 2015, Vol. 24
2015, Vol. 24



2. 华东师范大学生命学院, 上海 200062
鱼类处于水生态系统食物网的较高营养级,在水生态系统初级生产力调控、营养盐再生与循环及能量流动中起着非常重要的作用 [1]。鱼类食性信息是生态系统、食物网研究及生物学描述中令人关注的内容[2]。对鱼类食性研究最早采用的是胃肠内含物法,随着科学技术的发展,研究鱼类食性的新方法、新理论不断产生和发展[3]。不同的技术各有利弊,与新方法相比较,传统胃含物法具有更加直观、即时性等特点,但也存在食物种类难以辨认的局限。同时,传统胃肠内含物法选择何种指数衡量饵料重要性方面还存在争议[3],如CORTS推荐相对重要性百分比指数(IRI%)作为鱼类食性分析比较的标准测度工具(standardized measure)[4, 5],但HANSSON认为IRI%存在不足[2]。
刀鲚(Coilia nasus)隶属于鲱形目(Clupeiformes),鳀科(Engraulidae),鲚属(Coilia),它及其同物异名种[6]为长江中下游湖泊如洞庭湖、鄱阳湖、巢湖、太湖的优势类群[7]。目前,对刀鲚的研究报道主要集中在分类、洄游生殖、遗传进化、营养及渔业资源等方面[8],而对其食性的研究报道极少[9, 10, 11]。刀鲚为淀山湖的优势种[7],在淀山湖生态系统的结构和功能中可能起着比较重要的作用。本文根据2009年-2010年在淀山湖所采集的刀鲚样品,对其食物组成进行定量分析,同时比较了刀鲚食物组成的季节变化,研究结果有助于我们加深对刀鲚食性的认识,为淀山湖湖泊与渔业管理以及生态修复提供依据。
1 材料与方法 1.1 样品采集淀山湖(北纬31°04′~31°12′,东经120°54′~121°01′)是上海最大的天然淡水湖泊,于2009年8月至2010年7月设置6个站点逐月采集刀鲚(2010年1月未采集),即张家库(S1)、苏沪交界(S2)、水上乐园(S3)、急水港(S4)、淀峰(S5)、南湖中心(S6)[7](图1)。共采集刀鲚样品387尾,其中有效胃内含物样本371尾,刀鲚体长分布范围为5.0~26.8 cm,体质量分布范围为1.1~53.4 g。

|
图1 上海淀山湖鱼类调查采样点分布 Fig.1 The fish sampling stations in Dianshan Lake,Shanghai |
样品采集后放入低温保存箱带回实验室,测量其生物学指标如体长(0.1 cm)和体质量(0.1 g),并迅速解剖摘取胃样,将摘取的胃样置于10%福尔马林溶液固定保存。食物鉴定、计数、称重等参照叶佳林等[10]的方法进行。
1.3 数据处理摄食强度用空胃率和饱满指数来表示,公式如下[10]:
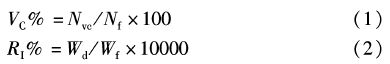
采用5个指数描述刀鲚各种食物类群的重要性,公式如下[12, 13]:
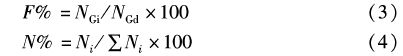
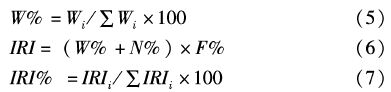
用香农多样性指数(Shannon-wiener指数)表示刀鲚食物多样性,公式如下[7]:

采用组平均聚类法[7]对N%、W%、F%和IRI%数据进行聚类分析,探讨各种指数在衡量不同食物种类对刀鲚重要性方面的差异,并采用N%和W%数据进行季节聚类分析,探讨刀鲚食物数量和重量组成的季节差异。聚类分析中,以d=1-r(r为Pearson相关系数)作为相异性指标,将d=0.3作为存在显著差异的依据[7]。
2 结果 2.1 刀鲚摄食强度刀鲚全年都有摄食活动,但摄食强度却存在明显变化,统计显示,空胃率季节间差异显著(X2=7.962,df=3,P=0.047),秋季空胃率最高,达17.65%,而其他季节则低于10%,饱满指数季节间差异显著(X2=11.652,df=3,P=0.009),春、夏两季刀鲚摄食强度较高,秋、冬两季摄食强度较低(图2)。

|
图2 刀鲚饱满指数和空胃率的季节变化 Fig.2 Seasonal variation of stomach fullness index and percentage of empty stomachs of Coilia nasus |
刀鲚食物由30小类、13大类组成,包括寡毛类、多毛类、轮虫、枝角类、桡足类、浮游植物、端足类、水生昆虫、虾类、鱼类等(表1)。

|
表1 淀山湖刀鲚的食物组成 Tab. 1 Diet composition of Coilia nasus in Dianshan Lake |
基于小类群食物组成数据进行的聚类分析表明,在衡量食物组成成分对刀鲚食性的重要性方面,各指数存在一定的差异,重量百分比(W%)单独聚为一类,其他3个指标(N%、IRI%和F%)聚为一类(图3),基于大类群数据聚类分析结果与此一致。

|
图3 基于食物小类群的刀鲚4种食物重要性指数聚类分析 Fig.3 Dendrogram of 4 indices describing the importance of each small group of diet to Coilia nasus |
N%、F%、IRI% 3个指标均显示,桡足类、枝角类在刀鲚食物组成中占绝对优势,枝角类的N%最高(55.88%),其中象鼻溞(34.8%)、秀体溞(9.12%)较多;桡足类的N%占第2位(39.67%),其中剑水蚤(13.15%)、哲水蚤(8.59%)较多。枝角类的F%最高(81.18%),其中象鼻溞较高(54.03%);桡足类的F%其次,其中剑水蚤(26.88%)、猛水蚤(14.52%)及哲水蚤(16.40%)的比例较高。桡足类(48.84%)和枝角类(46.39%)的IRI%相差不大,其中象鼻溞秀体溞、剑水蚤、哲水蚤的比例较大。W%显示虾类所占比列最高(37.62%),其次为桡足类(29.98%)和鱼类(22%),见表1。
2.3 刀鲚食物组成的季节性变化刀鲚食物数量组成没有显著的季节差异(图4),4个季节均以桡足类和枝角类占绝对优势,两者的贡献率在90%以上(表2)。

|
图4 淀山湖刀鲚食物数量组成季节聚类分析 Fig.4 Dendrogram of seasonal numerical composition of C.nasus’ diet in Dianshan Lake |

|
表2 刀鲚食物类群数量百分比(N%)的季节变化 Tab. 2 Seasonal variations of food composition in terms of numerical percentage (N%) of Coilia nasus |
刀鲚食物重量组成存在显著的季节变化,其中春季和冬季聚为一类,夏季和秋季聚为一类(图5)。春季(53.41%)和冬季(61.32%)均以桡足类比例较大,其次为虾类、鱼类、枝角类,且所占比例相似。夏季(57.79%)和秋季(84.82%)均以虾类的重量百分比占据优势,而桡足类和枝角类在食谱中的比例均在10%以下,另外夏季摄食较多的还有鱼类(30.29%),见表3。

|
图5 淀山湖刀鲚食物重量组成季节聚类分析 Fig.5 Dendrogram of seasonal gravimeric composition of C.nasus’ diet in Dianshan Lake |

|
表3 刀鲚食物类群重量百分比(W%)的季节性变化 Tab. 3 Seasonal variations of food composition in terms of gravimeric percentage (W%) of Coilia nasus |
刀鲚食物香农多样性指数春季最大(1.88),表明春季食物种类来源最为丰富,其次为冬季(1.54)和夏季(1.48),基本持平,而秋季最低(0.77),显示秋季食物来源最为单调。
3 讨论鱼类摄食与其所栖息水域的食物链位置、摄食场所、生活史等有重要关系[14]。一般认为,鱼类摄食行为与水温有较大关系,温度过高或过低通常会使鱼类减少或停止摄食[15]。冬季饵料生物量的减少也会导致鱼类在越冬期很少摄食[16]。此次调查与这些研究结果一致,淀山湖刀鲚在全年都有摄食活动,在冬季摄食强度最低,而随开春之后温度的变化,摄食强度逐渐增强,到夏季达到最高峰(淀山湖夏季温度最高、浮游动物生物量最高[17]),秋季,摄食强度又明显下降,直到冬季又达到最低值(图1)。许多研究也表明,鱼类的空胃率与平均饱满系数变化趋势相反[18],淀山湖刀鲚亦如此。
单一指数如N%、W%和F%可以从不同的角度洞悉捕食者的摄食习性,如数量测度主要提供摄食行为方面的信息,重量测度反映食物的营养价值[19],出现率代表种群尺度的食物习性[20],但这些单一指数在描述食物重要性时各有优缺点,数量测度方法容易高估个体小但数量大的饵料的重要性,而重量测度则容易高估大型饵料的重要性[13]。本研究显示,淀山湖刀鲚的饵料生物达13大类30小类。不同食性指数衡量食物类群对刀鲚的重要性有很大差别,桡足类和枝角类N%达95%以上,而W% 约占40%;鱼虾的N%仅有0.05%,但其W%却接近60%。为了克服单一指数的局限性,一些学者提出了综合性指数如相对重要性指数(IRI)[12]及其百分比指数(IRI%)[4],认为能较真实地反映捕食者的食性。对于综合性指数如IRI%,不同的学者有不同的看法[3],有的学者认为这类指数能够防止信息丢失、减少单一指数造成的误差[4, 21],但也有一些学者认为这类综合性指数降低了精确性、没有增加信息等而批评甚至反对使用这类指数[2, 13, 19]。聚类分析表明,淀山湖刀鲚食性IRI%、N%和F%聚为一类,W%单独聚为一类,说明IRI%未能提供新的信息,因此结合N%和W%指数即可较为全面地解释淀山湖刀鲚的食性。
不同湖泊刀鲚食物组成的数量百分比显示(表4),太湖刀鲚摄食数量比例最高的是桡足类、其次为枝角类[10],巢湖刀鲚和太湖鲚鱼摄食比例最高的枝角类、其次为桡足类[9, 11, 22]。淀山湖刀鲚的食性与后两者的研究较为接近,摄食数量比例最高的为枝角类、其次为桡足类。对桡足类而言,其对剑水蚤、哲水蚤的摄食数量比例较高,而对许水蚤的摄食可以基本忽略。一些调查发现许多鱼类对象鼻溞表现出较低的选择[23],而淀山湖刀鲚对象鼻溞的摄食数量比例很高,而对个体较大的网纹溞、裸腹溞等喜好程度较低,这与优化摄食理论的预测(鱼类捕食对浮游动物有明显的选择性,通常在同等能耗下优先选择个体较大的浮游动物[24])有所差别,可能与淀山湖浮游动物组成成分和密度有关,淀山湖水体浮游动物群落中象鼻溞、剑水蚤的优势度高,成为绝对优势种或类群,而裸腹溞、网纹溞、华哲水蚤等优势度低[25, 26]。淀山湖水体轮虫生物量和密度都高于枝角类、桡足类[25],但刀鲚食物中轮虫所占比例很低,特别是对轮虫优势种—龟甲轮虫的摄食数量比例较低,一方面可能与物种个体大小有关,另一方面可能与鱼类间捕食竞争有关,对其他鱼类而言,刀鲚更倾向于个体较大的桡足类和枝角类[27, 28]。另外,夏季刀鲚对子陵吻鰕虎鱼摄食加强,可能与其大小、种群数量、活动能力有关,子陵吻鰕虎鱼是淀山湖优势种群,夏季繁殖,生物量高[7]。刀鲚摄食的鱼虾数量比例很低,可能与虾和鱼作为刀鲚食物资源的可获得性不高有关[10]。从重量百分比来看,淀山湖刀鲚摄食较多虾类、桡足类和鱼类,与太湖刀鲚较为接近[10],同时与巢湖刀鲚亦较为相似,主要摄食浮游动物和鱼类[29]。

|
表4 不同湖泊刀鲚主要食物组成数量百分比 Tab. 4 The main food composition in terms of numerical percentage (N%) of fish species of genus Coilia in different lakes |
研究表明浮游动物食性鱼类对维持湖泊生态系统结构和功能的稳定起着重要作用,其对浮游动物的捕食能抑制湖泊中浮游动物生物量[30]。本文结果显示淀山湖刀鲚全年都在摄食,其食物成分中浮游动物数量比例在95%以上,特别是夏季、秋季,其浮游动物比例在99%以上,且主要捕食淀山湖浮游动物优势种,而对浮游植物很少摄食。刀鲚是淀山湖的绝对优势鱼类,其数量比例达到50%以上,而肉食性鱼类很少,如鲌类仅占2.7%[7]。根据“下行效应”,淀山湖刀鲚可能会间接促进湖泊中浮游植物的增长。因此,可以结合经典生物操纵和非经典生物操纵技术对淀山湖进行水质管理和生态修复。一方面,通过合理增加肉食性鱼类如鲌类等的比例,增加对刀鲚的捕食压力,抑制淀山湖刀鲚生物量,降低刀鲚对浮游动物特别是其优势种的捕食压力;另一方面,增殖放流可逃避被鱼食性鱼类捕食的大规格鲢、鳙,增加对大型浮游植物如蓝藻群体的摄食压力。通过这两种生物操纵技术的结合使用,可能有助于降低淀山湖浮游植物暴发的几率。
| [1] | SARVALA J, HELMINEN H, AUVINEN H. Portrait of a flourishing freshwater fishery:Pyhäjärvi, a lake in SW-Finland[J]. Boreal Environment Research, 1998, 3(4):329-345. |
| [2] | HANSSON S. Methods of studying fish feeding:a comment[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1998, 55(12):2706-2707. |
| [3] | 薛莹,金显仕.鱼类食性和食物网研究评述[J].海洋水产研究, 2003, 24(2):76-87.XUE Y, JIN X S. Review of the study on feeding habits of fishes and food webs[J]. Marine Fisheries Research, 2003, 24(2):76-87. |
| [4] | CORTÉS E. A critical review of methods of studying fish feeding based on analysis of stomach contents:application to elasmobranch fishes[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1997, 54(3):726-738. |
| [5] | CORTÉS E. Methods of studying fish feeding:reply[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1998, 55(12):2708-2708. |
| [6] | 周晓犊,杨金权,唐文乔,等.基于线粒体CO I基因DNA条形码的中国鲚属物种有效性分析[J].动物分类学报,2010, 35(4):819-826.ZHOU X D, YANG J Q, TANG W Q, et al. Species validities analyses of Chinese coilia fishes based on the DNA Barcoding on the Mitochondrial COI gene Sequences[J]. Acta Zootaxonomica Sinica, 2010, 35(4):819-826. |
| [7] | HU Z J, WANG S Q, WU H, et al. Temporal and spatial variation of fish assemblages in Dianshan Lake, Shanghai, China[J]. Chinese Journal of Oceanology and Limnology, 2014, 32(4):799-809. |
| [8] | 马良骁.我国刀鲚研究进展[J].渔业致富指南, 2014(22):16-17.MA L X. Advances on tapertail anchovy Coilia ectenes in China[J]. Fishery Guide to be Rich, 2014(22):16-17. |
| [9] | 刘恩生,鲍传和,吴林坤,等.太湖新银鱼、鲚鱼的食性比较及相互影响分析[J].湖泊科学, 2007, 19(1):103-110.LIU E S, BAO C H, WU L K, et al. Comparison of food composition and analysis on mutual effects between Neosalanx tangkahkeii taihuensis Chen and Coilia ecten taihuensis Yen et Lin in Lake Taihu[J]. Journal of Lake Sciences, 2007, 19(1):103-110. |
| [10] | 叶佳林,刘正文,王卫民,等.太湖梅梁湾刀鲚与间下(鱼箴)食性的比较[J].湖泊科学, 2007, 19(2):218-222.YE J L, LIU Z W, WANG W M, et al. Comparative study on the feeding habits of Coilia ectenes and Hyporhamphus intermedius in the Meiliang Bay of Lake Taihu[J]. Journal of Lake Sciences, 2007, 19(2):218-222. |
| [11] | 鲍传和,刘恩生,伍昌俊,等.巢湖刀鲚食物组成及渔获量变化的初步分析[J].淡水渔业, 2008, 38(6):19-22.BAO C H, LIU E S, WU C J, et al. Preliminary analysis on diet composition of Coilia ectenes and changes of its catches in Lake Chaohu[J]. Freshwater Fisheries, 2008, 38(6):19-22. |
| [12] | PINKAS L, OLIPHANT M S, IVERSON I L K. Food habits of albacore, bluefin tuna, and bonito in California waters[J]. Fish Bulletin, 1971, 152:1-105. |
| [13] | HYSLOP E J. Stomach contents analysis-a review of methods and their application[J]. Journal of Fish Biology, 1980, 17(4):411-429. |
| [14] | 张觉民,何志辉.内陆水域渔业自然资源调查手册[M].北京:中国农业出版社, 1991.ZHANG J M, HE Z H. Investigation Handbook on Natural Waters Fisheries Resources in the China Inland[M]. Beijing:China Agriculture Press, 1991. |
| [15] | 殷名称.鱼类生态学[M].北京:中国农业出版社, 1995.YIN M C. Fish Ecology[M]. Beijing:China Agriculture Press, 1995. |
| [16] | 林龙山,张寒野,李惠玉,等.东海带鱼食性的季节变化[J].中国海洋大学学报, 2006, 36(6):932-936.LIN L S, ZHANG H Y, LI H Y, et al. Study on seasonal variation of feeding habit of Hairtail (Trichiurus japonicus) in the East China Sea[J]. Journal of Ocean University of China, 2006, 36(6):932-936. |
| [17] | 李宏祥,田华,梁国康,等.淀山湖富营养化现状及生态修复措施分析[J].水资源保护, 2012, 28(3):83-87.LI H Y, TIAN H, LIANG G H, et al. Analysis of eutrophication and measures for ecological restoration in Dianshan Lake[J]. Water Resources Protection, 2012, 28(3):83-87. |
| [18] | CROZIER W W. Observations on the food and feeding of the angler-fish, Lophim piscatorius L., in the northern Irish Sea[J]. Journal of Fish Biology, 1985, 27(5):655-665. |
| [19] | MACDONALD J S, GREEN R H. Redundancy of variables used to describe importance of prey species in fish diets[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1983, 40(5):635-637. |
| [20] | CAILLIET G M. Several approaches to the feeding ecology of fishes[M]//SIMENSTAD C A, LIPOVSKY S J. Fish Food Habits Studies 1st Pacific Northwest Technical Workshop. Washington:Washington Sea Grant, 1997. |
| [21] | ASSIS C A. A generalized index for stomach contents analysis in fish[J]. Scientia Marina, 1996, 60(2/3):385-389. |
| [22] | 周德勇.太湖梅梁湾梅鲚的食性研究:胃含物和稳定同位素分析[D].武汉:华中农业大学, 2011.ZHOU D Y. Feeding habits of lake anchovy (Coilia ectenes) in Meiling bay, Taihu lake:stomach content and stable isotope analysis[D]. Wuhan:Huazhong Agricultural University, 2011. |
| [23] | LIU Z W. Diet of the zooplanktivorous icefish Neosalanx pseudotaihuensis Zhang[J]. Hydrobiologia, 2001, 459(1/3):51-56. |
| [24] | PYKE G H. Optimal foraging theory:a critical review[J]. Annual Review Ecology Systematics, 1984, 15:523-575. |
| [25] | 郑小燕,王丽卿,盖建军,等.淀山湖浮游动物的群落结构及动态[J].动物学杂志, 2009, 44(5):78-85.ZHENG X Y, WANG L Q, GAI J J, et al. Dynamic and community structure of zooplankton in Dianshan Lake[J], Chinese Journal of Zoology, 2009, 44(5):78-85. |
| [26] | 冯德祥,刘一,禹娜,等.淀山湖后生浮游动物群落结构特征分析[J].华东师范大学学报:自然科学版, 2011(6):122-131.FENG D X, LIU Y, YU N, et al. Study on meta-zooplankton community structure of Dianshan Lake[J]. Journal of East China Normal University:Natural Science, 2011(6):126-131. |
| [27] | HALL D J, THRELKELD S T, BURNS C W, et al. The size-efficiency hypothesis and the size structure of zooplankton communities[J]. Annual Review of Ecology and Systematics, 1976, 7(1):177-208. |
| [28] | KORPONAI J, MÁTYÁS K, PAULOVITS G, et al. The Effect of Different Fish Communities on the Cladoceran Plankton Assemblages of the Kis-Balaton Reservoir, Hungary[M]. Cladocera:the Biology of Model Organisms. Springer, 1997:211-221. |
| [29] | XU J, XIE P, ZHANG M, et al. Variation in stable isotope signatures of seston and a zooplanktivorous fish in a eutrophic Chinese lake[J]. Hydrobiologia, 2005, 541(1):215-220. |
| [30] | NORTHCOTE T G. Fish in the structure and function of freshwater ecosystems:A "Top-down" view[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1988, 45(2):361-379. |
2. School of Life Science, East China Normal University, Shanghai 200062, China