 2015, Vol. 24
2015, Vol. 24


2. 上海海洋大学国际海洋研究中心, 上海 201306;
3. 国家海洋局东海海洋预报中心, 上海 200081
水温是海域生态环境的重要影响因子,对海域初级生产力及生态动力学过程的影响尤为明显。浮游植物是海洋中的主要初级生产者,每一种浮游植物均有自己的最佳生长温度与可适应生长温度范围。如中肋骨条藻(Skeletonema costatuma)具有较广的温度适应性(15~ 25℃)[1];东海原甲藻(Prorocentrum donghaiense)适宜的温度范围是15~30℃,最适温度为22~28℃,低于9 ℃和高于33 ℃时不能进行光合作用[2],也有学者认为东海原甲藻对水温的耐受范围的下限和上限分别为10.2和30.6 ℃[3];海洋原甲藻(Prorocentrum micans Ehrenb)最大增殖速率发生在温度为25 ℃,在温度18~28 ℃范围内适宜生长[4]。浮游动物的生长同样也有自己的适宜水温范围[5, 6]。滨海电厂的温排水通常比环境水温高8~12 ℃左右[6, 7, 8],温排水在排水口附近扩展,大幅度的温升将对局部海域内浮游生物的生长产生显著影响。在低温季节促进生长,高温季节严重抑制生长。温排水的生态影响还包括卷载和余氯对浮游生物的杀伤[6, 9]。
象山港是一狭长型海湾(图1a),湾口至湾中部的水深约为15~30 m,湾顶的水深在10 m以下(图1b)。湾中部和湾顶分别有乌沙山和国华两大滨海电厂排放循环冷却水,相对封闭的地形造成水交换不畅,不利于温排水的扩散。2006年1月6日,象山港宁海峡山网箱养殖区附近海域甚至因温排水影响发生了罕见的大面积冬季赤潮,最大面积扩大至14 km2,厚度5~6 m,直至1月18日,赤潮方才最终消亡。可见,滨海电厂温排水在影响海域水温时空分布的同时,对海域浮游植物生物量的变化产生了深远影响。温排水的热冲击加之卷载和余氯胁迫效应,对浮游生物乃至整个海域的生态和环境均有显著影响[5, 6, 7, 8, 9, 10]。
对于滨海电厂温排水的生态和环境影响,以往的研究主要集中在温排水的温升及其影响范围的观测和数值模拟[8, 11],冷却水余热和余氯对浮游植物[7]和浮游动物的影响[5, 6],温排水对围格生态系统中浮游生物的影响[12]以及围格内水质模型的参数率定[13]等,少有应用浮游生物生态动力学数值模型开展的温排水生态影响研究。海洋数值模式现已成为海洋动力学研究领域的一个重要手段,本文选用发展较为成熟、在国内外应用广泛的海洋数值模式ECOM-si[8, 14] (Estuarine Coastal Ocean Model semi-implicit),并耦合一个太阳辐射和海面热通量计算模块,采用N2P2ZD型生态模型,数值模拟评估了象山港海域两大滨海电厂温排水对浮游生物的生态影响程度。
1 研究方法 1.1 研究区域图1a为研究区域和数值模式的计算网格。数值模式水动力和水温计算时间序列的验证均详见文献[8, 14]。电厂冷却水的取排水口位置见图1a所示,采用深取浅排的方式取排水,数值模式中亦按此方式设置。

|
图1 研究区域及数值模式网格剖分(a),港区水深(m)分布及2008年观测站点 Fig.1 Location of study area and meshes of numerical model (a),topography of the Xiangshan bay and observed sites of 2008 |
象山港海域的浮游植物种类较多,但以往的历次调查均显示,浮游植物以硅藻门为主,其次为甲藻门[15, 16, 17, 18],另有少量的金藻、蓝藻和绿藻等。常见优势种为琼氏圆筛藻(Coscinodiscus jonesianus)[11, 16, 17, 18]、中肋骨条藻等[18]。本文所用的生态模型移植自有限体积河口海岸模型(FVCOM)的可变生态模型(Flexible Biological Module,FBM)。根据象山港海域的浮游植物种类和营养盐分布特征,所用N2P2ZD型生态模型考虑以下6个主要的生态变量:溶解无机氮(DIN)、磷酸盐(PO4-P)、硅藻类(DIA)、甲藻类(FLA)、浮游动物(ZOO)和碎屑(DET),物质(生态参量)输运和扩散方程采用与盐度计算相同的数值解法,具体计算公式、生态模型(图2)变量间的关系以及生态模型与外部环境的关系见文献 [19]。

|
图2 N2P2ZD生态模型示意图 Fig.2 Scheme of N2P2ZD ecosystem model |
在象山港海域中,硅藻和甲藻是浮游植物生物量中占有最大比重的两个类群。中肋骨条藻( Skeletonema costatum)和东海原甲藻(Prorocentrum donghaiense,即具齿原甲藻Prorocentrum dentatum)是东海北部近海海域的主要藻种[20]。同时,这两个藻种也是近年来本海域最常爆发的赤潮生物种,且两个种群在生长过程中存在种间竞争现象[1]。 因此,本文以本海域典型的硅藻种中肋骨条藻和典型甲藻种东海原甲藻的生物生态学研究成果为基础,来进行硅藻和甲藻类的参数设置。本文所用浮游生态数值模式已在临近的长江口海域得到成功运用,除水温限制方案外,其它生态过程及其参数设置不再详述,具体的生态过程控制方程及其参数设置详见文献 [19]。
1.3 水温影响生态变量的计算方案本文所采用浮游植物生长温度限制方案为式(1)所示:
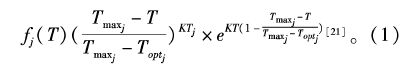
图3所示备选最优水温限制方案为:(1)FBM(Flexible biological model,海洋模式FVCOM所含的生态模块)中最佳温度方案[22]:f(T)=e-0.069|T-Topt|;(2)Q10方案[23]:f (T) =Sb(Q10)(T-Tb) /10,Q10=2,Tb=10 ℃;(3)Epply方案[24]:f (T ) =Vpexp(0.0633T) ;(4)本文参照文献[21]公式,对参数取值修正为:KT=3.5,Topt =23 ℃,Tmax =31 ℃。其它方案中,Topt也取23 ℃。Sb和Vp在此处为便于绘图,分别取为0.2和0.1。5个数据点为陈炳章等所测得的具齿原甲藻随温度变化的比生长率[25]。经对比分析,本文所采用李彦宾的方案[18]结果与陈炳章等[25]的实测比生长率最接近,相比传统的Q10、指数方案或单纯的最佳温度方案(FBM)为优。本文所采用的方案能体现生长率在最佳温度附近先增后降的趋势,且有较大温度区间位于最佳温度附近。FBM方案的最佳温度仅为一个点,且不受最高允许生长温度的控制,水温超过最佳温度较高时,生长率下降缓慢。Q10方案和Epply方案本质上比较接近,区别在于分别以10 ℃和0 ℃的生长率为基准。因为本海域的水温在夏季经常出现超过藻类最佳生长温度的情况,这两种方法接近乘方或指数增长的特性不适用本海域。本文的甲藻类温度限制生长函数采用上述方案及参数取值。硅藻采用相同的函数,仅最佳生长温度和最高可生长温度分别调整为Topt =25 ℃,Tmax =37 ℃。

|
图3 最优水温方案 Fig.3 Functions of optimal water temperature |
浮游动物生长受水温的限制条件为[22]:g=gme-0.069|T-Topt(Z)|。式中,g为浮游动物摄食率,gm为最大摄食率,T为水温,Topt(Z)为最佳摄食温度,文中取为20 ℃。水温高于或低于20 ℃时,摄食率均小于gm。碎屑的矿化速率受水温影响较大,文中碎屑矿化过程随温度的变化呈指数增长[22]:r=dn0e(0.025·T)。式中,r为矿化率,dn0为0摄氏度时的矿化率。
1.4 生态模型初始和边界条件设置生态模型中,主要关注水温对生物量的影响,因此不考虑港区周边河流排放营养盐的影响。营养盐的初始场中,DIN和PO4-P的浓度值选取文献[15]所给出的年平均值,且垂向均匀,DIN(三态氮浓度之和)为46.43 mmol/m3,PO4-P为1.16 mmol/m3。浮游生物和碎屑的初始场采用冷启动,取统一初始值0.2 mmol N/m3。边界条件也设为辐射边界条件。数值模型的物质守恒性是测试模式可行性的重要指标。文中“目标海域”设定为121.75°E以西的象山港海域。采用营养盐均值初始场方案,开边界采用辐射边界条件,使计算域基本保持高水平、等营养盐状态且突出水温因素的影响。
光照条件方面,采用年平均的透明度值1.2 m[15]。如此,可突出水温因素在温排水生态影响研究中的作用。国华电厂和乌沙山电厂温排水量分别为40 m3/s和52 m3/s,其中国华电厂7月和8月的温排水量为80 m3/s,冷却水的温升幅度为取水温度加8 ℃。电厂温排水中的余氯和卷载效应对浮游生物群落影响较大,0.2 mg /L的氯可以直接杀死冷却水中 60%~80%的藻类[10],而电厂卷载影响对浮游生物数量损伤率在10%~30%左右[26];电厂温排水系统卷载对海水浮游藻类数量的损伤率为11.98%~27.08%,而卷载效应对浮游动物的数量损伤率为31%~90% [27]。氯对冷却水中水生生物的杀伤强度与余氯浓度水平和作用时间(间接或连续添加)有关。在缺少象山港电厂余氯与卷载效应对浮游生物杀伤作用确切研究结果的情况下,本文生态模式中,温排水条件下考虑卷载和余氯的影响时,设定将进入取水口作为电厂冷却水的水体中50%的浮游植物和浮游动物杀死,即温排水仅保留50%的浮游生物量,死亡的50%浮游生物量变为等量的碎屑。
2 结果 2.1 月平均气象条件下的水温分布和温升规律月平均气象条件下数值模拟的水温分布规律为:2月份的月平均水温在9 ℃左右,乌沙山排水口附近仅在沿岸一带出现水温9.5 ℃的温升带,而港底的国华电厂附近则有大面积的水温达9.5 ℃的温升带。5月份水温在20 ℃左右,港底的水温高于港口。8月份的仍呈港底高于港口,平均水温30 ℃左右,港口29.5 ℃左右,港底可达31.0 ℃左右。11月份,由于港底处水深较浅更易失热,港口及外海水深大、水温高,出现了港口水温高于港底的情形,温排水口附近仍保持较高的水温。温排水的存在有助于缓解春季和秋冬季港底水温的下降,一定程度上有助于提高该时期海域的生产力。
本文中,所有水平分布图均为时间间隔12小时输出的月或年平均值。月平均气象条件作用下年均温升如图4所示,乌沙山电厂排水口附近因流速大、水交换条件较好,仅在沿岸形成一个高于0.4 ℃的狭长温升区,其范围仍小于国华电厂排水口附近高于0.5 ℃的温升区。乌沙山电厂附近大于0.5 ℃的温升区范围与国华电厂大于1 ℃的温升范围相当。国华电厂的温升范围大于乌沙山电厂,且对铁港的影响大于黄墩港。

|
图4 月平均气象条件作用下年均温升分布(℃) Fig.4 Distribution of annual temperature rise under monthly average meteorological data(℃) |
2008年1月、4月、7月和11月分别开展了象山港内多站位(图1b)观测。观测所得13个站位的表层叶绿素a浓度和表层水温分别如图5a、5b所示。上述4个(季节)中,象山港平均叶绿素a浓度分别为0.36±0.10、1.53±0.53、4.35±2.20、0.93±0.45 mg/m3;相应的季节平均水温为10.3±1.7、14.9±0.9、28.2±1.2、17.3±1.9 ℃。1月,平均表温仅10℃左右,整个港区生物量均很低,且港口附近水温高于港底,靠近国华电厂排水口的11、13号站点(图1b)水温明显高于港底的10、12号站点,但因冬季基础水温低、散热快,温升对浮游植物生物量无明显影响。4月,港底水温高于港口,且13号站点及位于铁港的11、12号站点叶绿素a浓度明显高于黄墩港的9、10号站点。7月,平均水温超过28 ℃,稍高于模式中设定的硅藻类最佳生长温度(25 ℃),仍远低于最高可生长温度(37 ℃),港内叶绿素a浓度达到4次观测中的最高浓度,且受温排水影响大的11、13号站点叶绿素a浓度远低于港底的10、12号站点。11月,港口水温高于港底,全港平均水温虽高于4月,但由于太阳高度角小于4月等原因,叶绿素a浓度低于4月,且离国华电厂排水口最近的13号站点的叶绿素a浓度高于周边其它港底站点。

|
图5 2008年象山港海域多站点观测所得叶绿素a浓度(a)和水温(b) Fig.5 Observed concentration of Chl-a (a) and water temperature (b) in Xiangshan Bay,2008 |

|
表1 象山港海域叶绿素a浓度历史观测数据 Tab. 1 Historical observed data of Chl-a in Xiangshan Bay |
本文在多年平均气象条件下运行生态模型4年,待各生态参量值(除营养盐)变化规律性稳定后,取第5年的数值模拟结果用于分析研究。风应力、气压、气温、相对湿度、云量等月平均气象数据均数字化自《渤海、黄海、东海海洋图集》。
目标海域生态变量的季节变化规律如图6所示。在月平均气象条件下,其浮游植物的季节变化基本规律如下:一般地,12月初开始,象山港海域的水温降至15 ℃以下,直至3月底前水温均维持在15 ℃以下,这一时期目标海域的生物量一直维持在低水平。4月中下旬后,水温上升至15 ℃以上,生物量快速增长,5月底左右,硅藻类和甲藻类的生物量均达到最高,分别达6.0 mg Chl-a/m3和3.0 mg Chl-a/m3,总浮游植物生物量最大值超过9.0 mg Chl-a/m3,接近文献所述的丰水期浮游植物生物量[15]。甲藻类的适宜水温相对硅藻低,在温排水影响下,春季水华会较硅藻更早到来。随着7月份水温超过25 ℃,硅藻类和甲藻类生物量均显著降低,特别是甲藻类(代表非耐高温种),在7月后始终维持在低生物量水平,8月底水温值达到最高后,硅藻类生物量从4.0 mg Chl-a/m3逐渐恢复到6.0 mg Chl-a/m3。硅藻类在9到10月仍维持高生物量的原因主要是所定义的此类浮游植物较耐高温,其次是由于甲藻类不耐高温、生物量低,在这一时期不构成对硅藻类营养盐的竞争。11月后,硅藻类生物量也随着水温下降而显著下降。至次年3月底前浮游植物生物量维持在1.0~1.5 mg Chl-a/m3,接近文献所述的枯水期浮游植物生物量[15]。表1所示为近30年文献所述的各月份象山港叶绿素a浓度。将表1数据点绘制于图6中(图中圆圈),可见月平均气象条件下的数值模拟浮游植物总生物量(图6,PHY)与大多数历史数据点符合良好,且体现了象山港海域硅藻生物量对甲藻生物量的绝对优势,与刘子琳等所得象山港浮游植物细胞丰度中硅藻占绝对优势(94%)、甲藻和蓝藻分别仅为4%和2%的结果接近[29]。
浮游动物和碎屑生物量在浮游植物低值期也维持低生物量,最低可至0.5 mg C/m3,随着水温升高,浮游植物量上升后,浮游动物和碎屑均快速增长,碎屑的高值期较浮游植物高值期后滞5~15天,浮游动物高值期较浮游植物高值期后滞30天左右。浮游动物和碎屑生物量与总浮游植物量类似,均具有双峰高值期,在春季水华期最高,秋季水华期后次高。碎屑量与浮游植物和浮游动物生物量直接相关,浮游动物量除了饵料来源(浮游植物和碎屑)的限制,还主要受水温的限制(本研究不考虑浮游动物被摄食)。

|
图6
无温排水时目标海域各参量季节变化 其中PHY、DIA和FLA为左坐标,其余项为右坐标,下同;图中圆圈为表1中的叶绿素a数据 Fig.6 Seasonal variation of ecological parameters without discharge of thermal water in the bay PHY、DIA and FLA in left Y-axis,other parameters in right Y-axis. Circles were drawn from the data in Table1. |
综上所述,周年变化的太阳辐射(光照)以及与之相关的水温是影响象山港水域浮游生态系统变化的主要因素。关于数值模式中太阳辐射的验证详见文献[8]。
生物量水平分布有以下几个特点:(1)浮游植物和浮游动物量由于卷载和余氯杀伤作用,在排水口附近维持一个低值区,而碎屑量则在排水口附近保持一个高值区;(2)生物量的分布态势基本为港底附近高于港口附近,仅8月份的浮游动物由于港底水温较高,港口的生物量高于港底;(3)生物量低值期,其水平分布的差异也相对较小,生物量高值期生物量水平分布差异也较大,最大在港底可达港口的2倍左右。上述特点中的(2)、(3)与2008年(图5)观测所得叶绿素a分布规律相符。
3 讨论 3.1 水温季节变化对象山港海域浮游植物生物量的影响纵然浮游植物现存生物量是光照、透明度、营养盐和水温等众多环境因素影响和浮游植物本身种类和丰度综合的结果,将图5a、5b中的实测所得2008年四季的叶绿素a和水温数据绘制点图(图7)后,还是可以发现,因象山港内呈狭长型,各部分海域的水温有差异,而叶绿素a浓度明显受到水温的影响。图7中添加由公式(1)计算浮游植物生长水温限制系数曲线(象山港海域的浮游植物以硅藻为主,故取Topt =25 ℃,Tmax=37 ℃),发现上述叶绿素a浓度点分布与水温限制曲线两侧,两者有较好的对应。由于水温限制系数的最大值在最佳生长温度25 ℃时为1,故将叶绿素a浓度值除以观测所得最高叶绿素a值可进行归一化,将归一化后数据计算与图7所得曲线的误差平方和,当KT取7.5时误差平方和为2.58,当KT取14.5时误差平方和最小,为1.54。

|
图7 2008年象山港叶绿素a浓度(图中点,左纵坐标)与浮游植物生长水温限制系数(图中实线与虚线,右纵坐标)的关系 注解 Fig.7 Observed concentration of Chl.a (dots, in left Y-axia) and temperature limiting coefficient of phytoplankton growth (solid and dashed lines, in right Y-axia) in Xiangshan Bay,2008 |
为了便于比较,各生物变量差异的物质量单位统一为mg C/m3。有温排水条件下与无温排水的生态变量差值见图8a和图8b(生态变量差异相对于无温排水条件下的%)。整个冬季的生态变量绝对差值较小,仅碎屑量由于卷载效应和余氯影响略有升高,冬季和初春的碎屑量相对差值比率可高达40%(此时,碎屑量处于相对低值水平),浮游植物生物量的下降在5%左右,浮游动物生物量下降达10%左右。定义浮游植物量(硅藻类和甲藻类之和)、浮游动物量和碎屑量之和为一个新的变量,即总有机体量(图中记为Bio_all)。总有机体在冬季基本与无温排水条件下持平。春季,有温排水条件下较无温排水条件下升温快,较早达到最大生物量值,因此春季的生物量相对无温排水条件下略增5%。春末,有温排水条件下的生物量出现一个正值期,此为无温排水条件下升温滞后所致。此后,初夏6月底之前,有温排水条件下浮游植物量又较无温排水条件下高,但此时的浮游动物量(模式中设定浮游动物最佳生长温度为20℃)由于水温的升高有15%左右的下降。6月底至9月初,随着水温的持续升高,有温排水条件下浮游生物量持续下降,浮游植物生物量下降15%左右,浮游动物量由于水温升高和食物来源的减少下降可达30%,此阶段的差值使全年基本维持正值的碎屑生物量也出现了最高达到15%的下降。9月底后,随着环境气温和水温的下降,温排水的增温效应显现,可使浮游植物生物量较无温排水条件下增加5%左右。12月起,增温效应开始小于卷载和余氯杀伤的效应,浮游植物又呈现小幅下降。

|
图8
有无温排水条件下生态变量的周年差异 (a.绝对生物量变幅,mg C/m3;b.相对生物量变幅,%) Fig.8 Annual difference of ecological parameters between cases with and without discharge of thermal water (a for absolute biomass,mg C/m3 and b relative for biomass,%) |
浮游植物生物量(图9)在排水口附近均有大幅下降(设卷载和余氯杀伤率50%),且国华附近大于乌沙山附近。2月份,排水口外围海域浮游植物生物量下降约0.5~1.0 mg C/m3(或1%~2%);5月份达到了8.0 mg C/m3以上(或5%以上);8月份生物量下降最大,可达30.0 ~50.0 mg C/m3(或15%~25%);11月份,除排水口附近因卷载和余氯影响浮游植物生物量下降外,外围海域因为温排水造成的温升效应,反而使浮游植物生物量有1.0~2.0 mg C/m3(或1~2%)的上升。除了甲藻类生物量最高为5月份外,其它季节生物量的增减都以硅藻类为主。5月份排水口外围海域的硅藻类生物量的下降仅为2.0 mg C/m3左右,而甲藻类下降可达4.0 mg C/m3。

|
图9 有无温排水条件下各季节浮游植物绝对生物量差异的水平分布(mg C/m3) Fig.9 Horizontal distribution of difference of absolute phytoplankton biomass(mg C/m3)between cases with and without discharge of thermal water during four seasons |
浮游动物生物量在2月份小幅下降,约0.6~0.8 mg C/m3;5月份,温排水影响区的浮游动物量下降达2.0 mg C/m3以上,卷载和余氯影响区外的浮游动物量反而因为升温早而使生物量略有增加;8月份达4.0 mg C/m3左右;11月份,下降量在2.0~3.0 mg C/m3左右。碎屑生物量始终在排水口附近保持高值。2月份,外围海域的碎屑量增加2 mg C/m3左右;5月份,外围海域碎屑增加量在3.0~4.0 mg C/m3以上;8月份,高水温的影响显著,使浮游植物生物量大幅下降,反而使碎屑量下降了10.0 mg C/m3;11月,碎屑量又回归至约3.0~4.0 mg C/m3以上的增量。总有机体量(图10)2月份平均值较无温排水时小幅增加,约0.5~1.0mg C/m3;5月份,总有机体量在两个排水口附近有较大幅度降低,达5.0~10.0 mg C/m3;8月份,总有机体量下降幅度最大,达30.0~80.0 mg C/m3;11月份,总有机体量则有2.0~4.0 mg C/m3的小幅上升。

|
图10 有无温排水条件下各季节总有机体量差异的水平分布(mg C/m3) Fig.10 Horizontal distribution of difference of total organic matter(mg C/m3)between cases with and without discharge of thermal water during four seasons |
如前所述,温排水对海域生态的影响,不能仅考虑其对浮游植物和浮游动物生物量的影响,还应考虑碎屑量以及总有机体量的增减来综合评价。其次,需要考虑海域的生态功能,不同的生态功能对浮游植物、浮游动物和碎屑的利用不同。最后,还应考虑浮游植物或动物优势种类的变迁而造成它们作为饵料的适口性的差异,以及水华或高生物量发生期的提前或滞后所造成的影响等。
3.3 电厂综合效应(卷载、余氯)对浮游生物生态系统的影响考虑卷载和余氯的影响,本文中是指进入取水口作为电厂冷却水的水体中,50%的浮游植物和浮游动物被杀死,并变为碎屑。温排水合并卷载和余氯效应终年在排水口附近使浮游植物生物量下降(图11)。对于排水口外围海域,乌沙山电厂温排水的影响较小,这与该电厂地处港区中段,流速大、扩散性好有关。国华电厂对港底浮游生态系统的影响较大,且对铁港海域的影响大于黄墩港。在铁港的浮游植物量年平均下降可达6.0 mg C/m3,浮游动物量下降达到5.0 mg C/m3,碎屑生物量则上升2.0~3.0 mg C/m3,造成总有机体量的年平均下降量在9.0 mg C/m3左右。

|
图11 考虑卷载和余氯影响下浮游植物生物量下降的年平均值 Fig.11 Horizontal distribution of annual average reduction of phytoplankton biomass with the influence of mechanical entanglement and residual chlorine |
卷载和余氯杀伤使碎屑生物量增加、使浮游植物和浮游动物生物量下降。由于浮游动物的生长速度较浮游植物慢,浮游动物群体耐受卷载和余氯杀伤的生物量恢复速率明显较浮游植物慢,因此,卷载和余氯杀伤对浮游动物的影响最大。不考虑卷载和余氯影响时,温升使浮游动物生物量春、秋、冬季的大部分时间均呈现5%~15%左右的增长,仅在夏季高温期有最大15%左右的降幅,这一降幅较考虑卷载和余氯杀伤影响时的20%~25%小5%~10%。卷载和余氯对浮游生物量的杀伤作用是终年存在的,而温升效应的作用在不同季节则有正有负。
4 结论本文应用海洋数值模式ECOM-si耦合一个可用于实时计算的太阳辐射强度和海面热通量计算模块,采用N2P2ZD型生态模型,研究评估了象山港海域的浮游生物量受港内两大滨海电厂(国华电厂和乌沙山电厂)温排水的影响程度。数值模式的模拟结果表明,温排水及其卷载和余氯杀伤效应对海域的浮游生物量有显著影响。在春季和初夏等增温季节,温排水的存在可使藻华的发生期提前,甚至发生冬季赤潮过程;夏季高温期,水温超过多数藻类的最佳生长温度,则可使浮游植物生物量下降,水温超过30 ℃的机率增大从而可降低赤潮发生的风险;在秋季降温期和冬季低温期,水温低于最佳生长温度,温排水的存在有助于缓解港底水温的下降,一定程度上有助于提高该时期海域的初级生产力。由于硅甲藻类适宜水温的差异,温排水不仅影响浮游生物量也影响海域的浮游生物种类构成。
卷载和余氯杀伤对浮游生态系统有一定的影响,绝对生物量方面对浮游植物影响较大,相对生物量方面对浮游动物影响较大,但其影响程度小于温排水造成的温升效应。除了季节变化上有增有减外,浮游植物和浮游动物量年平均水平由于温排水的温升、卷载和余氯杀伤效应而降低,碎屑量随之上升,总体上弥补了海域水体总有机体含量的降低,但海域的年均总有机体变化量仍为负值。国华电厂温排水的影响大于乌沙山电厂。无论从温升范围和浮游生物量变化幅度上,国华电厂对铁港的影响都大于黄墩港。
本文仅为数值模拟的初步研究结论,因篇幅限制,对于营养盐循环受水温的影响[16]、不同粒级浮游植物[31]受温升的差异等内容未能涉及。今后需要应用更多的观测、更为复杂的海洋浮游生态系统数值模型来对滨海电厂温排水的生态影响进行深入研究。
| [1] | 王宗灵,李瑞香,朱明远,等.半连续培养下东海原甲藻和中肋骨条藻种群生长过程与种间竞争研究[J].海洋科学进展, 2006, 24(4):495-503.WANG Z L, LI R X, ZHU M Y, et al. Study on population growth processes and interspecific competition of Prorocentrum donghaiense and Skeletonema costatum in semi-continuous dilution experiments[J]. Advances in Marine Science, 2006, 24(4):495-503. |
| [2] | 邓光,耿亚洪,胡鸿钧,等.几种环境因子对高生物量赤潮甲藻东海原甲藻光合作用的影响[J].海洋科学, 2009, 33(12):34-39.DENG G, GENG Y H, HU H J, et al. Effects of environmental factors on photosynthesis of a high biomass bloom forming species Prorocentrum donghaiense[J]. Marine Sciences, 2009, 33(12):34-39. |
| [3] | 陈艳拢,赵冬至,杨建洪,等.赤潮藻类温度生态幅的定量表达模型研究[J].海洋学报, 2009, 31(5):156-161.CHEN Y L, ZHAO D Z, YANG J H, et al. The quantitative study on ecological amplitude of sea temperature of algae[J]. Acta Oceanologica Sinica, 2009, 31(5):156-161. |
| [4] | 王正方,张庆,吕海燕.温度、盐度、光照强度和pH对海洋原甲藻增长的效应[J].海洋与湖沼, 2001, 32(1):15-18.WANG Z F, ZHANG Q, LV H Y. Effects of temperature, salinity, light and pH on the growth of red tide organisms Prorocentrum micans[J]. Oceanologia et Limnologia Sinica, 2001, 32(1):15-18. |
| [5] | 徐晓群,曾江宁,曾淦宁,等.滨海电厂温排水对浮游动物分布的影响[J].生态学杂志, 2008, 27(6):933-939.XU X Q, ZENG J N, ZENG G N, et al. Effects of thermal water discharged from coastal power plant on distribution of zooplankton[J]. Chinese Journal of Ecology, 2008, 27(6):933-939. |
| [6] | 江志兵,曾江宁,陈全震,等.滨海电厂冷却水余热和余氯对中华哲水蚤的影响[J].应用生态学报, 2008, 19(6):1401-1406.JIANG Z B, ZENG J N, CHEN Q Z, et al. Effects of residual heat and chlorine in cooling water from coastal power plant on Calanus sinicus[J]. Chinese Journal of Applied Ecology, 2008, 19(6):1401-1406. |
| [7] | 江志兵,曾江宁,陈全震,等.滨海电厂冷却系统温升和加氯对浮游植物联合作用的模拟研究[J].环境科学学报, 2009, 29(2):413-419.JIANG Z B, ZENG J N, CHEN Q Z, et al. A simulation of the combined effects of temperature increase and chlorination on phytoplankton in a coastal power plant cooling system[J]. Acta Scientiae Circumstantiae, 2009, 29(2):413-419. |
| [8] | 林军,章守宇,龚甫贤.象山港海洋牧场规划区选址评估的数值模拟研究:滨海电厂温排水温升的影响[J].上海海洋大学学报, 2012, 21(5):816-824.LIN J, ZHANG S Y, Gong F X. Numerical simulation study on site selection evaluation of marine ranching zone in Xiangshan Bay:Effects of thermal water discharged from coastal power plants[J]. Journal of Shanghai Ocean University, 2012, 21(5):816-824. |
| [9] | 林昭进,詹海刚.大亚湾核电站温排水对邻近水域鱼卵、仔鱼的影响[J].热带海洋学报, 2000, 19(1):44-51.LIN Z J, ZHAN H G. Effects of thermal effluent on fish eggs and larvae in waters near Daya bay nuclear plant[J]. Tropic Oceanology, 2000, 19(1):44-51. |
| [10] | 曾江宁,陈全震,郑平,等.余氯对水生生物的影响[J].生态学报, 2005, 25(10):2717-2724.ZENG J N, CHEN Q Z, ZHENG P, et al. Advanced in effect of residual chlorine on hydrobios[J]. Acta Ecologica Sinica, 2005, 25(10):2717-2724. |
| [11] | 张惠荣,赵瀛,杨红,等.象山港滨海电厂温排水温升特征及影响效应研究[J].上海海洋大学学报, 2013, 22(2):274-281.ZHANG H R, ZHAO Y, YANG H, et al. Study on the temperature rise characteristics and influence effects of thermal discharge from coastal power plant in Xiangshan Bay[J]. Journal of Shanghai Ocean University, 2013, 22(2):274-281. |
| [12] | 蔡泽富,杨红,焦俊鹏,等.温排水对围隔生态系统各粒级海洋浮游生物的影响[J].水产学报, 2011, 35(8):1240-1246.CAI Z F, YANG H, JIAO J P, et al. Effects of thermal effluent on different size-fractionated marine plankton in the mesocosm ecosystem[J]. Journal of Fisheries of China, 2011, 35(8):1240-1246. |
| [13] | 杨红,丁骏,王春峰,等.象山港围隔生态系水质模型研究[J].海洋科学, 2012, 36(7):14-22.YANG H, DING J, WANG C F, et al. Study on the water quality model of the mesocosm ecosystem in the Xiangshan Bay[J]. Marine Sciences, 2012, 36(7):14-22. |
| [14] | 林军,章守宇,龚甫贤.象山港海洋牧场规划区选址评估的数值模拟研究:水动力条件和颗粒物滞留时间[J].上海海洋大学学报, 2012, 21(3):452-459.LIN J, ZHANG S Y, GONG F X. Numerical study on site selection evaluation of the marine ranching planning zone in Xiangshan Bay:hydrodynamic conditions and particle residence time[J]. Journal of Shanghai Ocean University, 2012, 21(3):452-459. |
| [15] | 黄秀清,王金辉,蒋晓山.象山港海洋环境容量及污染物总量控制研究[M].北京:海洋出版社, 2008.HUANG X Q, WANG J H, JIANG X S. Study on total quantity control of marine environmental capacity and pollutant in Xiangshan[M]. Beijing:China Ocean Press, 2008. |
| [16] | 江志兵,陈全震,寿鹿,等.象山港人工鱼礁区的网采浮游植物群落组成及其与环境因子的关系[J].生态学报, 2012, 32(18):5813-5824.JIANG Z B, CHEN Q Z, SHOU L, et al. Community composition of net-phytoplankton and its relationship with the environmental factors at artificial reef area in Xiangshan Bay[J]. Acta Ecologica Sinica, 2012, 32(18):5813-5824. |
| [17] | 陈海峰,王云龙,廖勇,等.象山港人工鱼礁区浮游植物群落特征[J].海洋渔业, 2013, 35(4):396-404.CHEN H F, WANG Y L, LIAO Y, et al. Community characteristics of phytoplankton in artificial reef areas of Xiangshan Bay[J]. Marine Fisheries, 2013, 35(4):396-404. |
| [18] | 张海波,蔡燕红,项有堂.象山港水域浮游植物与赤潮生物种群动态研究[J].海洋通报, 2005, 24(1):92-96.ZHANG H B, CAI Y H, XIANG Y T. Study on the phytoplankton and HAB in Xiangshan Bay[J]. Marine Science Bulletin, 2005, 24(1):92-96. |
| [19] | 林军.长江口外海域浮游植物生态动力学模型研究[D].上海:华东师范大学, 2011.LIN J. A modeling study of the phytoplankton dynamics off the Changjiang Estuary[D]. Shanghai:East China Normal University, 2011. |
| [20] | 张秀芳,刘永健.东海原甲藻Prorocentrum donghaiense Lu生物学研究进展[J].生态环境, 2007, 16(3):1053-1057.ZHANG X F, LIU Y J. Advances in the biological study of dinoflagellate Prorocentrum donghaiense Lu[J]. Ecology and Environment, 2007, 16(3):1053-1057. |
| [21] | 李雁宾.长江口及邻近海域季节性赤潮生消过程控制机理研究[D].青岛:中国海洋大学, 2008.LI Y B. The study of the seasonal occurrence mechanism of HABs in the Changjiang Estuary and its adjacent sea[D]. Qingdao:Ocean University of China, 2008. |
| [22] | CHEN C S, BEARDSLEY R C, COWLES G. FVCOM User Manual[M]. 2006. |
| [23] | JI R B, DAVIS C, CHEN C S, et al. Influence of local and external processes on the annual nitrogen cycle and primary productivity on Georges Bank:A 3-D biological-physical modeling study[J]. Journal of Marine Systems, 2008, 73(1/2):31-47. |
| [24] | 刘浩,尹宝树.渤海生态动力过程的模型研究Ⅰ.模型描述[J].海洋学报, 2006, 28(6):21-31.LIU H, YIN B S. Model study on Bohai ecosystemⅠ. Model description and primary productivity[J]. Acta Oceanologica Sinica, 2006, 28(6):21-31. |
| [25] | 陈炳章,王宗灵,朱明远,等.温度、盐度对具齿原甲藻生长的影响及其与中肋骨条藻的比较[J].海洋科学进展, 2005, 23(1):60-64.CHEN B Z, WANG Z L, ZHU M Y, et al. Effects of temperature and salinity on growth of Prorocentrum dentatum and comparisons between growths of Prorocentrum dentatum and Skeletonema costatum[J]. Advances in Marine Science, 2005, 23(1):60-64. |
| [26] | 何琴燕,任敏,蔡燕红,等.滨海电厂温排水对生态环境影响研究进展[J].海洋开发与管理, 2012, (7):84-88.HE Q Y, REN M, CAI Y H, et al. Research progress on the effect of temperature and drainage on the ecological environment of coastal power plant[J]. Ocean Development and Management, 2012, (7):84-88. |
| [27] | 盛连喜,王显久,李多元,等.青岛电厂卷载效应对浮游生物损伤研究[J].东北师大学报自然科学版, 1994, (2):83-89.SHENG L X, WANG X J, LI D Y, et al. The study of plankton damaged in Qingdao power plant[J]. Journal of Northeast Normal University, 1994, (2):83-89. |
| [28] | 刘子琳,宁修仁,史君贤,等.象山港对虾增殖放流区浮游植物现存量和初级生产力[J].海洋学报, 1997, 19(6):109-115.LIU Z L, NING X R, SHI J X, et al. Primary productivity and phytoplankton of value-added release area of biomassin in Xiangshan bay[J]. Acta Oceanologica Sinica, 1997, 19(6):109-115. |
| [29] | 刘子琳,蔡昱明,宁修仁.象山港中、西部秋季浮游植物粒径分级、叶绿素a和初级生产力[J].东海海洋, 1998, 16(3):18-24.LIU Z L, CAI Y M, NING X R. The distribution of chlorophyll a and primary productivity in the middle and west of Xiangshan Bay[J]. Donghai Marine Science, 1998, 16(3):18-24. |
| [30] | 王菲菲,章守宇,林军.象山港海洋牧场规划区叶绿素a分布特征研究[J].上海海洋大学学报, 2013, 22(2):266-273.WANG F F, ZHANG S Y, LIN J. Study of chlorophyll a distribution in marine ranching planning area of Xiangshan Bay[J]. Journal of Shanghai Ocean University, 2013, 22(2):266-273. |
| [31] | 栾青杉,孙军. 2005年夏季长江口水域浮游植物群集特征及其与环境因子的关系[J].生态学报, 2010, 30(18):4967-4975.LUAN Q S, SUN J. Feature of phytoplankton assemblages in Yangtze River estuary and its relationship with environmental factors in summer 2005[J]. Acta Ecologica Sinica, 2010, 30(18):4967-4975. |
2. International center for marine studies, Shanghai Ocean University, Shanghai, 201306, China;
3. East China Sea Marine Forecasting Center, State Oceanic Administration, Shanghai 200081, China