鱼卵、仔鱼和稚鱼的分布[J]. 海洋科学进展, 2007, 25(4): 468–473. HUANG F P, HUANG J Z, YANG Y L, et al.Distributions of Fish Eggs and Larval Fish in the Jiaozhou Bay[J]. Advances in Marine Science, 2007, 25(4): 468–473. |
| [25] | 蒋玫, 王云龙, 林钦, 等.洋山深水港水域鱼卵仔鱼分布特征[J]. 海洋环境科学, 2008, 27(1): 43–46. JIANG M, WANG Y L, LIN Q, et al.Distribution characteristics of fish eggs and larvae around Yangshan Port in Hangzhou Bay[J]. Marine Enviromental Science, 2008, 27(1): 43–46. |
| [26] | 周永东, 金海卫, 蒋日进, 等.浙江中北部沿岸春、夏季鱼卵和仔稚鱼种类组成与数量分布[J]. 水产学报, 2011, 36(6): 880–889. ZHOU Y D, JIN H W, JIANG R J, et al.The category composition and abundance distributions of ichthyoplankton along the north-central coast of Zhejiang Province in spring and summer[J]. Journal of Fisheries of China, 2011, 36(6): 880–889. |
| [27] | 沈新强, 周永东.长江口、 杭州湾海域渔业资源增殖放流与效果评估[J]. 渔业现代化, 2007, 34(4): 54–57. SHEN X Q, ZHOU Y D.Enhencing release and effect assessment of fisheries resource in the Yangze River Estuary and Hangzhou Bay[J]. Fishery Modernization, 2007, 34(4): 54–57. |
2. Institute of Oceanology and Marine Fisheries Jiangsu, Nantong 226007, Jiangsu, China;
3. Law Enforcement Team, Agriculture Commission of Pudong District, Shanghai 201308, China