 2021, Vol. 40
2021, Vol. 40扩展功能
文章信息
- 米书慧, 赵唱, 孙嘉, 李宗智, 滕丽微, 刘振生
- MI Shuhui, ZHAO Chang, SUN Jia, LI Zongzhi, TENG Liwei, LIU Zhensheng
- 贺兰山马麝秋季生境选择
- Study on the Habitat Selection of Alpine Musk Deer in the Helan Mountains in Autumn
- 四川动物, 2021, 40(2): 169-175
- Sichuan Journal of Zoology, 2021, 40(2): 169-175
- 10.11984/j.issn.1000-7083.20200423
-
文章历史
- 收稿日期: 2020-11-07
- 接受日期: 2021-01-04






2. 华东理工大学人事处, 上海 200237;
3. 国家林业和草原局保护生物学重点实验室, 哈尔滨 150040
2. Department of Human Resources, East China University of Science and Technology, Shanghai 200237, China;
3. Key Laboratory of Conservation Biology, National Forestry and Grassland Administration, Harbin 150040, China
生境是野生动物个体、种群或群落赖以生存的空间,不仅为动物提供水和食物等资源,还为其提供繁殖、避敌和迁移的庇护场所(Morrison et al., 1992)。动物对生境的选择是对不断变化的环境条件的一种适应行为,是一个非常复杂的过程,发生在不同时空尺度上(Johnson,1980)。影响野生动物生境选择的因素主要包括食物、隐蔽条件、植被类型、气候条件、种间竞争和人为干扰等方面(张明海,李言阔,2005),其影响情况根据地理生态环境与动物生活史阶段的不同可能存在差异。对野生动物生境选择的研究是开展生境质量评价的基础,从而有针对性地制定保护和管理策略,满足动物生活需要,提高其繁殖与存活能力(马建章等,2004)。研究野生动物的生境选择,可以了解在人类社会迅速发展而导致野生动物生境丧失及破碎化不断加剧的背景下野生动物生境适应策略,这对于濒危野生动物保护与管理具有积极意义。
马麝Moschus sifanicus属偶蹄目Artiodactyla麝科Moschidae麝属Moschus,被列为国家Ⅰ级重点保护野生动物,中国特有种,仅分布于青藏高原及邻近省份,贺兰山曾是马麝的主要分布地之一。历史上偷猎盗猎行为以及生境破碎化等导致马麝种群数量骤减以至濒危。对贺兰山西坡马麝生存现状的调查表明,在近些年保护区严格的管护下,马麝的种群数量略有增长,但其分布范围仍较为局限,恢复、壮大贺兰山马麝种群仍然是保护工作的当务之急(杨云天等,2017)。秋季是马麝越冬能量储备关键时期, 当年生亚成体的扩散此时正处于开始期,马麝秋季的生境选择对其种群存活和来年繁殖成功具有重要意义(Meng et al., 2003;孟秀祥等,2011)。对贺兰山马麝秋季的生境选择特征开展研究,可进一步完善贺兰山马麝的生物学资料,揭示影响其不同季节生境选择的重要生态因子,为更好地保护马麝种群资源提供理论支撑。
1 研究区域与研究方法 1.1 研究区域概况贺兰山位于宁夏平原与阿拉善高原之间(105°44′~106°42′E,38°21′~39°22′N),以山脊为界,东坡属于宁夏回族自治区,西坡属于内蒙古自治区。贺兰山属阴山山系,山势呈南北走向,海拔一般为2 000~3 000 m,最高峰鄂博疙瘩位于主分水岭西侧,海拔为3 556.1 m(刘振生等,2007)。贺兰山属第三级地貌,总体呈东仰西倾,东坡山势巍峨险峻,山高沟深,西坡长而平缓,沟谷较小。气候具有典型的大陆性特征,是全国太阳辐射和日照时数最多的地区之一,属于干旱半干旱地区具有代表性的自然综合体和较完整的自然生态系统(庞博,2016)。地跨温带草原与荒漠两大植被区域,涵盖多种生境类型,是阿拉善-鄂尔多斯区域性生物多样性的核心区域,也是西北干旱区非常重要的生物资源宝库,野生动植物资源丰富(梁存柱等,2004;刘秉儒,2010)。
1.2 研究方法2014—2016年秋季(9—10月),结合以往调查中马麝分布位点,在其活动较为频繁的沟段对马麝的生境进行调查。共布设样线22条,基本涵盖了贺兰山的主要植被类型,分别位于哈拉乌头道沟、哈拉乌三道沟火烧坡沟、边渠子沟、大关沟、青树湾火烧坡沟、大青树湾沟、小青树湾沟、冰沟头道沟、冰沟二道沟、冰沟下八段、雪岭子头道沟、雪岭子二道沟,共计12条,每条样线长2.5~5.5 km。由于贺兰山地区马麝种群密度较低,难以观察到实体活动,因此在样线调查过程中,发现马麝活动痕迹(粪便、食痕、足迹、卧迹)时以活动痕迹为中心设置1个10 m×10 m的样方,在该样方四角与中心共设置5个1 m×1 m的小样方,记录其GPS信息及17种生态因子(植被类型、优势乔木、乔木密度、乔木高度、乔木距离、灌木密度、灌木高度、灌木距离、植被盖度、坡向、坡度、坡位、海拔、距水源距离、人为干扰距离、距裸岩距离和隐蔽级),在没有马麝活动痕迹且距利用样方500 m以上的生境中设定对照样方,设定方法与记录信息与利用样方相同。17种生态因子的判断标准如下(刘振生等,2005;滕丽微等,2007):
植被类型:山地疏林草原带、山地混交林带、山地针叶林带、亚高山灌丛和草甸带4种类型。
优势乔木:在所测量的样方中占比>70%的乔木为优势乔木,划分为灰榆Ulmus glaucescens、杜松Juniperus rigida、油松Pinus tabuliformis、山杨Populus davidiana、青海云杉Picea crassifolia和混合型(样方中无任一树种数量达70%)6类。
乔木密度:10 m×10 m样方中所有乔木的数量。
乔木高度:10 m×10 m样方中所有乔木高度的平均值。
乔木距离:10 m×10 m样方的中心点距最近乔木的距离。
灌木密度:10 m×10 m样方中所有灌木的数量。
灌木高度:10 m×10 m样方中所有灌木高度的平均值。
灌木距离:10 m×10 m样方的中心点距最近灌木的距离。
植被盖度:10 m×10 m样方中5个1 m×1 m的小样方内植被覆盖面积的平均值。
坡位:上坡位、中坡位和下坡位。
坡向:阳坡、阴坡和半阴或半阳坡。
坡度:利用65式军用罗盘仪上的坡度计测量。
海拔:利用GPS定位仪记录样方的海拔。
距水源距离:样方中心到水源的直线距离,利用GPS定位仪测算。
人为干扰距离:样方中心到护林点、公路等人为干扰源的直线距离,利用GPS定位仪测算。
距裸岩距离:样方中心到裸岩的直线距离,利用GPS定位仪测算。
隐蔽级:在样方中心树立一个高1 m的木杆,在东、西、南、北4个方位距离中心20 m处测量木杆能见度,即可看见木杆长度占总长度的百分比,然后计算平均值。
1.3 数据处理利用Marcum法(Marcum & Loftsguarden,1980)分析马麝秋季的生境选择,首先使用拟合优度卡方检验分析马麝对非数值型因子是否具有选择性,c2值越大表明两类样方中的生态因子间差异越大。然后用Bonferroni不等式分析马麝对其的选择偏好。Bonferroni置信区间计算公式如下:
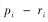
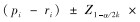
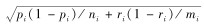
如果0在置信区间内,则有pi=ri,表明马麝对生态因子i为随机选择;如果置信区间的两端都大于0,则有ri显著小于pi,表明马麝避免选择生态因子i;如果置信区间的两端都小于0,则有ri显著大于pi,说明马麝偏好选择生态因子i。
对利用样方和对照样方内的数值型生态因子分别做单个样本的K-S检验,若检验结果不呈正态分布,需要先标准化样本再进行后续分析。在K-S检验结果为显著的基础上,使用Mann-Whitney U检验比较利用样方和对照样方的差异。利用逐步判别分析法对马麝利用样方和对照样方内差异显著的生态因子做进一步的分析,其结果中的Wilk's λ越大,说明该生态因子对马麝生境选择的影响越高,从而得到影响贺兰山马麝秋季生境选择的关键生态因子。
所有数据处理与分析在Excel 2013和SPSS 20.0中完成。
2 结果共记录样方256个(利用样方181个,对照样方75个)。对秋季的4种非数值型生态因子分析的结果表明:马麝对植被类型具有选择性(c2=50.003,df=3,P<0.01),偏好选择山地针叶林带,其他3种植被类型均不选择;对优势乔木具有选择性(c2=37.494,df=5,P<0.01),在优势乔木中选择青海云杉林生境,随机选择混合型生境,避免选择其他4种优势乔木生境;对坡向具有选择性(c2= 32.849,df=2,P<0.01),仅对阴坡有选择,不选择阳坡和半阴半阳坡生境;对坡位具有选择性(c2=16.541,df=2,P<0.01),选择中坡位,避免选择上坡位和下坡位(表 1)。
| 生态因子 Ecological factor |
类别 Category |
实际选择比例 Actual selected proportion (ri) |
期望利用比例 Expected selected proportion (pi) |
pi-ri的置信区间 Boferroni interval for pi-ri |
选择性 Preference |
| 植被类型 | 山地疏林草原带 | 0 | 0.111 | 0.085 7≤pi-ri≤0.136 3 | - |
| 山地混交林带 | 0.067 | 0.167 | 0.072 0≤pi-ri≤0.128 0 | - | |
| 山地针叶林带 | 0.933 | 0.528 | -0.423 8≤pi-ri≤-0.386 2 | + | |
| 亚高山灌丛和草甸带 | 0 | 0.194 | 0.169 9≤pi-ri≤0.218 1 | - | |
| 优势乔木 | 青海云杉 | 0.852 | 0.472 | -0.399 9≤pi-ri≤-0.360 1 | + |
| 杜松 | 0.074 | 0.139 | 0.037 2≤pi-ri≤0.092 8 | - | |
| 油松 | 0.030 | 0.194 | 0.136 7≤pi-ri≤0.191 3 | - | |
| 山杨 | 0 | 0.056 | 0.030 2≤pi-ri≤0.081 8 | - | |
| 灰榆 | 0 | 0.083 | 0.057 7≤pi-ri≤0.108 3 | - | |
| 混合型 | 0.044 | 0.056 | -0.017 0≤pi-ri≤0.041 0 | 0 | |
| 坡向 | 阴坡 | 0.956 | 0.611 | 0.164 1≤pi-ri≤0.201 9 | + |
| 阳坡 | 0.015 | 0.167 | -0.591 6≤pi-ri≤-0.554 4 | - | |
| 半阴坡或半阳坡 | 0.030 | 0.222 | 0.373 7≤pi-ri≤0.406 3 | - | |
| 坡位 | 上坡位 | 0.090 | 0.278 | 0.179 7≤pi-ri≤0.196 3 | - |
| 中坡位 | 0.687 | 0.333 | -0.377 3≤pi-ri≤-0.330 7 | + | |
| 下坡位 | 0.224 | 0.389 | 0.140 7≤pi-ri≤0.189 3 | - | |
| 注:+ 偏好选择,- 避免选择,0随机选择 Notes:+ preference,- avoidance,0 random | |||||
非参数检验Mann-WhitneyU检验对秋季利用样方与对照样方中的13种数值型变量进行比较,结果表明:海拔、乔木密度、乔木高度、乔木距离、灌木密度、人为干扰距离、距裸岩距离、隐蔽级之间存在显著差异,马麝在秋季偏好选择海拔高、乔木密度高、乔木距离近、灌木密度低、人为干扰距离远、裸岩距离远、隐蔽级高的栖息地类型(表 2)。
| 生态因子 Ecological factor |
利用样方 Used quadrats (n=181) |
对照样方 Control quadrats (n=75) |
Z | P |
| 海拔/m | 2 637.61±108.32 | 2 534.98±204.43 | -2.41 | < 0.05 |
| 乔木密度/(棵/100 m2) | 83.25±18.09 | 39.25±14.32 | -8.64 | < 0.01 |
| 乔木高度/m | 10.20±1.63 | 12.78±2.92 | 5.11 | < 0.01 |
| 乔木距离/m | 0.75±0.45 | 1.92±1.19 | 5.42 | < 0.01 |
| 灌木密度/(丛/100 m2) | 2.81±4.93 | 8.42±7.70 | 4.04 | < 0.01 |
| 灌木高度/m | 0.54±0.62 | 0.61±0.49 | 0.94 | >0.05 |
| 灌木距离/m | 0.94±1.28 | 0.33±0.51 | -0.66 | >0.05 |
| 植被盖度/% | 75.85±13.39 | 62.53±27.95 | -1.72 | >0.05 |
| 坡度/° | 28.96±7.61 | 23.14±15.71 | -1.77 | >0.05 |
| 水源距离/m | 1 838.44±1 443.57 | 1 589.67±734.81 | -0.01 | >0.05 |
| 人为干扰距离/m | 1 730.82±450.76 | 1 097.89±275.39 | 6.66 | < 0.01 |
| 距裸岩距离/m | 605.52±220.74 | 423.36±333.84 | -3.97 | < 0.01 |
| 隐蔽级/% | 52.24±16.89 | 50.83±16.10 | 2.90 | < 0.01 |
对贺兰山马麝秋季利用样方与对照样方中差异性显著的各生态因子进行逐步判别分析,结果表明:在影响马麝秋季生境选择的生态因子中,用于区分利用样方和对照样方的生态因子按照贡献值的大小依次为:隐蔽级、距裸岩距离、海拔、人为干扰距离、灌木密度、乔木高度、乔木距离和乔木密度(表 3)。利用这8种生态因子区分利用样方和对照样方时,正确率达到88.5%。
| 生态因子 Ecological factor |
判别系数 Coefficients |
Wilk's λ | F | P |
| 隐蔽级 | 0.429 | 0.957 | 7.507 | < 0.01 |
| 距裸岩距离 | -0.230 | 0.917 | 15.280 | < 0.01 |
| 海拔高度 | -0.282 | 0.910 | 16.672 | < 0.05 |
| 人为干扰距离 | 0.506 | 0.886 | 21.659 | < 0.01 |
| 灌木密度 | 0.366 | 0.856 | 28.334 | < 0.01 |
| 乔木高度 | 0.327 | 0.776 | 48.821 | < 0.01 |
| 乔木距离 | 0.457 | 0.663 | 85.823 | < 0.01 |
| 乔木密度 | -0.554 | 0.481 | 182.211 | < 0.01 |
栖息地管理是野生动物管理和保护的核心,生境选择研究是开展栖息地管理的前提和基础(Morris et al., 2008;Khadka & James,2016)。动物对环表 2贺兰山马麝秋季生境利用样方和对照样方中13种生态因子的比较境中不同生态因子的选择不同,同一物种在不同季节对生境的选择也存在差异,这是一种具有物种和地域特征的进化稳定对策,有利于动物的生存和发展(张冬冬等,2015)。本研究对马麝秋季生境利用样方和对照样方的生态因子进行分析,发现马麝在隐蔽级、距裸岩距离、海拔、人为干扰距离、灌木密度、乔木高度、乔木密度、植被类型、优势乔木、坡向和坡位等方面的选择都具有显著差异。
马麝秋季生活在贺兰山高海拔地区以青海云杉为优势乔木的山地针叶林中,而在孟秀祥等(2011)对甘肃兴隆山马麝秋季生境选择的研究中,马麝更偏好针阔混交林、阔叶林及灌丛等包含灌木成分的植被类型,这可能与秋季贺兰山地区人为活动频繁有关。秋季贺兰山阴坡盛产蘑菇,由于上山偷采蘑菇的人屡禁不止,成为贺兰山秋季最主要的人为干扰。马麝生性胆小,可能为了躲避人为干扰而选择在海拔较高的针叶林地带活动觅食。高大的青海云杉林能够为马麝提供较好的隐蔽条件,保护种群的安全,而且山地针叶林带开阔区域的草本、苔藓较多,食物资源较好,可以满足马麝为越冬及交配储备能量的需求。另外,麝类动物生性机警、行动敏捷,主要避敌方式为迅速跳跃逃跑,并不完全依赖隐蔽物,在针对其他地区麝类的栖息地研究中已有类似的发现(吴建平等,2007;孟秀祥等,2010;张冬冬等,2015;张冬冬, 张秋霞,2018),过于密集的灌丛可能影响马麝活动的便捷度并阻止其快速逃跑,因此马麝倾向于选择灌木密度相对较低的针叶林生境。
食物对动物的生境选择具有显著影响(赵宠南等,2012;穆文靖,2016;Nelson & Gillam,2017)。马麝属于典型的精饲者,以植物嫩叶细枝为食(徐嘉等,2018)。秋季由于气温下降,高海拔区灌木和草本植物逐渐枯萎,这个时期的马麝主要采食位于海拔2 700~3 100 m且多分布于阴坡的蓝锭果忍冬Lonicera edulis、密齿柳Salix characta等灌木以及秦氏黄芪Astragalus chingianus、折枝绣线菊Spiraea tomentulosa、内蒙野丁香Leptodermis ordosica等(赵唱, 2017),因此马麝对阴坡的选择性更强。在坡位选择上,马麝倾向于选择中坡位,一方面可能是中坡位的食物丰富度较上坡位高,且中坡位较高的乔木密度更利于马麝隐蔽;另一方面是中坡位海拔较高,人为干扰相对较小,是较适宜的栖息生境。秋季的贺兰山气温日较差大,冷暖气流交汇,降雨量增多,可以保障马麝的饮水需求,因此对水源距离的选择差异不显著。
隐蔽条件是影响动物生境选择的重要环境因子之一。本研究表明,隐蔽级是影响马麝秋季生境选择贡献值最大的生态因子。马麝性情胆怯,需要隐蔽度较高、距离干扰源远的生境以躲避敌害(吴建平等,2006)。夏季是马麝的哺乳期,而秋季马麝的幼崽正处于生长期,马麝还需要为越冬及交配储备能量,因此需要更好的隐蔽条件来保证幼崽的存活率和自身能量的摄取,同时也为躲避天敌节省能量。在2015年对贺兰山西坡马麝现状的研究中发现,过去猖獗的盗猎致使贺兰山地区马麝的生活习性发生了较大变化,栖息地海拔明显增高且更加隐蔽,平时活动更为机警(杨云天等,2017)。而秋季贺兰山气候宜人、物产丰富,是人类活动较为频繁的时期,马麝更需要选择隐蔽级更高的高海拔、高乔木密度的生境以躲避人们来往游览、采蘑菇、挖药材甚至偷猎的干扰。在对贺兰山马麝的保护管理中,应该注意加强管控力度,禁止居民进入保护区采摘野菜等违规行为,杜绝偷猎盗猎行为,尽可能将人为干扰降至最低,为马麝种群生存和发展的适宜栖息地提供保障。
梁存柱, 朱宗元, 王炜, 等. 2004. 贺兰山植物群落类型多样性及其空间分异[J]. 植物生态学报, 28(3): 361-368. DOI:10.3321/j.issn:1005-264X.2004.03.011 |
刘秉儒. 2010. 贺兰山东坡典型植物群落土壤微生物量碳氮沿海拔梯度的变化特征[J]. 生态环境学报, 19(4): 883-888. DOI:10.3969/j.issn.1674-5906.2010.04.025 |
刘振生, 王小明, 李志刚, 等. 2005. 贺兰山岩羊冬春季取食生境的比较[J]. 动物学研究, 26(6): 580-589. DOI:10.3321/j.issn:0254-5853.2005.06.002 |
刘振生, 王小明, 李志刚, 等. 2007. 贺兰山岩羊的数量与分布[J]. 动物学杂志, 42(3): 1-8. DOI:10.3969/j.issn.0250-3263.2007.03.001 |
马建章, 邹红菲, 贾竞波. 2004. 野生动物管理学[M]. .
|
孟秀祥, 潘世秀, 惠岑怿, 等. 2011. 甘肃兴隆山自然保护区濒危马麝(Moschus sifanicus)秋季利用生境特征[J]. 应用与环境生物学报, 17(3): 412-417. |
孟秀祥, 潘世秀, 栾晓峰, 等. 2010. 兴隆山自然保护区马麝春季生境选择[J]. 生态学报, 30(20): 5509-5517. |
穆文靖. 2016. 食物对原麝采食生境选择的影响研究[D]. 哈尔滨: 东北林业大学.
|
庞博. 2016. 贺兰山蓝马鸡繁殖期觅食地和巢址的微生境选择与利用[D]. 哈尔滨: 东北林业大学.
|
滕丽微, 刘振生, 宋延龄, 等. 2007. 海南大田自然保护区野猪的生境选择[J]. 动物学杂志, 42(1): 1-7. |
吴建平, 张海龙, 张勇. 2007. 大兴安岭原麝冬季的生境选择[J]. 动物学杂志, 42(4): 45-50. |
吴建平, 周玲玲, 穆立蔷. 2006. 小兴安岭通河林区原麝夏季对生境的选择[J]. 兽类学报, 26(1): 44-48. |
徐嘉, 暴旭, 刘振生, 等. 2018. 贺兰山同域分布高山麝和阿拉善马鹿秋季食性的比较研究[J]. 生态学报, 38(10): 3705-3711. |
杨云天, 杨玉萍, 娜荷芽, 等. 2017. 贺兰山西坡马麝生存现状研究[J]. 农业与技术, 37(8): 31. |
张冬冬, 张秋霞. 2018. 九龙山自然保护区林麝(Moschus berezovskii)的冬季生境选择[J]. 四川林业科技, 39(4): 64-68. |
张冬冬, 朱洪强, 葛志勇, 等. 2015. 黄泥河自然保护区原麝冬季栖息地的选择[J]. 西北农林科技大学学报(自然科学版), 43(6): 15-20. |
张明海, 李言阔. 2005. 动物生境选择研究中的时空尺度[J]. 兽类学报, 25(4): 395-401. |
赵唱. 2017. 贺兰山马麝的营养采食策略和微生境选择及评价研究[D]. 哈尔滨: 东北林业大学.
|
赵宠南, 苏云, 刘振生, 等. 2012. 贺兰山牦牛冬春季的生境选择[J]. 生态学报, 32(6): 1762-1772. |
Johnson DH. 1980. The comparison of usage and availability measurements for evaluating resource preference[J]. Ecology, 61(1): 65-71. |
Khadka KK, James DA. 2016. Habitat selection by endangered Himalayan musk deer (Moschus chrysogaster) and impacts of livestock grazing in Nepal Himalaya: implications for conservation[J]. Journal for Nature Conservation, 31: 38-42. |
Marcum CL, Loftsguarden DO. 1980. A nonmapping technique for studying habitat preferences[J]. Journal of Wildlife Management, 44(3): 963-968. |
Meng XX, Yang QS, Feng ZJ, et al. 2003. Timing and synchrony of parturition in Alpine musk deer (Moschus sifanicus)[J]. Folia Zoologica, 52(1): 39-50. |
Morris DW, Clark RG, Boyce MS. 2008. Habitat and habitat selection: theory, tests, and implications[J]. Israel Journal of Ecology and Evolution, 54(3-4): 287-294. |
Morrison ML, Marcot BG, Mannan RW. 1992. Wildlife habitat relationship: concepts and application[M]. .
|
Nelson JJ, Gillam EH. 2017. Selection of foraging habitat by female little brown bats (Myotis lucifugus)[J]. Journal of Mammalogy, 98(1): 223-231. |