 2019, Vol. 38
2019, Vol. 38扩展功能
文章信息
- 罗欢, 肖雪, 李玉杰, 侯金, 董鑫, 张晋东, 白文科, 李程
- LUO Huan, XIAO Xue, LI Yujie, HOU Jin, DONG Xin, ZHANG Jindong, BAI Wenke, LI Cheng
- 利用红外相机建立川金丝猴的行为谱及PAE编码系统
- Establishment of PAE Coding System and Ethogram of Rhinopithecus roxellana Using Infrared Camera
- 四川动物, 2019, 38(6): 646-656
- Sichuan Journal of Zoology, 2019, 38(6): 646-656
- 10.11984/j.issn.1000-7083.20180394
-
文章历史
- 收稿日期: 2018-12-18
- 接受日期: 2019-07-30






2. 西华师范大学生态研究院, 四川南充 637002
2. Institute of Ecology, China West Normal University, Nanchong, Sichuan Province 637002, China
外界环境和内在需求决定了动物行为的形成,而复杂多变的自然环境则是行为多样性形成的主要驱动力(严康慧等,2006)。通过建立行为谱对川金丝猴Rhinopithecus roxellana的各种行为进行定义和分类,将行为发生的环境、具体姿势、动作相对应,有助于我们理解其行为功能、行为与环境间的相互关系,为川金丝猴行为生态学的深入研究奠定基础(蒋志刚,2000)。
在野外动物行为的研究工作中,常用的观察方法有随机取样法、目标取样法和扫描取样法。其中,随机取样法是对动物行为习性的广泛观察和记录,目标取样法则是对动物某一特定活动的详细观察和记录,而扫描取样法是对动物发生的瞬时状态的观察和记录。以上观察方法具有直观性、真实性,但工作量大,且在相当一段时间内不能遇到目标动物,具有偶然性。野生川金丝猴活动范围广,对人类存在着警惕心理,因此很难直接观察其行为动作;且观察者距离被观察动物太近,会对其行为造成干扰,导致观察结果偏差。红外相机监测作为一种零损伤性的、可以24 h不间断持续工作的野外调查方法,已成为珍稀野生动物监测与行为观察研究中一种重要的研究手段(李晟等,2014;张晋东等,2015),且红外相机所捕获的视频和照片数据能真实地反映野外动物的行为、环境特征。因此,基于环境、行为的野生动物行为谱的建立和行为编码系统的编写可借助红外相机完成(王盼等,2018;张晋东等,2018)。
动物行为谱的建立和PAE编码系统的编写首见于蒋志刚(2000),作者分辨记录了麋鹿Elaphurus davidianus的12种姿势、92种动作和134种行为,且该方法得到了广泛认可。随后便逐渐有学者将此方法应用于灵长类的行为研究,如李勇等(2013)分辨并记录了滇金丝猴R. bieti的14种姿势、82种动作和143种行为,崔多英等(2014)分辨和记录了黔金丝猴R. brelichi的16种姿势、107种动作和163种行为等,这些研究为灵长类的行为学研究奠定了基础。本研究尝试利用红外相机对野生川金丝猴行为的监测数据建立川金丝猴的行为谱和行为编码系统,以此丰富灵长类行为研究的基础数据。
川金丝猴隶属于灵长目Primates疣猴亚科Colobinae金丝猴属Rhinopithecus,是中国特有的灵长类动物,国家Ⅰ级重点保护野生动物(蔚培龙,2011)。野生川金丝猴长年栖息于海拔1 500~3 300 m的森林中,主要分布在四川、湖北、甘肃、陕西等省的部分山区(汪松,1998)。目前的相关研究多基于圈养川金丝猴的行为或野生川金丝猴的特定行为,如余建秋等(2015)对圈舍丰容前后的川金丝猴行为进行观察,比较了各种行为发生的频次变化;依据取食优势判断川金丝猴群一雄多雌单元(OMU)的等级地位,根据优势资源占有情况判定雌性川金丝猴的等级地位等(王晓卫等,2007;赵海涛等,2013)。在这些行为生态学研究中,已发现诸多行为动作模式,如哺乳行为(杨晓军,1996)、交配行为(任宝平等,2002)、冲突行为(蔚培龙等,2009)、摄食行为和繁殖行为(Li,2010)、相互理毛行为(禹洋,2012)等,但尚未有针对野生川金丝猴行为进行系统编码的研究报道。为更好地进行川金丝猴野外行为研究,本研究整理分析红外相机监测所得的视频和照片数据,对野生川金丝猴的各种行为进行系统编码,构建行为谱,为深入研究野生川金丝猴行为积累基础数据。
1 研究区概况与研究方法 1.1 研究区域研究地点位于四川省汶川县卧龙国家级自然保护区(102°52′~103°24′E,30°45′~31°25′N),保护区是以保护大熊猫Ailuropoda melanoleuca、川金丝猴、小熊猫Ailurus fulgens等珍稀动物为主的自然保护区,东西长52 km,南北宽62 km,总面积约2 000 km2。保护区动植物种类繁多,植物种类数近4 000种,动物有2 200种左右,其中兽类103种,鸟类281种,昆虫1 700多种(谭迎春等,1995)。
研究区域为野生川金丝猴偏好的栖息环境——五一棚、核桃坪、牛头山和天台山(铁军等,2009)。五一棚观察站主要位于臭水沟流域,面积35 km2,海拔2 300~3 400 m,分布有针叶混交林和亚高山针叶林(胡锦矗,1990);核桃坪位于五一棚的东北方向,包含研究中心、黑岩窝和三岔沟,海拔1 840~3 200 m,主要分布有针阔混交林、亚高山针叶林、落叶阔叶林、常绿落叶阔叶混交林等(张晋东等,2015);牛头山位于保护区东南部,海拔1 900~3 186 m,主要分布有针阔混交林、落叶阔叶林、针叶林和灌丛、常绿落叶阔叶混交林;天台山位于保护区的东北方向,主要分布有落叶阔叶林和拐棍竹Fargesia robusta Yi林。
1.2 红外相机的安装及数据的预处理2012年1月—2016年6月,在研究区域内选择川金丝猴经常活动的地点(通过观察川金丝猴的粪便、足迹等判断)安装红外相机(Ltl Acorn ltl5210 ATM,深圳),其中五一棚15台、核桃坪83台、牛头山4台、天台山18台,共计120台(图 1)。首先将相机固定在离地面50~80 cm高的树干或其他固定物上,并适当调整角度以获取一定高度的数据,各相机间距约为200 m。在放置固定过程中,要求相机取景合适,牢固稳定。红外相机设置为2次连拍,拍摄间隔为2 s,在拍摄照片之后,相机自动切换为录像模式,时长为20 s。相机安置好后,测量GPS位点、坡度、坡向、海拔、生境等信息,并记录放置的时间、日期及相机编号。每2个月采集一次数据,同时更换电池。将拍摄到的照片与视频数据下载到电脑,按照相机编号,辨识并选出川金丝猴的照片与视频数据,分别存到文件夹中(苏彦捷等,1990;张晋东等,2015)。

|
| 图 1 卧龙国家级自然保护区红外相机布设位点 Fig. 1 Location of infrared camera traps in the Wolong National Nature Reserve |
| |
参照蒋志刚(2000)对麋鹿行为的编码方法,定义川金丝猴的行为、姿势、动作及环境。
行为:指川金丝猴在特定的环境条件下,采取一定的姿势来完成满足自身需求的一系列动作。
姿势:指川金丝猴在完成某动作时,身体的主要结构部分所处的位置和呈现的形状。
动作:指川金丝猴为完成某一目的,在较短时间内所发生的肌群运动,从而引发的机体部分结构运动、舒张、弯曲和位移。
环境:指川金丝猴完成动作时所依赖的除自身需求以外的条件,包括生物环境和非生物环境。
姿势、动作及环境是行为的3个要素。动物行为是以一定姿势为基础,一定环境为条件完成的一系列动作,对动物的生存具有重要意义。姿势、动作与环境的不同组合,产生了各种各样的动物行为。对行为姿势、动作、环境进行P码、A码和E码的编排,从而形成相对应的PAE编码系统(蒋志刚,2000;任仁眉等,2000;严康慧等,2006)。
2 结果与分析 2.1 川金丝猴的姿势编码结合李勇等(2013)对滇金丝猴以及崔多英等(2014)对黔金丝猴各种姿势的定义,本研究通过对红外相机数据的姿势辨别分析,发现川金丝猴有站、坐、走、立、躺、跑、跳、趴、抱、悬挂、臂摆荡、蜷缩、攀爬、携挂、哺乳、爬跨、匍匐17种姿势(表 1)。其中,前13种是两性共有的姿势,爬跨是雄性的常见姿势,携挂和哺乳多出现在成年雌性和婴幼猴之间。匍匐是川金丝猴的新姿势类型,与黔金丝猴和滇金丝猴不同。
| 编码Code | 姿势Posture | 定义Definition |
| 1 | 站 | 四肢直立承重 |
| 2 | 立 | 后肢直立承重,前肢离开支撑物 |
| 3 | 坐 | 两后肢向前伸展或弯曲,臀部接触地面、树枝等支撑物,前肢离开支撑物 |
| 4 | 走 | 前后肢或两后肢左右交错接触地面、树枝等支撑物,身体向前移动 |
| 5 | 跑 | 前后肢或两后肢快速左右交错接触地面、树枝等支撑物,身体快速向前移动 |
| 6 | 跳 | 两前肢或两后肢同时撑地/树枝使身体快速腾空,同时落地/落在树枝,身体向前或向两侧移动 |
| 7 | 趴 | 四肢和腹部同时接触地面、树枝等支撑物 |
| 8 | 抱 | 某一个体利用前肢将其他个体揽在怀中 |
| 9 | 躺 | 四肢伸展,身体一侧接触地面、树枝等支撑物 |
| 10 | 蜷缩 | 屈服者采取坐姿,低头,身体前倾,前肢放于膝盖,垂尾 |
| 11 | 悬挂 | 四肢或后肢固定于树枝等支撑物,身体下垂 |
| 12 | 臂摆荡 | 两前肢交替握住上部树枝等支撑物,两后肢悬空,向前移动 |
| 13 | 攀爬 | 前后肢左右交错接触树干、岩壁等支撑物,身体向上移动一定距离 |
| 14 | 爬跨 | 雄性身体前倾,两后肢踩在雌性的两后肢上,趴在雌性背部,两前肢抓住雌性的背部,头向下,尾巴自然下垂;同时,雌性与其配合,把臀部向上翘 |
| 15 | 携挂 | 携带者将婴幼猴放于腹部,婴幼猴用四肢抓住携带者的腹部,携带者通常用前肢揽着被携带者 |
| 16 | 哺乳 | 成年雌性采用站姿或坐姿给婴幼猴哺乳 |
| 17 | 匍匐 | 雌性向雄性邀请与之交配的行为模式,其面部和腹部紧贴地面或树枝等支撑物,前肢和后肢成弯曲状靠近腹部,尾巴自然下垂 |
结合川金丝猴的躯体结构划分:头颈部、眼耳鼻部、嘴部、四肢、腰荐腹部,共辨识发现川金丝猴的84种动作,并对动作进行系统编码(附表 1)。
| 动作Act | 编码Code | 动作Act | 编码Code |
| 头颈部 | 摘 | 42 | |
| 抬头 | 1 | 挖 | 43 |
| 低头 | 2 | 推 | 44 |
| 左转 | 3 | 摇 | 45 |
| 右转 | 4 | 理毛 | 46 |
| 后转 | 5 | 踩 | 47 |
| 前倾 | 6 | 四肢站立 | 48 |
| 摇头 | 7 | 三肢站立 | 49 |
| 回头 | 8 | 后肢站立 | 50 |
| 嘴部 | 前行 | 51 | |
| 啃 | 9 | 后退 | 52 |
| 嚼 | 10 | 侧迈步 | 53 |
| 咬 | 11 | 跑 | 54 |
| 舔 | 12 | 跳 | 55 |
| 扯 | 13 | 攀爬 | 56 |
| 衔 | 14 | 荡 | 57 |
| 吐 | 15 | 前肢击地 | 58 |
| 吻 | 16 | 前肢弯曲 | 59 |
| 张嘴 | 17 | 单前肢弯曲 | 60 |
| 闭嘴 | 18 | 前肢伸直 | 61 |
| 露齿 | 19 | 单前肢伸直 | 62 |
| 吞咽 | 20 | 后肢弯曲 | 63 |
| 吮吸 | 21 | 单后肢弯曲 | 64 |
| 喝 | 22 | 后肢伸直 | 65 |
| 哈欠 | 23 | 单后肢伸直 | 66 |
| 眼耳鼻部 | 腰荐腹部 | ||
| 闭眼 | 24 | 平腰 | 67 |
| 眨眼 | 25 | 伸腰 | 68 |
| 瞪眼 | 26 | 弯腰 | 69 |
| 注视 | 27 | 塌腰 | 70 |
| 眺望 | 28 | 左转身 | 71 |
| 嗅 | 29 | 右转身 | 72 |
| 触 | 30 | 抬尾 | 73 |
| 四肢 | 翘尾 | 74 | |
| 拍 | 31 | 垂尾 | 75 |
| 搓 | 32 | 摆尾 | 76 |
| 按 | 33 | 爬跨 | 77 |
| 握 | 34 | 匍匐 | 78 |
| 抓 | 35 | 勃起 | 79 |
| 捏 | 36 | 插入 | 80 |
| 揽 | 37 | 抽动 | 81 |
| 抱 | 38 | 射精 | 82 |
| 拉 | 39 | 停顿 | 83 |
| 扒 | 40 | 退下 | 84 |
| 掀 | 41 |
红外相机数据显示,川金丝猴行为发生的环境共23种,包含生物环境与非生物环境(表 2)。
| 环境 Environment |
生物环境 Biotic (E1) |
非生物环境 Abiotic (E2) |
编码 Code |
| 乔木 | √ | 1 | |
| 灌木 | √ | 2 | |
| 草丛 | √ | 3 | |
| 竹林 | √ | 4 | |
| 岩石 | √ | 5 | |
| 雪地 | √ | 6 | |
| 裸地 | √ | 7 | |
| 河沟 | √ | 8 | |
| 水塘 | √ | 9 | |
| 山脊 | √ | 10 | |
| 山坡 | √ | 11 | |
| 山谷 | √ | 12 | |
| 阳坡 | √ | 13 | |
| 阴坡 | √ | 14 | |
| 半阳坡 | √ | 15 | |
| 雄性 | √ | 16 | |
| 雌性 | √ | 17 | |
| 婴幼猴 | √ | 18 | |
| 少年猴 | √ | 19 | |
| 母子群 | √ | 20 | |
| 全雄群 | √ | 21 | |
| 家庭群 | √ | 22 | |
| 聚合群 | √ | 23 | |
| 单一个体 | √ | 24 | |
| 孤猴(老年雄性) | √ | 25 |
通过辨别红外相机数据,共观察到野外川金丝猴的116种行为。其中,跳跃采食是新发现的1种野外川金丝猴的摄食行为,即川金丝猴通过后肢发力跃身离开地面(树干等支撑物)以获取更高处优质食物的行为,该行为在其他2种金丝猴的行为谱中还未见报道。借鉴李勇等(2013)对滇金丝猴行为和崔多英等(2014)对黔金丝猴行为的定义,将川金丝猴的行为归属于摄食、调温、交配、母性和幼儿、亲密、作威、威胁、攻击、屈服、聚群、休息、运动、其他等13大类行为(附表 2)。
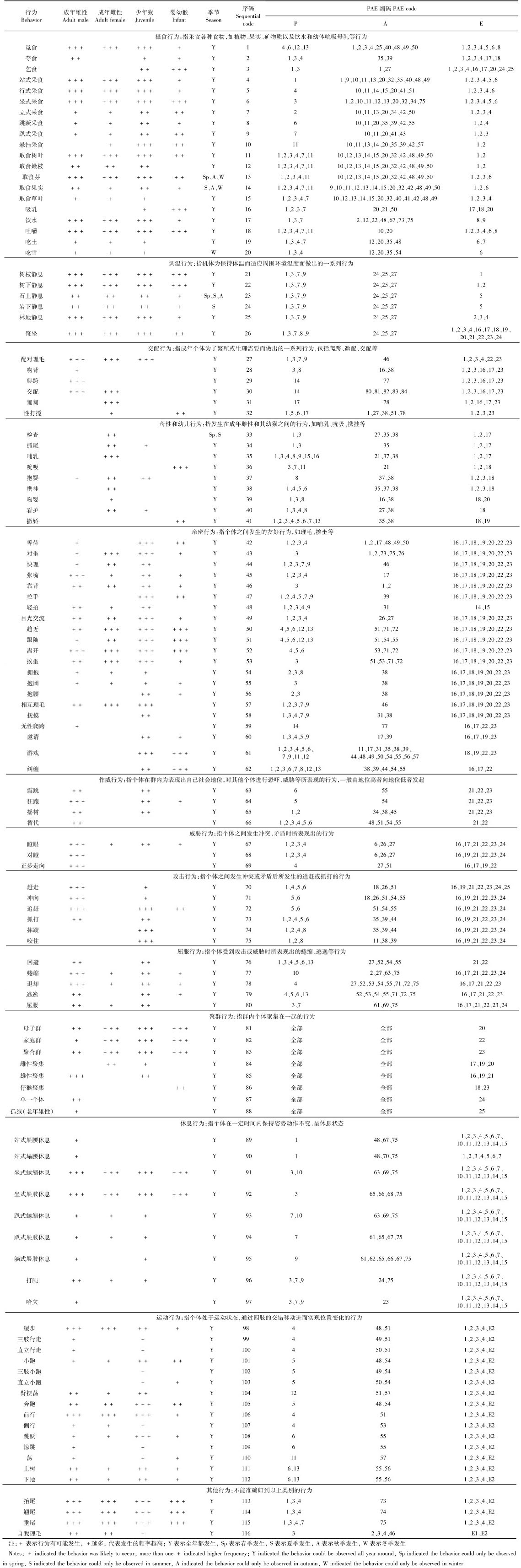
经本研究辨识发现川金丝猴共有17种姿势、84种动作和116种行为,并为其建立了行为谱和PAE编码系统,旨在为川金丝猴的野外行为学研究奠定基础。行为的发生是以姿势、动作的组合表达而实现的,金丝猴灵活的四肢赋予其多样的姿势、丰富的动作,加上多变的自然环境,这些促使了金丝猴行为多样性的形成。川金丝猴、滇金丝猴和黔金丝猴具有较近的亲缘关系,因此在行为模式上存在一定的相似性,但是各自栖息环境的不同以及长时间的地理隔离使得它们在相似的行为模式上表现出了各自的特殊性。这种特殊性主要体现在以下2个方面:首先,特殊的环境条件产生了特殊的行为,如野生川金丝猴为获得较高处的食物会选择跳跃采食,该行为较多发生在少年猴中,这可能与其利用食物资源的方式、取食习惯和体型有关;在川金丝猴的发情期,雌性会采取匍匐的姿势来邀请雄性进行交配,以完成繁殖。经统计,该行为在混合群中发生的频率较高,且通常发生在乔木、灌丛类生境中,但其中的内在驱动因素尚不清楚,还需进一步研究(Ren et al., 1995;严康慧等,2006)。以上2种行为在滇金丝猴和黔金丝猴的行为谱中尚未出现(李勇等,2013;崔多英等,2014)。其次,相同的动作模式也因为长时间的地理隔离和后天的学习而产生了不同的行为效应,例如同样是“快理”这一动作,在川金丝猴中表达了争斗后的和解,而在滇金丝猴中传达的却是母猴对婴幼猴受到惊吓后的抚慰。李勇等(2013)和崔多英等(2014)分别详细阐述了“瞪眼”“快理”“抱团”“抱腰”“支持”“张嘴”等动作在不同种金丝猴中所产生的不同行为效应。由此可见,同属的川金丝猴、滇金丝猴和黔金丝猴由于各自生存环境的差异、长时间的地理隔离和经验传递,导致了各自行为模式的差异(Nishida,1987;Whiten et al., 2001)。
川金丝猴是典型的树栖物种,红外相机虽不能全面监测其树上活动;但红外相机放置的合适角度、较大的视野范围,以及川金丝猴大部分时间在地面活动的习性,使其地面的各种行为及树上的大部分行为被捕获,因此利用红外相机完成野生川金丝猴行为谱的建立是切实可行的。用红外相机进行野外监测,没有观察者的干扰,能够更真实地记录野生动物在自然情况下的行为特征,且大大缩短了观察者的工作时间。另外,红外相机所得的数据可长期保存、能重复查阅,与圈养或野外直接观察所得的数据不同,能真实而生动地描述当时所出现的情景,对野生动物的行为观察方面有促进作用。但与圈养观察和野外直接观察相比,红外相机监测不能获得一些行为动作,如分娩行为、通讯行为、排遗行为等。这与红外相机的布设角度、范围、录制时间有限、录制的视频没有声音等因素有关,在进一步的研究中可以通过增加红外相机数量、丰富角度设置、扩展布设范围(如在农田等生境中布设)、提高红外相机性能等方面获取更多的有效数据,从而开展更深入研究。
致谢: 感谢卧龙国家级自然保护区邓生保护站杨建站长在红外相机布设过程中的宝贵建议;感谢杨文兵、杨洪协助野外工作;感谢西华师范大学生命科学学院的赵钱生、王娟、王盼、夏珊珊、吴沛桦、彭东、李安锐、张宗保、杨远丽等同学参与数据的整理录入工作。
崔多英, 牛克锋, Luen TC, 等. 2014. 黔金丝猴的行为谱及PAE编码系统[J]. 四川动物, 33(6): 815-828. |
胡锦矗. 1990. 大熊猫生物学研究与进展[M]. 成都: 四川科学技术出版社.
|
蒋志刚. 2000. 麋鹿行为谱及PAE编码系统[J]. 兽类学报, 20(1): 1-12. DOI:10.3969/j.issn.1000-1050.2000.01.001 |
李晟, 王大军, 肖治术, 等. 2014. 红外相机技术在我国野生动物研究与保护中的应用与前景[J]. 生物多样性, 22(6): 685-695. |
李勇, 任宝平, 黎大勇, 等. 2013. 滇金丝猴的行为谱及PAE编码系统[J]. 四川动物, 32(5): 641-650. |
任宝平, 夏述忠, 李庆芬, 等. 2002. 圈养雄性川金丝猴交配模式[J]. 动物学报, 48(5): 577-584. |
任仁眉, 严康慧, 苏彦捷, 等. 2000. 金丝猴的社会[M]. 北京: 北京大学出版社.
|
苏彦捷, 任仁眉, 严康慧, 等. 1990. 川金丝猴社会行为模式的观察研究[J]. 心理学报, 22(2): 159-167. |
谭迎春, 欧阳志云, 张和明, 等. 1995. 卧龙自然保护区生物多样性空间特征研究[J]. 中国生物圈保护区, (3): 19-24. |
铁军, 张晶, 彭林鹏, 等. 2009. 神农架川金丝猴栖息地优势树种生态位及食源植物[J]. 植物生态学报, 33(3): 482-491. DOI:10.3773/j.issn.1005-264x.2009.03.007 |
汪松. 1998. 中国濒危动物红皮书:兽类[M]. 北京: 科学出版社.
|
王盼, 李玉杰, 张晋东, 等. 2018. 卧龙国家级自然保护区野生岩羊行为谱及PAE编码系统[J]. 四川动物, 372(2): 211-218. |
王晓卫, 李保国, 吴晓民, 等. 2007. 依据取食优势判断秦岭川金丝猴群OMU的等级地位[J]. 兽类学报, 27(4): 344-349. DOI:10.3969/j.issn.1000-1050.2007.04.005 |
蔚培龙, 杨敬元, 鲍伟东, 等. 2009. 神农架川金丝猴投食群的攻击行为及等级序列[J]. 兽类学报, 29(1): 7-11. DOI:10.3969/j.issn.1000-1050.2009.01.002 |
蔚培龙. 2011. 神农架原始森林中的精灵——川金丝猴[J]. 旅游纵览, (2): 62-65. |
严康慧, 苏彦捷, 任仁眉. 2006. 川金丝猴社会行为节目及其动作模式[J]. 兽类学报, 26(2): 129-135. DOI:10.3969/j.issn.1000-1050.2006.02.004 |
杨晓军. 1996. 笼养川金丝猴哺乳行为的观察[J]. 甘肃农业大学学报, 31(4): 334-338. |
余建秋, 刘选珍, 王强, 等. 2015. 环境丰容对圈养川金丝猴行为影响的初步研究[J]. 四川动物, 34(2): 245-250. |
禹洋. 2012.川金丝猴(Rhinopithecus roxellana)相互理毛行为研究[D].长沙: 中南林业科技大学. http://cdmd.cnki.com.cn/article/cdmd-10538-1012397985.htm
|
张晋东, 李玉杰, 黄金燕, 等. 2018. 利用红外相机建立野生水鹿行为谱及PAE编码系统[J]. 兽类学报, 38(1): 1-11. |
张晋东, 李玉杰, 李仁贵. 2015. 红外相机技术在珍稀兽类活动模式研究中的应用[J]. 四川动物, 34(5): 671-676. |
赵海涛, 王晓卫, 齐晓光, 等. 2013. 秦岭川金丝猴成年雌性优势资源占有与社会等级的判定[J]. 兽类学报, 33(3): 215-222. |
Li YM. 2010. Seasonal variation of diet and food availability in a group of Sichuan snub-nosed monkeys in Shennongjia Nature Reserve, China[J]. American Journal of Primatology, 68(3): 217-233. |
Nishida T. 1987. Local traditions and cultural transmission[M]// Smuts BB, Cheney DL, Seyfarth RM. Primate societies. Chicago: University of Chicago Press.
|
Ren R, Yan K, Su Y, et al. 1995. The reproductive behavior of golden monkeys in captivity (Rhinopithecus roxellana roxellana)[J]. Primates, 36(1): 135-143. DOI:10.1007/BF02381921 |
Whiten A, Goodall J, Mcgrew WC, et al. 2001. Charting cultural variation in chimpanzees[J]. Behaviour, 138(11/12): 1481-1516. |