 2021, Vol. 40
2021, Vol. 40扩展功能
文章信息
- 田余波, 任艳, 刘海波, 毛羽, 吴发明
- TIAN Yubo, REN Yan, LIU Haibo, MAO Yu, WU Faming
- 甘肃哈思山区芫菁科主要药用昆虫的分布特征
- Distribution Characteristics of the Main Medicinal Insects of Meloidae in the Hasi Mountain, Gansu Province
- 四川动物, 2021, 40(4): 383-393
- Sichuan Journal of Zoology, 2021, 40(4): 383-393
- 10.11984/j.issn.1000-7089.20200032
-
文章历史
- 收稿日期: 2021-01-26
- 接受日期: 2021-05-06






2. 遵义医科大学药学院, 贵州遵义 563003
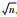 。
。2. Zunyi Medical University, Zunyi, Guizhou Province 563003, China
when m0 is 10 heads per plant. This study lays a foundation for the exploitation of medicinal insects resources of the Meloidae in the Hasi Mountain, and provides a theoretical basis for the decision-making of prevention and control of agricultural hazards caused by the excessive number of Meloidae insects.芫菁科Meloidae昆虫分布广泛,是豆类作物常见害虫(李晓婷,2010;潘昭等,2011),同时该科昆虫多具有重要药用价值,应用历史悠久(刘海波等,2019),如大斑沟芫菁Hycleus phaleratus (Pallas, 1781)和眼斑沟芫菁Hycleus cichorii (Linnaeus, 1758)被《中华人民共和国药典》收录(任艳等,2018;国家药典委员会,2020),其他有相关药用记载还包括绿芫菁Lytta caraganae (Pallas, 1781)、锯角豆芫菁Epicauta gorhami (Marseul, 1873)等(王雪梅,2007;王腾蛟等,2018)。芫菁科昆虫在新疆、甘肃、宁夏、河北、内蒙古等多地广泛分布(张润杨,2018;杨益春,2019),而哈思山区是甘肃芫菁科昆虫聚集的典型代表区域(贺春贵等,2005)。该类昆虫分泌的斑蝥素及其衍生物具有抗肿瘤、增强免疫、杀虫等作用,医疗价值极高(Yan,1993;曾瑶波等,2016;颜道宇等,2017;李济森等,2019)。作为重要的药用昆虫资源,当前市场需求量日益增长,在人工养殖方面存在瓶颈,无法规模化,临床使用仍以野外收集为主,过度不合理采集使其野生数量日渐减少(陈仕江,2004;王雪梅等,2007;莫让瑜,2014),而目前有关芫菁科昆虫分布特征的研究尚无详尽报道。
昆虫种群空间分布是昆虫种群的重要生态学属性之一(杨现明等,2020),不仅对了解昆虫种群的爆发、扩散具有重要的意义,而且在抽样技术及监测、预报中也发挥了一定的指导作用(鲁武锋等,2018)。对芫菁科药用昆虫聚集典型代表区域进行系统调查,探索其空间分布规律和生活习性,对该类药用昆虫资源的保护和综合开发利用具有重要的作用和实际意义。因此,本文采用随机取样法对芫菁科药用昆虫的分布进行调查,利用聚集度指标法、Taylor幂函数法及Iwao回归分析法对昆虫的空间分布类型及抽样技术进行研究,以期为芫菁科药用昆虫资源的监测、保护和综合开发奠定基础,也为防治该科昆虫因数量过度造成农业危害提供参考。
1 材料与方法 1.1 研究地概况哈思山区是以哈思山自然保护区为中心的农林牧交错带,地处甘肃省白银市靖远县境内,属陇中黄土高原农牧林区,地理坐标104°13′40″~104°35′00″E,36°58′20″~37°2′40″N,海拔2 400~3 017 m。年均气温6~7 ℃,年均降水量350~ 410 mm,降水极不均匀,多集中于7—9月(杨万军,2003;张启立等,2019)。全县现有耕地7.58万hm2,草原面积37.33万hm2,其中哈思山区草原主要为高寒草甸类、温性草原类和温性荒漠草原类(董四娟,2012)。该区域植被覆盖较好,但森林植被类型少,同时由于地处旱地农业向荒漠牧地过渡边界,荒漠化较严重,主要有石质荒漠、沙质荒漠和盐化荒漠(舒海元,1997;李连惠,2003)。
1.2 调查方法本研究于2019年7月,当地芫菁科昆虫频繁活动时期,根据哈思山区概况及地形地貌特征,选择农田型、草地型、森林型和荒漠型4种具有代表性的生态类型样地各3块,每块样地设置5 m2(n=7) 正方形样方,样方间距约为500 m。部分采样生境如图 1所示,记录样方内芫菁科主要药用昆虫及其所分布植物的种类和数量。采用实验室自制昆虫捕捉装置进行样方内目标昆虫的捕捉,进行分类学鉴定。

|
| 图 1 哈思山区芫菁科主要药用昆虫采样生境(任艳 摄) Fig. 1 Sampling habitat map of the main medicinal insects of Meloidae in the Hasi Mountain (photoed by REN Yan) |
| |
芫菁科主要药用昆虫空间分布型测定取样方法:选取14个样方(农田型和草地型样地各7个样方),采用随机抽样法取样,每个样方随机选取20株植株,记录植株上芫菁科昆虫数量,并进行空间分布型测定。
1.3 分析方法 1.3.1 空间分布型采用聚集度指标法(王智,2007;贾彦霞等,2019)、Taylor幂函数法(Taylor,1961;杨紫涵等,2020)和Iwao回归分析法(Iwao,1968;刘延超等,2018)对芫菁科主要药用昆虫的空间分布型进行测定。
聚集度指标法:计算得到各样方昆虫的平均虫口密度(m)、样本方差(S2)、平均拥挤度m*。再按照聚集度指标的计算公式分别求出扩散系数(C):C=S2/m、聚块性指数m*/m、负二项分布k值:k=m2/(S2-m)、丛生指数(I):I=(S2-m)/m、聚集度指数(Ca):Ca=1/k=(S2-m)/m2和La指标:La=m-m/S2+1。
Taylor幂函数法:根据Taylor(1961)提出的m与S2的回归关系公式:S2=lga+blgm,式中,a和b分别表示抽样因素和聚集特征。当lga=0、b=1时,种群为随机分布;当lga>0、b=1时,种群在一切密度下呈聚集分布,且不依赖于密度;当lga>0、b>1时,种群在一切密度下呈聚集分布,且依赖于密度聚集;当lga < 0、b < 1时,种群为均匀分布,且密度越高,分布越均匀。
Iwao回归分析法:m*-m回归模型为:m*=α+ βm,m*和m表示平均拥挤度与平均密度。式中,α为分布基本成分的分布性质(平均拥挤程度),当α=0时,分布的基本成分为单个个体;当α>0时,个体间相互吸引,分布的基本成分为个体群;当α < 0时,个体间相互排斥。β为基本成分的空间分布型,当β=1时,种群为随机分布;当β>1时,种群为聚集分布;当β < l时,种群为均匀分布。
1.3.2 聚集原因分析应用Blackith(1958)提出的聚集均数λ对芫菁科主要药用昆虫的聚集因素进行分析。聚集均数公式为:λ=mγ/2k,式中,k为各样方昆虫负二项分布指数,m为平均值,γ为df=2k、P=0.05时的χ2值。当λ≥2时,聚集主要是昆虫的本身习性和环境条件综合影响的结果;当λ < 2时,聚集是由环境因素引起(Blackith,1958;阎雄飞等,2019)。
1.3.3 抽样技术根据Iwao(1968)提出的理论抽样数计算公式:N=t2/D2[(α+1)/m+β-1],计算芫菁科主要药用昆虫在不同虫口密度下的最适抽样数。式中,N为最适抽样数,D为允许误差,取t=1.96(95%可靠度),m为平均虫口密度(张治科等,2018)。
采用Iwao(1968)提出的序贯抽样理论公式: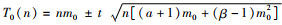
运用SPSS 20.0进行数据统计分析,运用Heat Mapper进行聚类热点图分析。
2 结果与分析 2.1 哈思山区芫菁科主要药用昆虫的分布 2.1.1 芫菁科主要药用昆虫种类及分布情况对所有样地中的芫菁科药用昆虫进行鉴定,常见有西北斑芫菁Mylabris sibirica (Fischer von Waldheim, 1823)、丽斑芫菁M. speciosa (Pallas, 1781)、圆点斑芫菁M. aulica (Ménétriés, 1832)、暗头豆芫菁Epicauta obscurocephala (Reitter, 1873)、扁角豆芫菁E. impressicornis (Pic, 1913)和绿芫菁Lytta caraganae (Pallas, 1781)。调查发现,在不同类型样地中芫菁科主要药用昆虫数量及种类均存在一定差异(表 1)。该区域芫菁科昆虫分布于农田型样地和草地型样地,且农田型样地分布数量远多于草地型样地。西北斑芫菁、丽斑芫菁、圆点斑芫菁、暗头豆芫菁、扁角豆芫菁及绿芫菁在2种类型样地中均有分布,其中,西北斑芫菁在2种样地中最常见、数量最多,圆点斑芫菁次之。森林型和荒漠型样地中均未发现芫菁科昆虫分布。有研究报道,芫菁科昆虫啃食沙棘Hippophae rhamnoides、沙枣Elaeagnus angustifolia、锦鸡儿Caragana microphylla等的叶片(王文和,2015),但本研究的沙棘林及其他森林型样地中均未发现芫菁科昆虫分布。
| 样地类型 Type of plot |
试验点 Test sample |
海拔 Elevation/m |
地理坐标 Geographic coordinate |
昆虫种类 Insect species |
| 农田型 | 万家岘 | 2 169 | 104°35′E, 37°03′N | 西北斑芫菁Mylabris sibirica, 圆点斑芫菁M. aulica, 扁角豆芫菁Epicauta impressicornis, 暗头豆芫菁E. obscurocephala |
| 海家沟 | 2 150 | 104°36′E, 37°01′N | 西北斑芫菁, 圆点斑芫菁, 丽斑芫菁M. speciosa | |
| 雪山寺 | 2 114 | 104°34′E, 37°01′N | 西北斑芫菁, 圆点斑芫菁, 扁角豆芫菁 | |
| 草地型 | 沙河村 | 2 240 | 104°32′E, 37°00′N | 西北斑芫菁, 绿芫菁Lytta caraganae, 丽斑芫菁 |
| 哈思山 | 2 429 | 104°35′E, 37°00′N | 西北斑芫菁, 扁角豆芫菁, 暗头豆芫菁 | |
| 松柏村 | 2 365 | 104°37′E, 37°00′N | 暗头豆芫菁, 圆点斑芫菁 | |
| 荒漠型 | 上红崖湾 | 1 907 | 104°35′E, 37°05′N | — |
| 阎砂河村 | 1 919 | 104°36′E, 37°03′N | — | |
| 九队村 | 1 981 | 104°35′E, 37°05′N | — | |
| 森林型 | 曼仗湾 | 2 134 | 104°32′E, 37°03′N | — |
| 沙河村 | 2 216 | 104°32′E, 37°01′N | — | |
| 四队沟 | 2 208 | 104°34′E, 37°01′N | — |
调查发现, 芫菁科主要药用昆虫分布的植物种类较多(表 2),共10科17种植株,集中于菊科Astera ceae、豆科Leguminosae和唇形科Lamiaceae植物。其中,西北斑芫菁分布的植物种类最丰富,共计11种,是区域内最常见的芫菁类昆虫。根据昆虫在不同植物上的分布情况,对6种芫菁类昆虫进行聚类分析,从分布热点图(图 2)可以看出,芫菁类昆虫可分为2类,西北斑芫菁和圆点斑芫菁为一类,绿芫菁、扁角豆芫菁、丽斑芫菁和暗头豆芫菁为另一类。这是由于西北斑芫菁和圆点斑芫菁对植物的选择相似,2种昆虫多分布在同一样地,均主要取食密花香薷Elsholtzia densa、紫苜蓿Medicago sativa、乳苣Mulgedium tataricum等植物。而其他4种芫菁分布相对零散,其中绿芫菁和扁角豆芫菁分布的植物种类相对较少,绿芫菁主要取食野豌豆Vicia sepium,该地区绿芫菁活动时间集中在豆类植物开花期间,是危害豆类作物生产最主要害虫;扁角豆芫菁主要分布在黄芪Astragalus membranaceus植株上,同时该地区大量种植紫苜蓿作为牧草,其在紫苜蓿地中也分布;丽斑芫菁主要分布在串铃草Phlomis mongolica、冰草Agropyron cristatum等上;暗头豆芫菁分布范围较广,在农田型和草地型样地中均有分布,主要分布在河北石头花Gypsophila tschiliensis和白花枝子花Dracocephalum heterophyllum的植株上,但总体数量较少。
| 植物Plant | 分布昆虫物种 Insect distribution |
|
| 科Family | 种Species | |
| 菊科 Asteraceae |
乳苣 Mulgedium tataricum |
西北斑芫菁Mylabris sibirica, 圆点斑芫菁M. aulica |
| 苦荬菜 Ixeris polycephala |
西北斑芫菁, 圆点斑芫菁 | |
| 中亚紫菀木 Asterothamnus centraliasiaticus |
西北斑芫菁, 扁角豆芫菁Epicauta impressicornis, 暗头豆芫菁E.obscurocephala | |
| 大蓟 Cirsium japonicum |
圆点斑芫菁, 丽斑芫菁M. speciosa | |
| 唇形科 Lamiaceae |
密花香薷 Elsholtzia densa |
西北斑芫菁, 圆点斑芫菁, 暗头豆芫菁 |
| 串铃草 Phlomis mongolica |
西北斑芫菁, 丽斑芫菁, 绿芫菁Lytta caraganae | |
| 白花枝子花 Dracocephalum heterophyllum |
暗头豆芫菁 | |
| 豆科 Leguminosae |
黄芪 Astragalus membranaceus |
西北斑芫菁, 圆点斑芫菁, 扁角豆芫菁, 暗头豆芫菁 |
| 紫苜蓿 Medicago sativa |
西北斑芫菁, 圆点斑芫菁, 扁角豆芫菁 | |
| 野豌豆 Vicia sepium |
绿芫菁 | |
| 十字花科 Cruciferae |
油菜 Brassica campestris |
西北斑芫菁 |
| 蒺藜科 Zygophyllaceae |
骆驼蓬 Peganum harmala |
丽斑芫菁 |
| 禾本科 Gramineae |
冰草 Agropyron cristatum |
丽斑芫菁, 暗头豆芫菁 |
| 茄科 Solanaceae |
马铃薯 Solanum tuberosum |
扁角豆芫菁 |
| 石竹科 Caryophyllaceae |
河北石头花 Gypsophila tschiliensis |
西北斑芫菁, 暗头豆芫菁 |
| 伞形科 Umbelliferae |
碱蛇床 Cnidium salinum |
西北斑芫菁 |
| 百合科 Liliaceae |
薤白 Allium macrostemon |
西北斑芫菁, 圆点斑芫菁 |

|
| 图 2 芫菁科主要药用昆虫在宿主植株上的分布热点 Fig. 2 Hot distribution spots of the main medicinal insects of Meloidae on host plants |
| |
对各样方的调查数据进行聚集度指标计算(表 3),各样方的m*>m、C>1、I>0、Ca>0、m*/m>1、La/m>1、k>0,各项指标表明芫菁科主要药用昆虫的空间分布型为聚集分布。
| 样地 Sample |
m | S2 | m* | C | I | Ca | m*/m | La | k |
| 1 | 1.600 | 2.779 | 2.336 | 1.737 | 0.737 | 0.461 | 1.460 | 2.024 | 2.171 |
| 2 | 2.750 | 5.039 | 3.582 | 1.832 | 0.832 | 0.303 | 1.303 | 3.204 | 3.304 |
| 3 | 1.300 | 2.092 | 1.909 | 1.609 | 0.609 | 0.469 | 1.468 | 1.679 | 2.134 |
| 4 | 2.350 | 4.976 | 3.467 | 2.117 | 1.117 | 0.476 | 1.475 | 2.878 | 2.103 |
| 5 | 2.200 | 4.461 | 3.228 | 2.028 | 1.028 | 0.467 | 1.467 | 2.707 | 2.141 |
| 6 | 2.450 | 4.766 | 3.395 | 1.945 | 0.945 | 0.386 | 1.386 | 2.936 | 2.592 |
| 7 | 0.800 | 1.853 | 2.116 | 2.316 | 1.316 | 1.646 | 2.645 | 1.364 | 0.608 |
| 8 | 0.950 | 1.748 | 1.790 | 1.840 | 0.840 | 0.884 | 1.884 | 1.407 | 1.131 |
| 9 | 1.350 | 2.427 | 2.148 | 1.800 | 0.800 | 0.593 | 1.591 | 1.794 | 1.692 |
| 10 | 1.550 | 2.647 | 2.258 | 1.708 | 0.708 | 0.457 | 1.457 | 1.964 | 2.190 |
| 11 | 0.733 | 1.248 | 1.436 | 1.703 | 0.703 | 0.959 | 1.959 | 1.146 | 1.043 |
| 12 | 1.133 | 1.427 | 1.392 | 1.259 | 0.259 | 0.229 | 1.229 | 1.339 | 4.366 |
| 13 | 2.000 | 2.551 | 2.276 | 1.276 | 0.276 | 0.138 | 1.138 | 2.216 | 7.260 |
| 14 | 1.400 | 1.960 | 1.800 | 1.400 | 0.400 | 0.286 | 1.286 | 1.686 | 3.500 |
将m和S2进行对数转换,分析二者之间线性关系(图 3:a),芫菁科主要药用昆虫Taylor幂函数方程为:lgS2=0.204 2+1.103 8lgm(r=0.886),式中lga=0.204 2>0、b=1.103 8>1,芫菁科主要药用昆虫种群在一切密度下呈聚集分布,且具密度依赖性,聚集强度随种群密度的升高而增加,与应用聚集度指标法测定的结果一致。

|
| 图 3 芫菁科主要药用昆虫lgS2-lgm(a)、m*-m(b)回归分析 Fig. 3 lgS2-lgm (a) and m*-m(b) regression analysis of the main medicinal insects of Meloidae |
| |
将m和m*进行线性回归(图 3:b),m*-m模型回归方程为:m*=0.642 5+ 1.069 7m(r=0.916),由α=0.642 5>0、β=1.0697>1可知,芫菁科主要药用昆虫的空间分布型为聚集分布,种群分布的基本成分是个体群,个体间相互吸引,与采用聚集度指标法和Taylor幂法则分析法测定的种群空间分布型的结果一致。
2.2.4 聚集原因分析根据昆虫的聚集原因分析(表 4),各样方中芫菁科主要药用昆虫的λ>2,表明其聚集原因可能是由其本身的取食、产卵等习性及温度、日照等周围环境条件综合影响或其中之一引起。
| 样地 Sample |
评价指标 Evaluation index |
|||
| m | 2k | γ | λ | |
| 1 | 1.600 | 4.342 | 10.030 | 3.696 |
| 2 | 2.750 | 6.608 | 13.490 | 5.614 |
| 3 | 1.300 | 4.268 | 9.913 | 3.019 |
| 4 | 2.350 | 4.206 | 9.815 | 5.484 |
| 5 | 2.200 | 4.282 | 9.936 | 5.105 |
| 6 | 2.450 | 5.184 | 11.350 | 5.364 |
| 7 | 0.800 | 1.216 | 4.304 | 2.832 |
| 8 | 0.950 | 2.262 | 6.467 | 2.716 |
| 9 | 1.350 | 3.384 | 8.455 | 3.373 |
| 10 | 1.550 | 4.380 | 10.090 | 3.571 |
| 11 | 0.733 | 2.086 | 6.147 | 2.160 |
| 12 | 1.133 | 8.732 | 16.542 | 2.146 |
| 13 | 2.000 | 14.520 | 24.366 | 3.356 |
| 14 | 1.400 | 7.000 | 14.070 | 2.814 |
利用理论抽样数计算公式,取置信水平t=1.96(概率为0.95),芫菁科主要药用昆虫理论抽样模型为:N= (6.310/m+ 0.268)/D2。当D为0.1、0.2和0.3时,m在1、2、4、6、8、10、15……50的条件下,计算得出不同密度条件下的最适抽样数(表 5)。在允许误差一致的情况下,随着平均虫口密度的增加,芫菁科主要药用昆虫的最适抽样数逐渐减少;在平均虫口密度相同的情况下,随着允许误差的增大,芫菁科主要药用昆虫的最适抽样数逐渐减少。
| 允许误差 Permissible error |
平均虫口密度 Average density of Meloidae insects/(头/株) |
|||||||||||||
| 1 | 2 | 4 | 6 | 8 | 10 | 15 | 20 | 25 | 30 | 35 | 40 | 45 | 50 | |
| 0.1 | 658 | 342 | 185 | 132 | 106 | 90 | 69 | 58 | 52 | 48 | 45 | 43 | 41 | 40 |
| 0.2 | 164 | 86 | 63 | 33 | 26 | 22 | 17 | 15 | 13 | 12 | 11 | 11 | 10 | 10 |
| 0.3 | 73 | 38 | 21 | 15 | 12 | 10 | 8 | 6 | 6 | 5 | 5 | 5 | 5 | 4 |
根据序贯抽样理论公式,取置信水平t=1.96(概率为0.95),得到芫菁科主要药用昆虫的序贯抽样模型为:T1, T0(n)=nm0±1.96 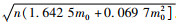
| 防治指标 Control index /(头/株) |
α | β | 序贯抽样模型 Sequential sampling model |
| 2 | 0.642 5 | 1.069 7 | T1, T0(n)=2n±3.700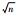 |
| 4 | 0.642 5 | 1.069 7 | T1, T0(n)=4n±5.434 |
| 6 | 0.642 5 | 1.069 7 | T1, T0(n)=6n±6.892 |
| 8 | 0.642 5 | 1.069 7 | T1, T0(n)=8n±8.223 |
| 10 | 0.642 5 | 1.069 7 | T1, T0(n)=10n±9.480 |
| 12 | 0.642 5 | 1.069 7 | T1, T0(n)=12n±10.690 |
| 抽样量 Sampling |
m0=2 | m0=4 | m0=6 | m0=8 | m0=10 | m0=12 | ||||||
| T1 | T0 | T1 | T0 | T1 | T0 | T1 | T0 | T1 | T0 | T1 | T0 | |
| 10 | 32 | 8 | 57 | 23 | 82 | 38 | 106 | 54 | 130 | 70 | 154 | 86 |
| 20 | 57 | 23 | 104 | 56 | 151 | 89 | 197 | 123 | 242 | 158 | 288 | 192 |
| 30 | 80 | 40 | 150 | 90 | 218 | 142 | 285 | 195 | 352 | 248 | 419 | 301 |
| 40 | 103 | 57 | 194 | 126 | 284 | 196 | 372 | 268 | 460 | 340 | 548 | 412 |
| 50 | 126 | 74 | 238 | 162 | 349 | 251 | 458 | 342 | 567 | 433 | 676 | 524 |
| 60 | 149 | 91 | 282 | 198 | 413 | 307 | 544 | 416 | 673 | 627 | 803 | 637 |
| 70 | 171 | 109 | 325 | 235 | 478 | 362 | 629 | 491 | 779 | 621 | 929 | 751 |
| 80 | 193 | 127 | 369 | 271 | 541 | 418 | 714 | 566 | 885 | 715 | 1 056 | 846 |
| 90 | 215 | 145 | 412 | 308 | 605 | 475 | 789 | 642 | 990 | 810 | 1 181 | 979 |
| 100 | 237 | 163 | 454 | 346 | 669 | 531 | 882 | 718 | 1 095 | 905 | 1 307 | 1 093 |
在甘肃哈思山区不同类型样地中,调查发现的芫菁科药用昆虫共3属6种,分布于农田型和草地型样地中,西北斑芫菁、丽斑芫菁、圆点斑芫菁、暗头豆芫菁、扁角豆芫菁及绿芫菁在2种类型样地中均有分布。在调查过程中发现,该区域草地型样地中适宜芫菁科昆虫取食的植物较农田型样地少,多分布在禁牧区和保护区草地,整体分布数量远少于农田型样地。此外,有研究发现芫菁科昆虫与蝗虫、蜜蜂存在着明显的相关性(胡子渊,2020),其幼虫主要取食蝗虫卵或蜜蜂幼虫,而农田中密集分布的作物及其他植物通常吸引了大量的蝗虫和蜜蜂。这可能是造成该区域芫菁科主要药用昆虫在农田型和草地型样地中分布数量差异的原因。
昆虫的空间分布型是由昆虫种群的生物学、生态学特性和所处生境条件所决定的空间分布方式(王小武等,2017;鲁武锋等,2018)。本研究通过聚集度指标法、Taylor幂函数法及Iwao回归法分析哈思山区芫菁科药用昆虫的分布特征,结果均表明芫菁科药用昆虫的空间分布型为聚集分布,分布的基本成分是个体群,与李亚林(2010)研究中描述芫菁科昆虫具有群聚性相符合。芫菁科昆虫为复变态昆虫,雌虫产卵量大,平均产卵达500余粒(刘洋洋,2016)。幼虫为肉食性,主要取食蝗虫卵,部分取食蜂类幼虫。成虫为植食性,喜食植物的花和嫩叶,具有集群危害的特性,调查过程中甚至发现不同种类的芫菁科昆虫会聚集在一起取食,如西北斑芫菁与圆点斑芫菁多分布在同一样地,这可能属于营养群集或“婚期”群集(杨玉霞,2007)。芫菁科药用昆虫的聚集原因究竟是本身的取食、产卵、交配等习性及温度、日照等周围环境条件综合影响,还是其中之一所引起,仍需进一步研究。在空间分布型研究的基础上,本研究对芫菁科昆虫的抽样技术进行分析;在同一允许误差内,随着平均虫口密度的增加,最适抽样数逐渐减少。不同虫口密度下的最适理论抽样数随成虫数量的增大而减小。因而在调查时,通过预估植株上芫菁科药用昆虫的虫口密度,确定抽样的数量,当虫口密度较低时,应适当增加长植株的抽样株数,反之,则可适当减少。同时还确定了芫菁科昆虫的理论抽样模型,建立了不同防治指标下芫菁科昆虫的序贯抽样模型及一定允许误差和不同虫口密度下的序贯抽样结果表。
昆虫是动物药的重要来源之一,是中药的重要组成,占有独特地位。然而,与药用植物相比,有关药用昆虫资源方面的研究相对薄弱(韦桂宁等,2019;张星贤等,2019)。由于其动态的生活史,在时间和空间上的不确定导致研究的不易性,严重制约药用昆虫资源的开发利用。芫菁科药用昆虫种类众多,形成的中药或民间药种类丰富,如中药品种斑蝥、青娘子、葛上亭长、地胆等(黄文华,2001),但是现代药学研究集中在大斑沟芫菁和眼斑沟芫菁分泌的斑蝥素及其衍生物方面(刘亚楠,2013)。一方面可能因过度利用造成2种芫菁野生资源一定程度的破坏,另一方面却使得同类资源大量浪费。因此对于药用昆虫而言,针对其同科同属或者近缘种属大类资源进行系统研究,既能够为医药产业提供丰富的资源,也不易造成资源的过度使用和破坏。
本研究建立了芫菁科主要药用昆虫农田型样地理论抽样模型及序贯抽样模型,分析了该地区芫菁科主要药用昆虫分布特征和生活习性,研究结果为该类药用昆虫资源的监测提供参考,一方面有利于其可持续应用,一方面可为农业生产中该类昆虫数量过度造成的危害提供防治理论依据。
陈仕江, 程地芸, 曾纬, 等. 2004. 斑蝥成虫食物及投食方式探讨[J]. 中国中药杂志, 29(12): 1214, 1218. DOI:10.3321/j.issn:1001-5302.2004.12.035 |
董四娟. 2012. 县级土地利用总体规划实施评价研究[D]. 陕西: 西北农林科技大学.
|
国家药典委员会. 2020. 中华人民共和国药典: 一部[M]. 北京: 中国医药科技出版社.
|
贺春贵, 周军, 吴劲锋, 等. 2005. 甘肃苜蓿田芫菁的种类为害及防治[J]. 草原与草坪, (3): 21-23, 26. DOI:10.3969/j.issn.1009-5500.2005.03.005 |
胡子渊. 2020. 中国青藏高原芫菁科分类[D]. 河北: 河北大学.
|
黄文华. 2001. 芫菁的人工饲养和生殖生态研究[D]. 福州: 福建师范大学.
|
贾彦霞, 王辉, 常春燕, 等. 2019. 柠条重要害虫四线奇尺蛾天津亚种幼虫的空间分布研究[J]. 环境昆虫学报, 41(2): 329-334. |
李济森, 刘石涛, 王平, 等. 2019. 斑蝥素衍生化活性及毒性规律研究进展[J]. 中草药, 50(7): 1734-1738. |
李连惠. 2003. 哈思山油松林群系结构及演替探讨[J]. 林业实用技术, (8): 11-12. |
李晓婷. 2010. 中国芫菁科分类研究(鞘翅目: 多食亚目: 拟步甲总科)[D]. 陕西: 西北农林科技大学.
|
李亚林. 2010. 河北的芫菁资源及扁角豆芫菁的生物学特性与虫体利用[D]. 河北: 河北大学.
|
刘海波, 石晏丞, 任艳, 等. 2019. 斑蝥民族药用考证及其近缘种现代研究概况[J]. 中成药, 41(3): 691-694. |
刘亚楠. 2013. 中药斑蝥研究进展[J]. 中药与临床, 4(4): 50-52. |
刘延超, 高宇, 李颖姣, 等. 2018. 东北春大豆田间黄蓟马空间分布型及抽样技术[J]. 中国油料作物学报, 40(2): 269-274, 301. |
刘洋洋. 2016. 贵州药用芫菁形态学和分子鉴定[D]. 贵阳: 贵州大学.
|
鲁武锋, 兰明先, 夏涛, 等. 2018. 泽兰实蝇幼虫和虫瘿的空间分布型及抽样技术[J]. 南方农业学报, 49(5): 884-890. DOI:10.3969/j.issn.2095-1191.2018.05.09 |
莫让瑜, 孙年喜, 彭锐. 2014. 南方大斑蝥不同地理种群成虫的食物偏嗜性研究[J]. 中国中药杂志, 39(22): 4293-4296. |
潘昭, 任国栋, 李亚林, 等. 2011. 河北省芫菁种类记述(鞘翅目: 芫菁科)[J]. 四川动物, 30(5): 728-730, 733, 845. DOI:10.3969/j.issn.1000-7083.2011.05.011 |
任艳, 石晏丞, 刘海波, 等. 2018. 芫青类中药本源考[J]. 中药材, 41(11): 2703-2708. |
舒海元. 1997. 甘肃省靖远县草地资源及提高草地生产力的途径[J]. 甘肃农业大学学报, (2): 42-46. |
王腾蛟, 赵成坚, 谷颖乐, 等. 2018. HPLC法测定绿芫菁中斑蝥素的含量[J]. 中国药房, 29(7): 930-932. |
王文和. 2015. 玛可河林区绿芫菁对沙棘危害调查与防治对策[J]. 绿色科技, (4): 47-48. DOI:10.3969/j.issn.1674-9944.2015.04.019 |
王小武, 丁新华, 吐尔逊, 等. 2017. 新疆荒漠绿洲稻区稻水象甲幼虫、蛹的空间分布型及抽样技术[J]. 西北农业学报, 26(9): 1385-1394. |
王雪梅, 陈祥盛, 李晓飞. 2007. 芫菁科昆虫的生物学特性及人工养殖研究概况[J]. 贵州农业科学, 35(2): 140-143. DOI:10.3969/j.issn.1001-3601.2007.02.052 |
王雪梅. 2007. 锯角豆芫菁与毛胫豆芫菁的生物学及人工饲养研究[D]. 贵阳: 贵州大学.
|
王智. 2007. 枇杷园蜘蛛混合种群空间分布型的动态分析[J]. 四川动物, 26(1): 101-102. DOI:10.3969/j.issn.1000-7083.2007.01.025 |
韦桂宁, 韦玉茹, 李冬梅. 2019. 药用昆虫药理活性研究进展[J]. 广西科学, 26(5): 477-483. DOI:10.3969/j.issn.1005-9164.2019.05.005 |
阎雄飞, 蒲泰勋, 李刚, 等. 2019. 枣飞象成虫在陕北枣区的空间分布型及抽样技术[J]. 应用昆虫学报, 56(3): 585-594. |
颜道宇, 郑典鹏, 高维鸿, 等. 2017. 去甲斑蝥素诱导肿瘤干细胞凋亡作用研究[J]. 中草药, 48(11): 2237-2241. |
杨万军. 2003. 哈思山林区森林植被水文效应调查研究[J]. 中国水土保持, (12): 24-25. DOI:10.3969/j.issn.1000-0941.2003.12.012 |
杨现明, 孙小旭, 赵胜园, 等. 2020. 小麦田草地贪夜蛾的发生为害、空间分布与抽样技术[J]. 植物保护, 46(1): 10-16, 23. |
杨益春. 2019. 贺兰山鞘翅目昆虫多样性及空间分布格局[D]. 银川: 宁夏大学.
|
杨玉霞. 2007. 中国豆芫菁属Epicauta分类研究(鞘翅目: 拟步甲总科: 芫菁科)[D]. 河北: 河北大学.
|
杨紫涵, 何沐阳, 李建芳, 等. 2020. 草地贪夜蛾幼虫在苗期玉米田的空间分布格局及其抽样技术[J]. 环境昆虫学报, 42(4): 817-828. DOI:10.3969/j.issn.1674-0858.2020.04.4 |
尹炯, 罗志明, 黄应昆, 等. 2018. 甘蔗蓟马在不同甘蔗品种上空间分布型及理论抽样数[J]. 环境昆虫学报, 40(3): 702-707. |
曾瑶波, 刘晓玲, 李创军, 等. 2016. 斑蝥化学成分及体外抗肿瘤活性研究[J]. 中国中药杂志, 41(5): 859-863. |
张启立, 任路明, 韩鼎, 等. 2019. 甘肃省哈思山自然保护区药用植物调查与保护[J]. 中国实验方剂学杂志, 25(24): 131-135. |
张润杨. 2018. 河北省甲虫物种多样性与地理分布格局[D]. 河北: 河北大学.
|
张星贤, 阮洁, 马占强. 2019. 我国药用昆虫资源研究的历史沿革与现状初探[J]. 生物加工过程, 17(6): 615-622. |
张治科, 吴圣勇, 雷仲仁, 等. 2018. 宁夏辣椒花期西花蓟马的空间分布特征研究[J]. 西北农林科技大学学报(自然科学版), 46(3): 142-147. |
Blackith RE. 1958. Nearest-neighbour distance measurements for the estimation of animal populations[J]. Ecology, 39(1): 147-150. DOI:10.2307/1929978 |
Iwao S. 1968. A new regression method for analyzing the aggregation pattern of animal populations[J]. Researches on Population Ecology, 10(1): 1-20. DOI:10.1007/BF02514729 |
Taylor LR. 1961. Aggregation, variance and the mean[J]. Nature, 189(4766): 732-735. DOI:10.1038/189732a0 |
Yan MS. 1993. The preliminary observation on immunosuppressive effect of norcantharidin in mice[J]. Immunopharmacology and Immunotoxicology, 15(1): 79-85. DOI:10.3109/08923979309066935 |